西藏巴松錯夏季浮游植物群落的分布
2023-04-29 20:08:07劉洋安瑞志王陳潘成梅巴桑
水生態學雜志 2023年6期
關鍵詞:群落結構
劉洋 安瑞志 王陳 潘成梅 巴桑
摘要:揭示西藏巴松錯浮游植物群落結構的水平和垂直分布特征及其與環境間的關系,為進一步研究高寒湖泊生態系統提供基礎資料。于2020年7月對巴松錯出入河流及其湖體設置 15 個水平采樣點,并選擇湖中心樣點B15 進行垂直分層采樣,同時測定水體相關理化指標。調查共采集水樣275個,分析了浮游植物物種組成、細胞豐度、生物量和物種多樣性等群落結構特征,并探討浮游植物對水平和垂直環境梯度變化的響應。結果表明:(1)巴松錯夏季共鑒定浮游植物167種,隸屬于5門8綱20目29科66屬,物種組成為硅藻-綠藻-藍藻型;(2)在水平方向上,平均細胞豐度為1.845×104 個/L,平均生物量為0.012 mg/L;在垂直方向上,平均細胞豐度為1.006×104 個/L,平均生物量為0.006 mg/L;(3)巴松錯夏季浮游植物群落結構水平分布較為均勻,群落復雜程度相對較高,較垂直分布更穩定;(4)RDA分析結果表明,電導率是影響巴松錯夏季浮游植物水平群落變異的主要環境因子,濁度是影響垂直變異的主要環境因子。
關鍵詞:浮游植物;群落結構;水平分布;垂直分布;巴松錯
中圖分類號:Q179.1? ? ? ? 文獻標志碼:A? ? ? ? 文章編號:1674-3075(2023)06-0104-09
作為湖泊生態系統的初級生產者,浮游植物的動態分布受多因素控制,與水體透明度、濁度、水溫、鹽度、DO和pH等水環境因子密切相關,同時受到水平和垂直環境梯度的影響(Ptacnik et al,2008)。孫萍等(2020)調查了熱帶東印度洋春季浮游植物群落結構水平和垂向上浮游植物種類及豐度的差異性,邢冰偉等(2020)分析了九寨溝國家級自然保護區長海夏季浮游植物群落結構的水平和垂直分布,然而有關青藏高原湖泊浮游植物群落結構水平和垂直分布方面的相關研究鮮有報道。
青藏高原高寒湖泊分布星羅密布,占中國湖泊總面積一半以上,是高原陸-氣系統的基本組成要素(靳錚等,2020)。相對于其他湖泊而言,高寒湖泊受人類活動干擾較小,對氣候變化的響應更快、更敏感,基本保持原始生態系統。自然狀態下湖泊水生生物群落組成及其對環境的響應,已引起國內外學者的廣泛關注(王捷等,2015;陳立婧等,2013;Li et al,2021)。巴松錯位于尼洋河最大支流巴河的寬谷中,是西藏東南部最大的高山冰川堰塞湖之一,在維持區域生態平衡、調節氣候、保持生物多樣性和提高居民生產、生活等方面起著重要作用。目前,有關巴松錯的水生生物學研究主要集中在浮游植物(安瑞志等,2020;2021)和原生動物(楊欣蘭等,2022)等方面,但針對巴松錯浮游植物的系統和持續研究仍較少。
為了解西藏巴松錯浮游植物群落結構水平和垂直分布差異及其與環境的關系,于2020年7月對巴松錯出入河流及其湖體的浮游植物進行了水平和垂直分層定性定量采集,研究分析了浮游植物物種組成、細胞豐度、生物量和物種多樣性等群落結構特征,掌握巴松錯水平和垂直上浮游植物群落分布格局和規律,并探討浮游植物對水平和垂直環境梯度變化的響應,以期為進一步研究高寒湖泊生態系統提供資料。
