濟南濟西濕地公園浮游動物功能類群及其對環境因子的響應
2023-04-29 21:17:39商書芹賈麗王帥帥殷旭旺白海鋒
水生態學雜志 2023年6期
商書芹 賈麗 王帥帥 殷旭旺 白海鋒
摘要:研究濟南濟西濕地公園浮游動物的功能類群及其與環境因子的關系,了解濕地生態系統的結構與功能,可為城市濕地環境狀況的評價及管理提供理論依據。于2020年4月至2021年3月在濟西濕地公園6個采樣點開展了浮游動物群落結構及環境因子調查,并分析了影響浮游動物功能群的主要環境因子。結果表明:濟西濕地公園全年共采集到浮游動物4個大類66種,平均密度為181.8個/L,平均生物量為0.210 mg/L;浮游動物劃分為7個功能群,春季、夏季、冬季均以功能群輪蟲濾食者(RF)和原生動物濾食者(PF)為主,RF最高占比為62.86%(冬季)、PF最高占比為20.75%(春季),秋季以功能群RF和小型浮游動物濾食者(SCF)為主,占比分別為58.54%、12.20%;浮游動物Shannon-Wiener多樣性指數、Pielou均勻度指數和Margalef豐富度指數平均值分別為2.504、0.708、2.307;綜合營養狀態指數水質評價結果顯示濟西濕地公園水質整體處于中營養化水平;環境因子與生物典范對應分析表明WT、COND、和PO[3-4]是顯著影響浮游動物功能群分布的主要環境因子。
關鍵詞:浮游動物;功能群;多樣性評價;濟西濕地公園
中圖分類號:Q178.1? ? ? ? 文獻標志碼:A? ? ? ? 文章編號:1674-3075(2023)06-0113-09
浮游動物依賴水流作用在水體中營浮游生活,作為水生態系統的初級消費者,通過“上行下效”作用進行著系統內營養轉化、能量流動和信息傳遞等生態過程,其種類和數量的變動會影響水體中其他水生生物的群落結構。浮游動物具有種類多、世代時間短、對環境敏感和方便采集等特點,常被選作指示河流、湖泊等水體受污染的指示生物類群(Elijah et al,2011),如在富營養化水體中,臂尾輪蟲(Brachionus)、龜甲輪蟲(Keratella)、劍水蚤(Cyclops)等一般作為優勢種群出現(朱浩等,2020)。浮游動物對水溫、溶解氧、pH、營養鹽以及水體營養化程度等環境因子的變化極其敏感,能很好地評價水體質量(馮世敏等,2017;白海鋒等,2021)。功能群是指存在相似結構或功能的一類物種集合體,是基于系統內相似特征或相似行為進行的物種歸類(龔森森等,2022),通過功能群的研究,可以更直接地反映生態環境影響生物群落的生態過程。因此,對浮游動物功能群的研究能夠更深入了解浮游動物在水體能量流動中的作用,對于分析評價水域生態系統健康狀況具有重要意義(Hoeinghaus et al,2007)。
濟西國家濕地公園位于山東省濟南市西部城區,南臨南水北調濟平干渠,東臨玉清湖水庫,西北臨黃河,屬黃河流域濟南段典型的城市濕地。地理坐標116°46'51.9"~116°48'53.6" E,36°38'22.2"~36°40'2.3" N,總面積約33.4 km2,一期建設已完成11.3 km2。濟西濕地是平原型積水洼地,匯集了黃河、濟平干渠、玉符河等豐富的地表水資源,水質清澈,濕地內河流、港汊交織形成大小島嶼近100座,水生生物豐富多樣。濟西濕地承擔著濟南市的飲用水源保護、防洪安全、生態涵養等重要功能,被稱為“城市之腎”,根據規劃濟西濕地劃分為保育區、恢復重建區、宣教展示區、合理利用區和管理服務區等區域。研究表明濕地生態系統是物質循環與能量轉換的重要生態載體,亦是受人類活動干擾最為強烈的生態系統之一,特別是城市濕地(Wang et al, 2019)。