1? ?材料與方法
1.1? ?研究區域概況
巴松錯(29°59′~30°04′N,93°53~94°01′E)位于念青唐古拉山東端南麓、西藏東南部和林芝市西北部工布江達境內,屬尼洋河水系,外觀狹長呈新月形,分為東、西2個湖盆,湖面平均海拔3 469 m,最大湖深約120 m,湖泊面積25.9 km2,流域面積 1 290 km2。該湖泊四面環山,氣候溫和,屬于典型的高原溫帶半濕潤季風氣候,年均降水量 646 mm,主要集中在4-10月,年平均氣溫6.3 ℃,年無霜期175 d,年日照時數為2 016 h(中國科學院青藏高原綜合科學考察隊,1981)。湖水補給主要為天然降水和仲措弄巴曲、新措弄巴曲、白朗曲、羅結曲和扎拉河的冰川融雪水,經由巴河流出,最終匯入尼洋河(中國科學院青藏高原綜合科學考察隊,1984)。
1.2? ?樣點設置
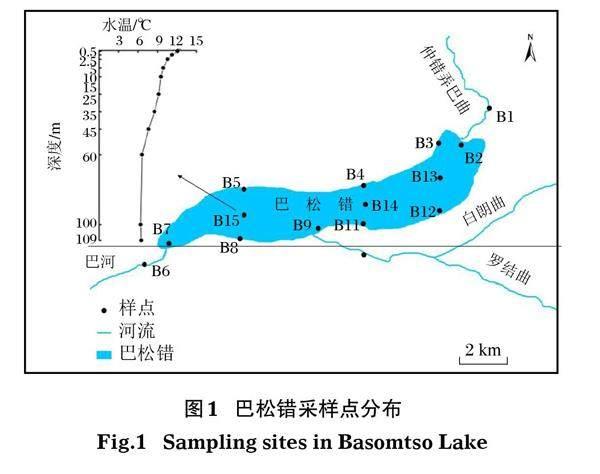
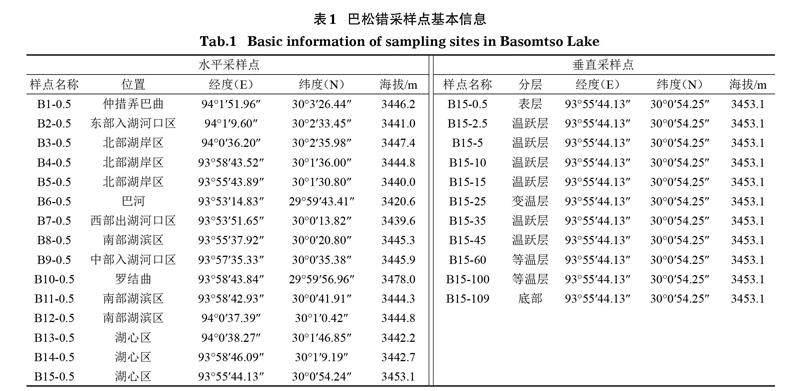
2020年7月,根據巴松錯的生態環境特征、功能分區與梯度結構等特征,在巴松錯出入河流及其湖體設置15個水平采樣點(圖1),并選擇湖中心樣點B15進行垂直分層采樣,現場使用傳統掛錘式測得B15樣點水深為109.5 m,使用賽氏盤測定水體透明度為0.69 m;現場以1 m為間隔測定溫度,以≥0.1℃/m的溫度變化梯度為溫躍層深度(黃磊等,2015),以表層水溫改變≥0.5℃時所對應的深度為混合層深度(黃亞男等,2017),以湖底以上0.5 m水深作為底層深度(中國科學院南京地理與湖泊研究所,2015),并繪制相應的垂直變化曲線,據此確定垂直采樣深度。采樣點的基本信息見表1。
1.3? ?樣品采集及鑒定
浮游植物定性、定量樣品根據《淡水浮游生物研究方法》(章宗涉和黃祥飛,1991)進行采集,物種鑒定參考相關文獻(遲若文,1990;中國科學院青藏高原綜合科學考察隊,1992;朱蕙忠和陳嘉佑,2000;胡鴻鈞和魏印心,2006)。水深小于40 m的水樣使用有機玻璃采水器采集,水深40 m以上的水樣則使用卡蓋式采水器(QCC15)采集,河流水樣直接使用2 L采水器采集。其中1 L水樣用于定性樣品,現場使用25號浮游生物網過濾濃縮至50 mL,并立即用4%的甲醛溶液固定保存;1 L水樣用于定量樣品,現場立即用15 mL魯哥氏碘液固定,帶回實驗室沉淀48 h后濃縮至30 mL。計數時,移取0.1 mL濃縮勻液至0.1 mL計數框,在OLYMPUS CX23顯微鏡下進行物種鑒定,每個樣品計數2~3次。浮游植物生物量(濕重) 通過體積換算間接獲得(Sun & Liu,2003;Hillebrand et al,1999;Kremer et al,2014)。
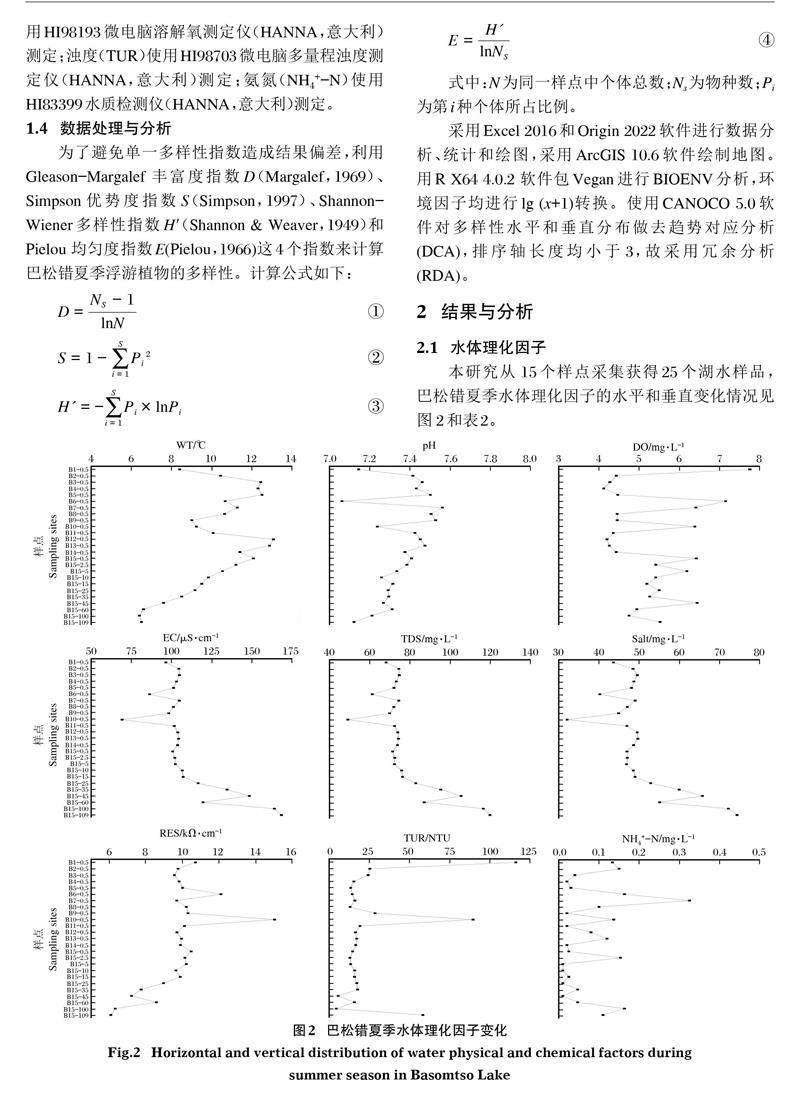
pH、電導率(EC)、鹽度(Salt)及總溶解鹽(TDS)使用PCSTestr35防水型多參數筆試測試儀(EUTECH,美國) 測定;水溫(WT)和電阻率(RES)使用HI98192微電腦電阻率測定儀測定;溶解氧(DO)使用HI98193微電腦溶解氧測定儀(HANNA,意大利)測定;濁度(TUR)使用HI98703微電腦多量程濁度測定儀(HANNA,意大利)測定;氨氮(NH4+-N)使用HI83399水質檢測儀(HANNA,意大利)測定。
1.4? ?數據處理與分析
為了避免單一多樣性指數造成結果偏差,利用Gleason-Margalef 豐富度指數D(Margalef,1969)、Simpson優勢度指數S(Simpson,1997)、Shannon-Wiener多樣性指數H′(Shannon & Weaver,1949)和Pielou 均勻度指數E(Pielou,1966)這4個指數來計算巴松錯夏季浮游植物的多樣性。計算公式如下:
式中:N為同一樣點中個體總數;Ns為物種數;Pi為第i種個體所占比例。
采用Excel 2016和Origin 2022軟件進行數據分析、統計和繪圖,采用ArcGIS 10.6軟件繪制地圖。用R X64 4.0.2 軟件包Vegan進行BIOENV分析,環境因子均進行lg (x+1)轉換。使用CANOCO 5.0軟件對多樣性水平和垂直分布做去趨勢對應分析(DCA),排序軸長度均小于3,故采用冗余分析(RDA)。
2? ?結果與分析
2.1? ?水體理化因子
本研究從 15個樣點采集獲得25個湖水樣品,巴松錯夏季水體理化因子的水平和垂直變化情況見圖 2和表2。
2.2? ?巴松錯浮游植物群落結構
2.2.1? ?物種組成? ?巴松錯夏季共鑒定浮游植物167種,隸屬于5門8綱20目29科66屬,水平方向物種數大于垂直方向,物種組成均以硅藻門-綠藻門-藍藻門為主,裸藻門和隱藻門種類較少,僅出現在水平方向(表3)。
巴松錯夏季浮游植物群落多集合韋恩圖如圖3。在水平方向上,各樣點浮游植物的共有種類僅有1種,為肘狀針桿藻(Synedra ulna)。相比較于水平方向,在垂直方向上,各樣點浮游植物不存在共有種類。
2.2.2? ?細胞豐度和生物量? ?巴松措夏季浮游植物細胞豐度和生物量的水平和垂直變化如圖4。在水平方向上,細胞豐度的變化范圍是0.195×104~7.095×104 個/L,平均值為1.845×104 個/L,不同樣點間細胞豐度差異性較大,最高值在B13-0.5,為7.095×104 個/L,最低值在B12-0.5,為0.195×104 個/L;硅藻門細胞豐度占比相對最高,為22.22%~100%,其次是綠藻門和藍藻門,細胞豐度變化范圍分別為0~0.628×104和0~1.256×104 個/L,占比為0~55%和0~38.89%;生物量為0.003~0.044 mg/L,平均值為0.012 mg/L,不同樣點間浮游植物生物量差異性較大,最高值在B13-0.5,最低值在B14-0.5;硅藻門生物量占比相對最高,變化范圍為10.98%~100%,其次是綠藻門和藍藻門,生物量變化范圍分別為0~0.013和0~0.008 mg/L,占比為0~88.94%和0~27.49%;