因此,濟西濕地的建設和維護不僅在城市生態文明建設中具有重要作用,而且在南水北調中線的生態環境保護中具有突出的地位。目前關于濟西濕地水生生物資源研究報道相對較少,而對濕地浮游動物功能群的劃分及分析研究還未開展。基于此,為了掌握濟西濕地公園的生態環境狀況和水生生物資源,2020-2021年在濕地各功能區共選擇6個調查點,調查探討了浮游動物功能群的結構特征及環境影響因素,以期為濟西濕地的水生態系統功能多樣性研究和可持續發展利用提供支撐。
1? ?材料與方法
1.1? ?點位設置
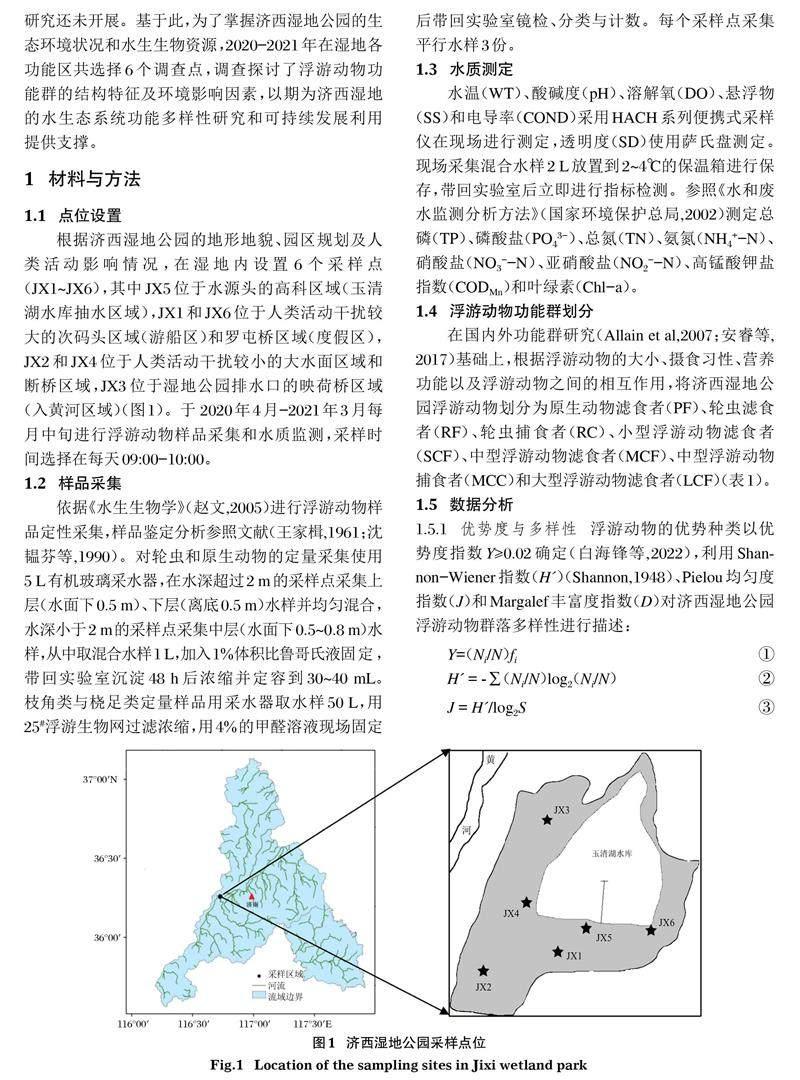
根據濟西濕地公園的地形地貌、園區規劃及人類活動影響情況,在濕地內設置6個采樣點(JX1~JX6),其中JX5位于水源頭的高科區域(玉清湖水庫抽水區域),JX1和JX6位于人類活動干擾較大的次碼頭區域(游船區)和羅屯橋區域(度假區),JX2和JX4位于人類活動干擾較小的大水面區域和斷橋區域,JX3位于濕地公園排水口的映荷橋區域(入黃河區域)(圖1)。于 2020年4月-2021年3月每月中旬進行浮游動物樣品采集和水質監測,采樣時間選擇在每天09:00-10:00。
1.2? ?樣品采集
依據《水生生物學》(趙文,2005)進行浮游動物樣品定性采集,樣品鑒定分析參照文獻(王家楫,1961;沈韞芬等,1990)。對輪蟲和原生動物的定量采集使用5 L有機玻璃采水器,在水深超過2 m的采樣點采集上層(水面下0.5 m)、下層(離底0.5 m)水樣并均勻混合,水深小于2 m的采樣點采集中層(水面下0.5~0.8 m)水樣,從中取混合水樣1 L,加入1%體積比魯哥氏液固定,帶回實驗室沉淀48 h后濃縮并定容到30~40 mL。枝角類與橈足類定量樣品用采水器取水樣50 L,用25#浮游生物網過濾濃縮,用4%的甲醛溶液現場固定后帶回實驗室鏡檢、分類與計數。每個采樣點采集平行水樣3份。
1.3? ?水質測定
水溫(WT)、酸堿度(pH)、溶解氧(DO)、懸浮物(SS)和電導率(COND)采用HACH系列便攜式采樣儀在現場進行測定,透明度(SD)使用薩氏盤測定。現場采集混合水樣2 L放置到2~4℃的保溫箱進行保存,帶回實驗室后立即進行指標檢測。參照《水和廢水監測分析方法》(國家環境保護總局,2002)測定總磷(TP)、磷酸鹽(PO43-)、總氮(TN)、氨氮(NH4+-N)、硝酸鹽(NO3--N)、亞硝酸鹽(NO2--N)、高錳酸鉀鹽指數(CODMn)和葉綠素(Chl-a)。
1.4? ?浮游動物功能群劃分
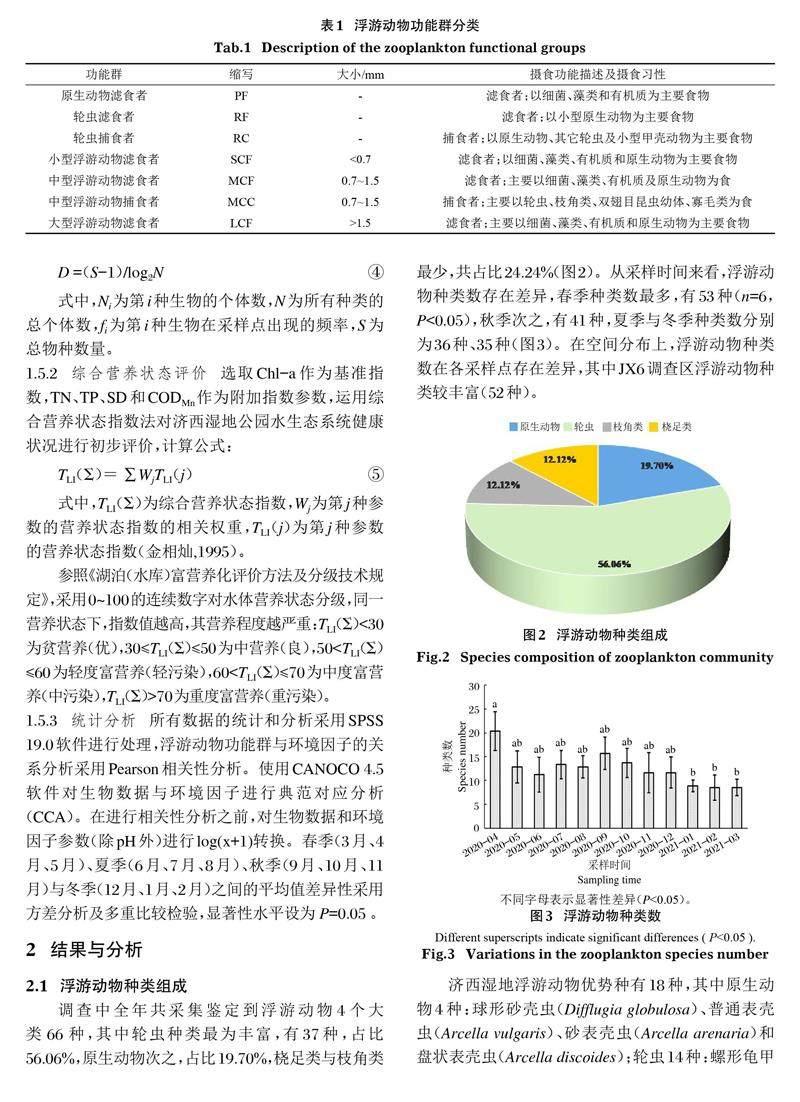
在國內外功能群研究(Allain et al,2007;安睿等,2017)基礎上,根據浮游動物的大小、攝食習性、營養功能以及浮游動物之間的相互作用,將濟西濕地公園浮游動物劃分為原生動物濾食者(PF)、輪蟲濾食者(RF)、輪蟲捕食者(RC)、小型浮游動物濾食者(SCF)、中型浮游動物濾食者(MCF)、中型浮游動物捕食者(MCC)和大型浮游動物濾食者(LCF)(表1)。
1.5? ?數據分析
1.5.1? ?優勢度與多樣性? ?浮游動物的優勢種類以優勢度指數Y≥0.02確定(白海鋒等,2022),利用Shannon-Wiener指數(H')(Shannon,1948)、Pielou均勻度指數(J)和Margalef豐富度指數(D)對濟西濕地公園浮游動物群落多樣性進行描述:
式中,Ni為第i種生物的個體數,N為所有種類的總個體數,fi為第i種生物在采樣點出現的頻率,S為總物種數量。