在垂直方向上,細胞豐度的變化范圍是0.21×104~2.79×104 個/L,平均值為1.006×104 個/L,不同垂直分層細胞豐度差異性較大,最高值在B15-100,為2.79×104 個/L,最低值在B15-109,為0.21×104 個/L,硅藻門細胞豐度占比相對最高,變化范圍為54.55%~100%,藍藻門僅在B15-109樣點出現,占該樣點豐度的45.45%,綠藻門在B15-5和B15-15樣點出現,分別占細胞豐度的6.25%和2.33%;生物量的變化為0.001~0.027 mg/L,平均值為0.006 mg/L,不同垂直分層浮游植物生物量差異性較大,最高值在B15-15,為0.027 mg/L,最低值在B15-25,為0.001 mg/L;硅藻門生物量占比相對最高,為31.81%~100%,藍藻門僅在B15-109樣點出現,占該樣點生物量的68.19%,綠藻門在B15-5和B15-15樣點出現,分別占生物量的11.56%和0.86%;
2.2.3? ?多樣性指數? ?巴松措各站點夏季浮游植物物種多樣性的水平和垂直變化如圖5。在水平方向上,Margalef豐富度指數為0.756~2.807,均值1.734;Simpson優勢度指數為0.695~0.921,均值為0.832;Shannon-Wiener多樣性指數為1.487~2.792,均值為2.28;Pielou均勻度指數為0.715~0.858,均值為0.806;在垂直方向上,Margalef豐富度指數為0.392~1.507,均值為0.955;Simpson優勢度指數為0.583~0.881,均值為0.698;Shannon-Wiener多樣性指數為0.998~2.251,均值為1.643;Pielou均勻度指數為0.685~0.853,均值為0.739。總體上看,水平方向的Margalef豐富度指數、Simpson優勢度指數、Shannon-Wiener多樣性指數和Pielou均勻度指數均高于垂直方向。
2.3? ?多樣性與環境因子的關系
BIOENV分析發現,pH、DO、EC、TDS、Salt和RES對巴松錯夏季浮游植物群落多樣性水平分布的影響較大;EC和TUR對巴松錯夏季浮游植物群落多樣性垂直分布的影響較大。
為進一步分析上述最優環境因子對巴松錯夏季浮游植物群落多樣性水平和垂直分布的影響,對群落多樣性與環境因子進行RDA分析(圖6)。結果表明,在水平方向上,第1排序軸與第2排序軸對群落多樣性的方差的解釋率分別為48.45%和0.94%,群落多樣性與環境因子的相關性分別為0.7117和0.4762,前2軸累計方差為49.39%,上述環境因子對巴松錯夏季浮游植物群落多樣性水平分布的影響大小順序為EC、TDS、DO、Salt、pH和RES,其中EC(P=0.038,F=6.1)是顯著解釋性變量,對群落變異的解釋率為31.9%,是影響巴松錯夏季浮游植物水平群落變異的主要環境因子。在垂直方向上,第1排序軸與第2排序軸對群落多樣性的方差的解釋率分別為57.67%和0.3%,群落多樣性與環境因子的相關性分別為0.7693和0.4424,前2軸累計方差為57.97%,上述環境因子對巴松錯夏季浮游植物群落多樣性垂直分布的影響大小順序為TUR和EC,其中TUR(P=0.008,F=12.1)是顯著解釋性變量,對群落變異的解釋率為57.4%,是影響巴松錯夏季浮游植物群落垂直變異的主要環境因子。
3? ?討論
3.1? ?浮游植物群落結構特征
本研究共鑒定出浮游植物167種,隸屬于5門8綱20目29科66屬,少于本課題組之前報道的242種(安瑞志等,2021),可能和2次調查的采樣周期、樣點范圍及分類鑒定的詳細程度等因素有關。這也反映了巴松錯浮游植物群落仍然處于動態演替過程中。從浮游植物種類來看,物種組成以硅藻占絕對優勢, 其次為綠藻和藍藻,這與本課題組2017-2018年間對巴松錯浮游植物調查結果基本一致(安瑞志等,2021),主要原因可能是大多數硅藻都是狹冷性物種,適合在較寒冷和貧營養的水環境中生活(潘成梅等,2021),而巴松錯是一個過水型湖泊,其水位相較于吞吐型或封閉型湖泊更為穩定(廖夢娜等,2020),浮游植物常年在較低溫水環境下生存,已形成了動態的適應性機制,這一結果與色林錯(劉洋等,2022)、浪錯(扎西拉姆等,2021)的研究結果報道基本一致。
巴松錯夏季浮游植物群落的分布具有較明顯的水平和垂直空間差異,主要體現在物種數、細胞豐度和生物量等方面。結果表明,浮游植物垂直方向的物種數、細胞豐度和生物量均低于水平方向。在水平方向上,生物量最高值出現在湖心區域的B13-0.5樣點,為0.044 mg/L,這主要是因為該樣點EC(104.33 μS/cm)最高,高EC與有機質降解和離子的輸入有著極大的聯系,能夠使藻類的光合作用得到較好的補充,為浮游植物的大量繁殖提供了較好的營養條件(李鋒,2021)。在垂直方向上,生物量最大值出現在B15-15,為0.027 mg/L,主要原因可能是巴松錯為深水湖泊, 在夏季具有明顯的溫躍層,水體混合交換弱,水深15 m是巴松錯浮游植物最適的生長區域,形成了浮游植物上下層分布的明顯差異,這與潘繼征等(2009)的研究和邢冰偉等(2020)的研究結果基本一致。
巴松錯夏季浮游植物群落的的水平和垂直分布差異不僅體現在物種數、細胞豐度和生物量上,還體現在物種多樣性上。物種多樣性是生物與環境因子長期作用和適應的結果,用來衡量浮游植物群落結構是否穩定(邱小琮等,2011)。基于浮游植物對湖泊生態系統群落穩定性進行生物學評價有很多方法,本文選擇了應用最為廣泛的Margalef豐富度指數、Simpson優勢度指數、Shannon-Wiener多樣性指數和Pielou 均勻度指數這4個指數進行評價分析。結果表明,水平方向的4個指數值均高于垂直方向,表明巴松錯夏季浮游植物群落結構水平分布較為均勻,群落復雜程度相對較高,較垂直分布更穩定,這與柯志新等(2011)和籍辰旭等(2021)的研究結果基本一致。
3.2? ?浮游植物多樣性與環境因子的關系