1.5.2? ?綜合營養狀態評價? ?選取Chl-a作為基準指數,TN、TP、SD和CODMn作為附加指數參數,運用綜合營養狀態指數法對濟西濕地公園水生態系統健康狀況進行初步評價,計算公式:
式中,TLI(Σ)為綜合營養狀態指數,Wj為第j種參數的營養狀態指數的相關權重,TLI(j)為第j種參數的營養狀態指數(金相燦,1995)。
參照《湖泊(水庫)富營養化評價方法及分級技術規定》,采用0~100的連續數字對水體營養狀態分級,同一營養狀態下,指數值越高,其營養程度越嚴重:TLI(Σ)<30為貧營養(優),30≤TLI(Σ)≤50為中營養(良),50
1.5.3? ?統計分析? ?所有數據的統計和分析采用SPSS 19.0軟件進行處理,浮游動物功能群與環境因子的關系分析采用Pearson相關性分析。使用CANOCO 4.5軟件對生物數據與環境因子進行典范對應分析(CCA)。在進行相關性分析之前,對生物數據和環境因子參數(除pH外)進行log(x+1)轉換。春季(3月、4月、5月)、夏季(6月、7月、8月)、秋季(9月、10月、11月)與冬季(12月、1月、2月)之間的平均值差異性采用方差分析及多重比較檢驗,顯著性水平設為 P=0.05 。
2? ?結果與分析
2.1? ?浮游動物種類組成
調查中全年共采集鑒定到浮游動物4個大類66種,其中輪蟲種類最為豐富,有37種,占比56.06%,原生動物次之,占比19.70%,橈足類與枝角類最少,共占比24.24%(圖2)。從采樣時間來看,浮游動物種類數存在差異,春季種類數最多,有53種(n=6,P<0.05),秋季次之,有41種,夏季與冬季種類數分別為36種、35種(圖3)。在空間分布上,浮游動物種類數在各采樣點存在差異,其中JX6調查區浮游動物種類較豐富(52種)。
不同字母表示顯著性差異(P<0.05)。
濟西濕地浮游動物優勢種有18種,其中原生動物4種:球形砂殼蟲(Difflugia globulosa)、普通表殼蟲(Arcella vulgaris)、砂表殼蟲(Arcella arenaria)和盤狀表殼蟲(Arcella discoides);輪蟲14種:螺形龜甲輪蟲(Keratella cochlearis)、矩形龜甲輪蟲(Keratella quadrala)、緣板龜甲輪蟲(Keratella ticinensis)、針簇多肢輪蟲(Polyarthra trigla)、長肢多肢輪蟲(Polyarthra dolichoptera)、唇形葉輪蟲(Notholon labis)、顫動疣毛輪蟲(Synchacta tremula)、卜氏晶囊輪蟲(Asplanchna brightwelli)、長三肢輪蟲(Filinia longisela)、暗小異尾輪蟲(Trichocerca pusilla)、萼花臂尾輪蟲(Brachionus calyciflorus)、角突臂尾輪蟲(Brachionus angularis)、裂足臂尾輪蟲(Brachionus diversicornis)、剪形臂尾輪蟲(Brachionus forficula)。浮游動物種類優勢度最大的是螺形龜甲輪蟲(Y=0.438),在各調查點出現次數最多的是螺形龜甲輪蟲和針簇多肢輪蟲。
2.2? ?浮游動物功能群
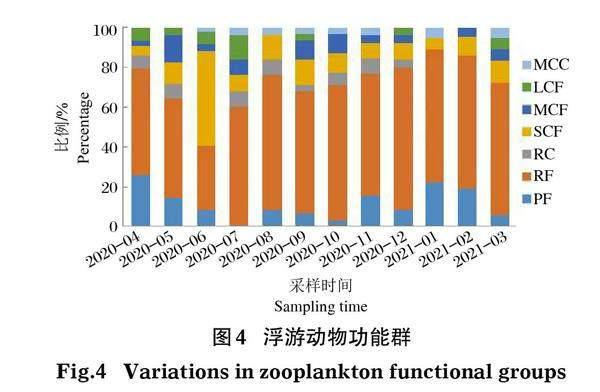
1月-12月調查中濟西濕地各采樣點采集到的浮游動物劃分為7個功能群,其中PF13種,占比19.70%,RF34種,占比51.52%,RC3種,占比4.55%,SCF6種,占比9.09%,MCF5種,占比7.58%,LCF4種,占比6.06%,MCC1種,占比1.52%。浮游動物功能群在時間上呈季節性變化,春季以功能群RF和PF占優勢,種類數占比分別為52.83%、20.75%;夏季與春季一致,RF和PF占優勢,種類數占比分別為58.33%、11.11%;秋季功能群RF和SCF占優勢,分別為58.54%、12.20%;冬季與春夏2季保持一致,以功能群RF和PF占優勢,種類數占比分別為62.86%、17.14%。春夏季LCF多于秋冬季,MCC在4個季節中占比最小(圖4)。
浮游動物密度變化范圍為16.0~824.0個/L,年平均值為181.8個/L。其中,夏季浮游動物平均密度顯著高于秋季(P<0.05),冬季浮游動物平均密度顯著高于春季(P<0.05),密度變化趨勢呈現出冬季>夏季>秋季>春季,密度最大值出現在7月份(322.2個/L)(圖5)。4個季節浮游動物生物量為0.004~1.667 mg/L,年平均值為0.210 mg/L,各采樣點生物量的變化趨勢與密度變化趨勢存在差異。其中,春季浮游動物平均生物量顯著高于夏、秋、冬季浮游動物平均生物量(P<0.05),生物量變化趨勢呈現出春季>秋季>夏季>冬季,生物量最大值出現在4月份(0.556 mg/L)(圖6)。
2.3? ?浮游動物多樣性
濟西濕地公園浮游動物生物多樣性分析結果如圖7所示。浮游動物Shannon-Wiener多樣性指數(H')變化范圍為1.990~3.045,平均值為2.504,多樣性指數季節變化趨勢表現出秋季(2.656)>春季(2.595)>夏季(2.493)>冬季(2.273),季節差異不顯著,4月生物多樣性指數最大。Pielou均勻度指數(J)的變化范圍為0.642~0.779,平均值為0.708,季節變化特征為秋季(0.715)>冬季(0.710)>春季(0.703)>夏季(0.702),差異不顯著,3月均勻度指數最小。Margalef豐富度指數(D)的變化范圍為1.451~3.791,平均值為2.307,季節變化特征為秋季(2.649)>夏季(2.331)>春季(2.322)>冬季(1.925),冬季與春季、夏季和秋季均存在顯著差異(n=6,P<0.05),豐富度指數最大值出現在4月。
2.4? ?環境因子與營養狀態
濟西濕地公園環境因子的變化如表2所示,DO和NH4+-N全年平均達Ⅰ類水質標準,TP和CODMn全年平均達Ⅱ類水質標準,TN全年平均達Ⅲ類水質標準。隨著時間變動,WT存在季節性顯著差異(P<0.05),NO2--N季節差異不顯著。DO、pH、COND和SD冬季均高于其他季節,SS、TN和NO3--N秋季均大于其他季節,CODMn春季最高,夏季TP、Chl-a、NH4+-N相對較高,氮磷比值年平均大于16。綜合營養狀態指數評價顯示,濟西濕地公園水體夏季處于輕度富營養化水平,春季、秋季和冬季,均處于中營養化水平。從整體來看,濟西濕地公園水體呈現中營養化,污染程度為輕度,季節差異不顯著。
2.5? ?浮游動物功能群與環境因子相關性
浮游動物功能群之間Pearson相關分析顯示浮游動物7個功能群之間的相關性不明顯(表3),而浮游動物功能群與環境因子之間Pearson相關分析顯示浮游動物功能群密度與14個環境因子中的WT、COND、DO、NO2- -N、TP、PO43-、SS及Chl-a具有顯著的相關性(P<0.05)。其中,浮游動物功能群SCF同COND 為極顯著負相關(n=6,P<0.01),同NO2- -N為極顯著正相關(P<0.01),WT和TP與浮游動物功能群SFC為顯著正相關(P<0.05),DO與浮游動物功能群SFC為顯著負相關(P<0.05);浮游動物功能群LCF同PO43--P為顯著正相關(P<0.05),浮游動物功能群MCF同SS為顯著負相關(P<0.05),浮游動物功能群PF同Chl-a為顯著負相關(P<0.05)(表4)。