環境因子是浮游植物群落分布的主要影響因素,水體透明度、濁度、溶解氧、EC、pH、水溫和鹽度等環境因子都對浮游植物群落分布產生影響(趙秀俠等,2018)。本研究中,BIOENV分析結果顯示,在水平方向上,pH、DO、EC、TDS、Salt和RES對巴松錯夏季浮游植物物種多樣性的影響較大;在垂直方向上,EC和TUR對巴松錯夏季浮游植物物種多樣性垂直分布的影響較大。但不同環境因子對浮游植物群落結構變化的影響也存在差異。王宇飛等(2015)的研究發現影響漢豐湖夏季浮游植物群落結構特征的主要因素是溫度、DO、pH、TN、TP、 DN和DP;牛遠等(2016)的研究發現環境變量較好地解釋了撫仙湖夏季熱分層時期浮游植物群落組成的空間變化,WT、EC和pH是最關鍵影響因子,而營養鹽的作用并不顯著。本研究中,RDA分析結果表明,EC是影響巴松錯夏季浮游植物水平群落變異的主要環境因子,TUR是影響垂直變異的主要環境因子。EC與水體營養鹽含量有關系,而營養鹽為浮游植物提供充足的營養,EC通過影響浮游植物的代謝活動和繁殖從而影響浮游植物的群落結構(趙秀俠等,2021)。而濁度(TUR)是由于水中含有泥沙、黏土、有機物、無機物、浮游生物等懸浮物質所造成的,可使光散射或吸收,影響浮游植物對光的吸收(聶小芬等,2021)。本研究采樣時間為夏季,降雨徑流所攜帶的陸源物質發生沉降,也會引起水體濁度升高,透明度降低,光合作用減弱,在垂向上易于產生生物分層,對浮游植物的垂直分布具有一定的抑制作用(安瑞志等,2021)。因此,浮游植物多樣性的變化是多種環境因子在水平與垂直分布共同作用的結果。
本研究只涉及到1個季節以及水體中的部分理化因子, 不能得出巴松錯浮游植物水平與垂直分布的季節動態變化,今后有待于加強對其季節動態變化研究,并進行初級生產力估算的研究,以便為碳中和背景下高寒湖泊固碳潛力的計量及估算提供數據支持。
參考文獻
安瑞志,潘成梅,劉洋,等,2020. 巴松措(湖)夏季浮游植物優勢種群分布格局與共存機制研究[J].高原科學研究,4(2):27-36.
安瑞志,潘成梅,塔巴拉珍,等,2021. 西藏巴松錯浮游植物功能群垂直分布特征及其與環境因子的關系[J].湖泊科學,33(1):86-101.
陳立婧,楊菲,吳淑賢,等,2013. 西藏那曲地區鹽湖浮游植物群落結構的特征[J].上海海洋大學學報,22(4):577-585.
遲若文,1990.西藏硅藻圖集[M].拉薩:西藏人民出版社.
胡鴻鈞,魏印心,2006.中國淡水藻類—系統、分類及生態[M].北京:科學出版社.
黃磊,王君波,朱立平等,2015. 納木錯水溫變化及熱力學分層特征初步研究[J].湖泊科學,27(4):711-718.
黃亞男,紀道斌,龍良紅,等,2017. 三峽庫區典型支流春季特征及其水華優勢種差異分析[J].長江流域資源與環境,26(3):461-470.
籍辰旭,陳逍遙,陳火榮,等,2021. 三沙灣春季浮游植物群落結構特征[J].漁業研究,43(1):42-52.
靳錚,張雪芹,次旦央宗,2020. 藏南羊卓雍錯湖面非封凍期溫濕變化及輻射平衡分析[J].干旱區研究,37(4):947-955.
柯志新,黃良民,譚燁輝,等,2011. 2007年夏季南海北部浮游植物的物種組成及豐度分布[J].熱帶海洋學報,30(1):131-143.