對浮游動物功能群的時空分布進行去趨勢分析(DCA),排序結果顯示4個軸最大長度大于4,屬于單峰型模型,使用典范對應分析(CCA)對浮游動物功能群與Pearson相關分析篩選出來的8個主要環境因子進行分析。CCA排序表明2個排序軸物種與環境因子之間的相關系數均為1,表明排序結果可靠,能夠很好地反映出浮游動物與環境因子之間的相互關系。排序分析結果顯示,環境因子中WT、COND和PO43-是顯著影響浮游動物功能群時空變化的主要環境因子(P<0.05)(圖8)。
3? ?討論
3.1? ?浮游動物群落結構變化特征
本研究結果顯示濟西濕地公園浮游動物物種組成以普生性小型浮游動物(輪蟲―原生動物)為主,占總種類數的3/4,大型浮游動物種類數所占比例相對較低,這與國內許多水體浮游動物種類組成結構相似,具有典型的湖泊、濕地浮游動物群落結構特征(李強等,2015;白海鋒等,2022),而且與濟南地區水域調查結果(種類組成)存在一致性(曹龍智等,2019;劉麟菲等,2019;相華等,2021)。有研究表明在相對靜態的水體中,浮游動物存在小型化趨勢及季節性演替特征,小型浮游動物中的輪蟲類生活史為孤雌生殖,此方式能在較短的時間內達到較高的繁殖率,使其在適宜溫度下種類數量能夠快速增加,從而首先占據更多的生態空間,成為群落優勢種,因此在水域生態系統中浮游動物通常以輪蟲類占主導(Pandey & Upadhyay,2013)。濟西濕地浮游動物趨于小型化,主要與水體的營養變化和魚類的攝食壓力相關,已有研究表明水體富營養化和營養鹽結構的改變使水域浮游植物群落構成趨于小型化,進而通過食物鏈的傳遞效應引起攝食者浮游動物的小型化(Lee et al,2015)。魚類攝食壓力大的水體浮游動物主要以小型輪蟲和小型枝角類為主,而魚類攝食壓力較小的水體以大型橈足類和枝角類占優勢(苗騰等,2013;習麗紅等,2015)。濟西濕地公園每年對觀賞魚類(如錦鯉、金魚等)進行增殖放流,由于其食性為雜食性和濾食性,可通過食物鏈對浮游動物群落的演替產生較大影響。在季節變化中,夏冬季浮游動物種類組成相對較簡單,這主要與夏季人類頻繁的活動增加了水體中營養鹽和有機質含量,以及冬季較低的水溫和夏季持續的降水改變了浮游動物生存環境,進而對浮游生物的群落結構造成影響(趙睿智等,2020)。橈足類無節幼體是濟西濕地公園浮游動物中出現頻率較多的一類,無節幼體主要以水體中懸浮的細菌為食,細菌資源量的多寡直接影響無節幼體的生物量(Lewandowska et al,2014)。調查研究顯示,無節幼體在春夏季占據優勢,其現存量也相對較大,主要因為春秋季適宜的溫度促進了細菌的生長,大量繁殖的細菌為無節幼體提供了豐富食物。
3.2? ?浮游動物功能群特征及多樣性
從時間上看,濟西濕地公園浮游動物功能群季節變化不顯著,春季、夏季和冬季均以功能群RF和PF為主,秋季SCF替代了PF成為了優勢功能群,功能群RF在全年4個季節均出現。浮游動物功能群PF適宜生活在寡污性水體中,在春季、夏季和冬季功能群PF占優勢,說明秋季濕地公園水質相對較好,主要由于秋季水溫很適合大部分藻類的生長繁殖,尤其是硅藻和綠藻。浮游動物功能群SCF以濾食細菌、藻類、有機質和原生動物為主,其濾食效率取決于食物粒徑,尤其與藻類的大小(粒徑)有關。秋季浮游藻類主要以單細胞綠藻為主,其適口性(粒徑范圍2~5 μm)促進了功能群SCF占優勢。在浮游動物群落結構研究中,通常用多樣性指數來判斷水質狀況,多樣性指數、均勻度指數和豐富度指數越高,水生態系統越穩定,水質越好(劉鋼等,2018)。調查顯示,浮游動物多樣性存在季節差異,春季隨著溫度的升高,光照強度增加,浮游生物的生長速度加快,休眠卵漂浮水面孵化,種類數量迅速增加,促進了多樣性提高。在濟西濕地公園的浮游動物優勢種群中,出現了多種富營養水體的耐污種,如臂尾輪蟲、龜甲輪蟲和多肢輪蟲等(白海鋒等,2022),這表明濟西濕地公園水體受到了一定污染,存在富營養化問題。依據營養狀態指數和多樣性指數對濟西濕地公園水體進行評價(金相燦,1995;鄭丙輝等,2007),結果顯示濟西濕地公園水體處于中營養水平,水體污染程度為輕度,其水質狀況相對優于濟南大明湖水體和小清河流域(李瑩,2022)。地理位置差異、自凈能力大小和人類活動干擾強弱對城市水體的水質污染程度和營養化水平都存在很大的影響。
3.3? ?浮游動物功能群與環境因子的關系
水生態系統中環境因子的改變,驅動浮游動物的群落不斷進行適應性演替,借助演替模型可有效預警水環境的變化。本研究典范對應分析表明,水溫是影響濟西濕地公園浮游動物功能群時空分布的主要環境因子之一,這與王帥帥等(2017)對濟南地區主要水域的浮游動物調查研究結果一致。水溫是水域生態系統中影響生物群落結構組成的主要影響因子,水溫的變化對浮游動物的生長、繁殖以及休眠卵的孵化等影響極為顯著(彭水秀等,2019)。此外,水溫通過影響浮游植物的生長和分布,上行效應到浮游動物,驅動浮游動物群落結構和時空分布。本研究顯示浮游動物密度和生物量在春冬季相對較高,這可能是不同類群的浮游動物對水溫的適應性存在差異,適宜的溫度可縮短其發育時間,超出適宜溫度反而會對其生長繁殖產生影響。電導率一般用來反映水中溶解的無機物和有機物含量,與浮游植物的葉綠素濃度具有顯著相關性(Jiang et al,2017),電導率通過影響水體中浮游植物的群落結構來驅動浮游動物的時空分布(陸欣鑫等,2014;Zhao et al,2020)。有學者研究認為,電導率與濕地周邊土地利用類型(居民用地、商業用地等)有極高的相關性,能夠較好地反映出通過徑流進入水體的離子總量,在城鎮化發展較快的地區特別顯著(Ren et al,2003)。調查顯示,電導率春、冬季高,夏、秋季低,與季節溫度變化呈負相關。電導率的季節變化可能與水位及水體的更換量有關,夏、秋季水量充沛,降雨和徑流對水體的離子含量起到了稀釋作用,而春、冬季較高濃度的離子對維持浮游動物細胞的酸堿平衡和滲透壓具有重要影響。磷是浮游植物生長代謝所必需的元素,水體中其含量的升高或降低都會影響浮游植物的生長及群落的演替,通過浮游植物的上行效應間接影響浮游動物的生長及時空分布。濟西濕地公園具有生態服務的功能,其價值大小存在季節性,春、夏、秋季較強的人類活動和相對較多的徑流注入,導致水體中營養鹽含量增加,為浮游動物的生長繁殖提供了有利條件,促進了浮游動物的數量增加,但也導致一些耐污性小型浮游甲殼動物成為優勢種群(溫周瑞和謝平,2013),增加了生物群落結構單一化風險,對濕地水生態和水資源安全構成威脅,這與濟西濕地公園周邊城市快速化發展有關。因此,減少水污染、降低人為干擾、提高濕地生境多樣性是提高生物群落穩定性和保護濕地生態環境的重要舉措。
參考文獻
安睿,王鳳友,于洪賢,等,2017. 三環泡濕地浮游動物功能群季節變化及其影響因子[J]. 生態學報,37(6):1851-1860.