廖夢娜,金伊麗,李晨瑜,等,2020. 藏東南巴松錯200年沉積過程及其對硅藻記錄的影響[J].生態學報,40(3):1089-1100.
李鋒,2021. 二灘水庫浮游植物群落結構與水環境因子時空變化及其相關性研究[D].重慶:西南大學.
劉洋,安瑞志,巴桑,2022. 西藏色林錯-普若崗日國家公園潛在建設區夏季浮游植物群落生態特征[J].水生態學雜志,43(6):51-58.
聶小芬,楊正健,王從鋒,等,2021. 瀾滄江梯級水庫建設下水體電導率和濁度時空分布特征[J].三峽大學學報(自然科學版),43(1):18-24.
牛遠,孔祥虹,余輝,等,2016. 撫仙湖夏季熱分層時期浮游植物空間分布特征[J].生態學雜志,35(7):1865-1871.
潘成梅,劉洋,安瑞志,等,2021.西藏麥地卡濕地的浮游植物——1.優勢種的時空生態位[J].湖泊科學,33(6):1805-1819.
潘繼征,熊飛,李文朝,等,2009. 撫仙湖浮游植物群落結構、分布及其影響因子[J].生態學報,29(10):5376-5385.
邱小琮,趙紅雪,2011.寧夏沙湖浮游植物群落結構及多樣性研究[J].水生態學雜志,32(1):20-26.
孫萍,李艷,潘玉龍,等,2020. 熱帶東印度洋春季浮游植物群落結構空間特征分析[J].海洋學報,42(8):76-88.
邢冰偉,徐季雄,曹玥,等,2020. 九寨溝國家級自然保護區長海夏季浮游植物群落結構及生態評價[J].湖泊科學,32(4):1088-1099.
王捷,李博,馮佳,等,2015. 西藏西南部湖泊浮游藻類區系及群落結構特征[J].水生生物學報,39(4):837-844.
王宇飛,趙秀蘭,何丙輝,等,2015. 漢豐湖夏季浮游植物群落與環境因子的典范對應分析[J].環境科學,36(3):922-927.
楊欣蘭,潘瑛子,巴桑,2022.巴松措原生動物群落結構季節及垂直分布[J].生態學報,42(08):3216-3227.
趙秀俠,方婷,楊坤,等,2018.安徽沱湖夏季浮游植物群落結構特征與環境因子關系[J].植物科學學報,36(5):687-695.
趙秀俠,盧文軒,梁陽陽,等,2021.安徽三座通江湖泊秋季浮游植物群落結構特征及其影響因子[J].生態學雜志,40(1):67-75.
扎西拉姆,陳美群,牟振波,等,2022. 西藏浪錯夏季浮游植物組成與環境因子的關系[J].水產科學,41(1):137-142.
中國科學院青藏高原綜合科學考察隊,1981.西藏水利[M].北京:科學出版社.
中國科學院青藏高原綜合科學考察隊,1984.西藏河流與湖泊[M].北京:科學出版社.
中國科學院青藏高原綜合科學考察隊,1992.西藏藻類[M].北京:科學出版社.
中國科學院南京地理與湖泊研究所,2015. 湖泊調查技術規程[M].北京:科學出版社.
朱蕙忠,陳嘉佑,2000.中國西藏硅藻[M].北京:科學出版社.
Hillebrand H,Dürselen C D,Kirschtel D,et al,1999. Biovolume calculation for pelagic and benthic microalgae[J]. Journal of Phycology,35(2): 403-424.
Kremer C T,Gillette J P,Rudstam L G,et al,2014. A compendium of cell and natural unit biovolumes for >1200 freshwater phytoplankton species[J]. Ecology,95(10): 2984.
Li Z X,Gao Y,Wang S Y,et al,2021. Phytoplankton community response to nutrients along lake salinity and altitude gradients on the Qinghai-Tibet Plateau[J]. Ecological Indicators,128: 107848.
Margalef R,1969. Diversity and Stability in Ecological Systems[M]. Upton: Brookhacen National Laboratory:13-26.
Pielou E C,1966.The measurements of diversity in different types of biological collections[J].Journal of Theoretical Biology,13:131-144.