白海鋒,孔飛鶴,王怡睿,等,2021. 北洛河流域浮游動物群落結構時空特征及其與環境因子相關性[J]. 大連海洋大學學報, 36(5):785-795.
白海鋒,宋進喜,龍永清,等,2022. 紅堿淖浮游動物群落結構特征及其與環境因子的關系[J]. 生態與農村環境學報,38(8):1064-1075.
曹龍智,郭偉,王帥帥,等,2019. 黃河流域和淮河流域濟南段浮游動物群落結構多樣性及水質健康評價[J]. 水產科技情報,46(4):230-233,240.
馮世敏,吳明姝,熊蓮,等,2017. 太平湖浮游動物動態演替與環境因子的相關性研究[J]. 水生生物學報,41(3):700-711.
龔森森,李雪梅,吳嘉偉,等,2022. 湖北長湖夏、秋季浮游生物功能群特征及主要影響因子[J]. 水生生物學報,46(5):707-717.
國家環境保護總局,2002. 水和廢水監測分析方法[M]. 北京:中國環境科學出版社.
金相燦,1995. 中國湖泊環境[M]. 北京:海洋出版社.
李強,田華,姜民,等,2015. 淀山湖浮游動物群落結構特征及其影響因子[J]. 水生態學雜志,36(4):69-77.
李瑩,2022. 濟南典型水生態系統浮游生物群落結構及水生態健康評價[D]. 大連: 大連海洋大學.
劉鋼,孟云飛,吳丹,等,2018. 青藏高原可魯克湖浮游動物群落結構特征及水質評價[J]. 大連海洋大學學報,33(3):379-386.
劉麟菲,徐宗學,殷旭旺,等,2019. 濟南市不同區域水生生物與水環境因子的響應關系[J]. 湖泊科學,31(4):998-1011.
陸欣鑫,劉妍,范亞文,2014. 呼蘭河夏秋兩季浮游植物功能分組演替及其驅動因子[J]. 生態學報,34(5):1264-1273.
苗騰,高健,陳炳輝,等,2013. 惠州西湖生態修復對浮游甲殼動物群落結構的影響[J]. 生態科學,32(3):324-330.
彭水秀,張坤,鄧道貴,等,2019. 南漪湖春夏季浮游植物群落結構及其與環境因子的關系[J]. 生物學雜志,36(1):35-38,47.
沈韞芬,章宗涉,龔循矩,等,1990. 微型生物監測新技術[M]. 北京:中國建筑工業出版社.
王家楫,1961. 中國淡水輪蟲志[M]. 北京:科學出版社.
王帥帥,王博涵,金文,等,2017. 濟南地區豐水、枯水期浮游動物的群落結構特征[J]. 水產學雜志,30(2):26-30.
溫周瑞,謝平,2013. 太湖梅梁灣、貢湖灣水質與浮游甲殼動物密度季節變化[J]. 長江大學學報(自然科學版),10(17):36-51.
習麗紅,李慧明,林秋奇,等,2015. 熱帶富營養水庫敞水區浮游動物群落結構與季節變化:以廣東大沙河水庫為例[J]. 湖泊科學,27(6):1049-1058.
相華,賈麗,馮新華,等,2021. 濟南地區浮游動物群落結構與環境因子關系研究[J]. 人民黃河,43(S1):80-82,84.
趙睿智,趙紅雪,邱小琮,2020. 黑河干流浮游動物與水環境因子關系的多元分析[J]. 水生態學雜志,41(6):81-88.
趙文,2005. 水生生物學[M]. 北京:中國農業出版社.
鄭丙輝,田自強,張雷,等,2007. 太湖西岸湖濱帶水生生物分布特征及水質營養狀況[J]. 生態學報,27(10):4214-4223.
朱浩,劉興國,陳曉龍,等,2020. 鰣鯸淀浮游動物群落特征及水質評價[J]. 水生生物學報,44(6):1313-1321.
Allain J B, Kerri F, Beatrix E B, 2007. Functional diversity of crustacean zooplankton communities: towards a trait-based classification[J]. Freshwater Biology,52:796-813.
Elijah O O, Mucai M, Charles C N, et al, 2011. Zooplankton partitioning in a tropical alkaline-saline endorheic Lake Nakuru, Kenya: Spatial and temporal trends in relation to the environment[J]. Lake & Reservois: Research and management, 16:35-47.