Ptacnik R,Lepist L,E Willén,et al,2008. Quantitative responses of lake phytoplankton to eutrophication in Northern Europe[J]. Aquatic Ecology,42(2):227-236.
Shannon C E,Weaver W,1949.The mathematical theory of communication[M]. Urbana:University of Illinois Press.
Simpson E H,1997. Measurement of Diversity[J]. Journal of Cardiothoracic and Vascular Anesthesia,11(6):812-812.
Sun J,Liu D Y,2003. Geometric models for calculating cell biovolume and surface area for phytoplankton. Journal of Plankton Research,25(11): 1331-1346.
(責任編輯? ?鄭金秀)
Horizontal and Vertical Distributions of the Summer Phytoplankton
Community in Basomtso Lake of Xizang, China
LIU Yang1, 2, AN Rui‐zhi1,2, WANG Chen1,2, PAN Cheng‐mei1,2, BA Sang1,2
(1. Center for Carbon Neutrality in the Earth's Third Pole, Tibet University, Lhasa? ?850000, P.R. China;
2. Laboratory of Wetland and Catchment Ecosystem in Tibetan Plateau, School of Ecology
and Environment, Tibet University, Lhasa? ?850000, P.R. China)
Abstract: In this study, we explored the horizontal and vertical distributions of the phytoplankton community and its relationship with environmental factors in Basomtso Lake of Tibet, aiming to provide basic data for the further study of plateau lake ecosystems. In July 2020, horizontal water samples were collected at 15 sampling sites (B1-15) and vertical samples were collected at 11 depths at B15, the sampling site near the center of Basomtso Lake. Physical and chemical parameters of the lake water were also measured. A total of 275 water samples were collected for phytoplankton analysis, focusing on species composition, cell abundance and biomass, and species diversity. The response of phytoplankton to changes in horizontal and vertical environmental gradients were analyzed by redundancy analysis (RDA). Results show that: (1) A total of 167 phytoplankton species from 66 genera, 29 families, 20 orders, 8 classes and 5 phyla were identified, with dominance by bacillariophyta-chlorophyta-cyanophyta. Species richness of the horizontal distribution was higher than that of the vertical distribution. (2) Horizontally, the average cell abundance and biomass were 1.845×104 cells/L and 0.012 mg/L, and vertically, the average cell abundance and biomass were 1.006×104 cells/L and 0.006 mg/L. (3) The Margalef richness, Simpson dominance, Shannon-Wiener diversity and Pielou evenness indices of the summer phytoplankton community in the horizontal distribution were all higher than those of the phytoplankton community in the vertical distribution. These indices show that the horizontal distribution was more uniform, and that community complexity and stability were higher than in the vertical distribution. (4) RDA showed that electrical conductivity (EC) was the primary environmental factor associated with the horizontal distribution of phytoplankton, and turbidity (TUR) was the primary environmental factor associated with the vertical distribution of phytoplankton in Basomtso Lake. The results of this study provide basic data for further study of alpine lake ecosystems.
Key words:phytoplankton; community structure;horizontal distribution; vertical distribution;
Basomtso Lake
收稿日期:2021-12-17? ? ? 修回日期:2023-09-25
基金項目:國家自然科學基金(32070418,31660620);西藏大學培育基金(ZDQMJH19-34);2022 年中央財政支持地方高校改革發展專項資金 (藏財預指[2022]1號)。
作者簡介:劉洋,女,1988年生,實驗師,主要從事高寒濕地浮游植物生態學研究。E-mail:ly@utibet.edu.cn
通信作者:巴桑,男,1970年生,教授,主要從事高寒濕地與流域生態系統研究。E-mail:hbasang2003@aliyun.com
猜你喜歡
熱帶農業科學(2016年11期)2017-01-21 14:59:21
熱帶農業科學(2016年11期)2017-01-21 14:57:08
河北漁業(2015年12期)2015-12-21 14:37:31
河北漁業(2015年11期)2015-11-18 16:39:54
河北漁業(2015年11期)2015-11-18 16:37:49
河北漁業(2015年10期)2015-10-15 11:14:57
湖北農業科學(2015年18期)2015-10-13 21:34:01
河北漁業(2015年7期)2015-07-21 11:06:52
河北漁業(2015年6期)2015-06-24 11:04:37
安徽農學通報(2015年5期)2015-03-23 23:51:20