Hoeinghaus D J, Winemiller K O, Agostinho A A, 2007. Landscape-scale hydrologic characteristics differentiate patterns of carbon flow in large-river food webs[J]. Ecosystems,10(6):1019-1033.
Jiang W, Zhou C, Ji D B, et al, 2017. Comparison of relationship between conduction and algal bloom in Pengxi River and Modao River in Three Gorges Reservoir[J]. Environmental Science,38(6):2326-2335.
Lee Y C, Park M O, Jung J, et al, 2015. Taxonomic variability of phytoplankton and relationship with production of CDOM in the polynya of the Amundsen Ses, Antarctica[J]. Deep-Sea Research II,123:30-41.
Lewandowska A M, Hillebrand H, Lengfellner K, et al, 2014. Temperature Effects on Phytoplankton Diversity: The Zooplankton Link[J]. Journal of Sea Research,85:359-364.
Pandey A, Upadhyay H C, 2013. Studies on zooplankton diversity of western Ramganga River in Almora (Uttarakhand) India[J]. Int J Sci Res,5(5):1192-1194.
Ren W W, Zhong Y, Meligrana J, et al, 2003. Urbanization, landuse, and water quality in Shanghai 1947-1996[J]. Environment International,29:649-659.
Shannon C E,1948. The mathematical theory of communication [J]. Bell System Technical Journal,27(3):379-423.
Wang M Y, Liu R H, Wang Y, et al, 2019. Temporal and spatial distribution of main contaminations and eutrophication assessment in wetlands of Ziya River Basin[J]. Periodical of Ocean University of China,49:93-100.
Zhao G N, Pan B Z, Li Y P, et al, 2020. Phytoplankton in the heavy sediment-laden Weihe River and its tributaries from the northern foot of the Qinling Mountains: Community structure and environmental drivers[J]. Environmental Science and Pollution Research International,27(8):8359-8370.
(責任編輯? ?張俊友? ?熊美華)
Zooplankton Functional Groups and Their Response to Water Physiochemical
Factors in Jixi Wetland Park in Jinan
SHANG Shu‐qin1, JIA Li1, WANG Shuai‐shuai1, YIN Xu‐wang2, BAI Hai‐feng3,4
(1. Jinan Hydrology Center, Jinan 250014, P.R. China;
2. College of Fisheries and Life Science, Dalian Ocean University, Dalian? ?116023, P.R. China;
3. Shaanxi Institute of Geo-environment Monitoring, Xi'an? ?710054, P.R. China;
4. Yellow River Institute of Shaanxi Province, Xi'an? ?710127, P.R. China)
Abstract:Jixi wetland park is a typical urban wetland located in the urban area of west Jinan City, providing drinking water for Jinan City, flood control and ecological conservation. In this study, we explored the characteristics of zooplankton functional groups as well as their relationship with the physiochemical factors in Jixi wetland park. The objectives were to understand the structure and function of the wetland ecosystem and provide theoretical support for evaluating and managing water quality in Jixi wetland. From April 2020 to March 2021, zooplankton sampling and environmental monitoring was conducted monthly at six representative sampling sites (JX1-JX6) in Jixi wetland. A total of 66 zooplankton species were collected during the investigation and divided into 7 functional groups (PF: protozoan filter feeders, RF: rotifer filter feeders, RC, rotifer carnivores; SCF: small copepods and cladocera filter feeders, MCF: middle copepods and cladocera filter feeders, MCC: middle copepods and cladocera carnivores, LCF: large copepods and cladocera carnivores). Zooplankton density and biomass were in the ranges of 16.0 -824.0 ind/L and 0.004 -1.667 mg/L, with average annual values of 181.8 ind/L and 0.210mg/L. Groups RF (rotifer filter feeders) and PF (protozoan filter feeders) were dominant in spring, summer and winter, and the highest proportion of group RF (62.86%) was in winter, and the highest proportion of group PF (20.75%) was in spring, and group RF (rotifers filter feeders) (58.54%) and SCF (small copepods and cladocera filter feeders) (12.20%) were dominant in autumn. The mean value of the Shannon-Wiener diversity index, Pielou evenness index and Margalef richness index of the zooplankton community were 2.504, 0.708 and 2.307, respectively. The water quality was evaluated using the comprehensive trophic level index and the results show that the water quality of Jixi wetland was mesotrophic. Canonical correspondence analysis shows that WT, COND and PO43- were the environmental factors significantly affecting zooplankton functional groups. This study will help guide ecological effect assessment and water resource protection of Jixi wetland.
Key words:zooplankton; functional groups; diversity assessment; Jixi wetland park
收稿日期:2022-09-27? ? ? 修回日期: 2023-06-19
基金項目:國家自然科學基金項目(41977193)。
作者簡介:商書芹,女,1978年生,高級工程師,從事水文生態學研究。E-mail: 316796647@qq.com
通信作者:殷旭旺。E-mail: yinxuwang@dlou.edu.cn
白海鋒。E-mail: baihaifeng2002@126.com
