水稻–冬季秸稈覆蓋種植馬鈴薯模式對水稻齊穗期根際土壤代謝組表達特征的影響①
2023-05-26 02:52:26王孟佳馮向前馬橫宇劉元輝徐春梅章秀福王丹英
土壤 2023年2期
王孟佳,馮向前, 2,馬橫宇,劉元輝,褚 光,徐春梅,章秀福,王丹英,陳 松*
水稻–冬季秸稈覆蓋種植馬鈴薯模式對水稻齊穗期根際土壤代謝組表達特征的影響①
王孟佳1,馮向前1, 2,馬橫宇1,劉元輝1,褚 光1,徐春梅1,章秀福1,王丹英1,陳 松1*
(1水稻生物學國家重點實驗室(中國水稻研究所),杭州 311400;2長江大學農學院,湖北荊州 434025)
水稻–冬季秸稈覆蓋種植馬鈴薯是南方稻區(qū)一種經濟高產的水–旱輪作模式,其對水稻根際土壤代謝物的影響可能是調控水稻高產的關鍵,但目前相關研究尚未見報道。本研究基于周年水–旱輪作模式長期定位試驗,以水稻–冬季空閑(CK)模式為對照和水稻–馬鈴薯(稻草覆蓋種植并還田,RP)輪作模式為研究對象,采集水稻齊穗期根際土壤樣品,利用液相色譜–質譜法(LC-MS)測定根際土壤代謝物種類與豐度,研究RP輪作模式水稻齊穗根際土壤代謝物組分和功能及其與產量形成的關系。結果表明:①采用OPLS-DA模型篩選差異代謝物(HMDB數(shù)據(jù)庫)發(fā)現(xiàn),與CK模式相比,RP輪作模式具有201個有顯著性差異(<0.05)的根際土壤代謝物,分別歸屬于脂質和類脂分子、有機酸及衍生物、有機氧化合物、苯丙烷和聚酮等11個父類;其中苯丙烷和聚酮、丙烯醇脂質、苯及其衍生物、甾體內酯、氨基酸/肽、碳水化合物和脂肪酰糖苷等代謝物豐度發(fā)生顯著變化,可作為表征RP輪作模式下水稻根際土壤差異性代謝物;②有機氧化合物、有機酸及衍生物和苯丙烷和聚酮等代謝物豐度與土壤有機質、全鉀、堿解氮和速效鉀含量差異呈顯著正相關(=0.816 ~ 0.938),其可通過為水稻直接提供養(yǎng)分、協(xié)助提高土壤養(yǎng)分供給或間接調節(jié)水稻生長等提高稻田綜合生產力;③核苷/核苷酸、脂質和類脂分子等差異代謝物與水稻產量差異呈顯著正相關(=0.957 ~ 0.999),其可能是通過參與一系列代謝途徑提高水稻抗逆能力,最終對水稻產量產生一定影響。
水稻根際;代謝組;輪作;秸稈還田
冬季稻草覆蓋種植馬鈴薯-水稻輪作是一種生態(tài)經濟的種植模式,在有效利用資源和養(yǎng)分的前提下,有助于作物穩(wěn)產高效[1],因而在生產上受到廣大農戶歡迎。該模式集成冬季作物輪作與秸稈覆蓋和翻耕還田,不僅可以有效提高土壤理化性質,還能顯著改善土壤環(huán)境和微生物活性,有利于土壤養(yǎng)分的釋放和作物吸收利用,從而提高作物產量[2],但目前對其穩(wěn)產高效的土壤調控機理研究較少。另外,大量秸稈還田(即外源有機碳添加)為土壤提供復雜且穩(wěn)定的有機質組分[3],其活性有機質組分已被證實不僅可以培育各種土壤生物,協(xié)調與促進各項土壤生理生化過程,調控土壤各組分周轉,還是協(xié)調有機質礦化和作物養(yǎng)分需求的關鍵角色[4-6]。但土壤的時空不均衡性,決定了農田土壤會隨著作物生長形成一個影響稻田有機質轉化的特異性熱區(qū)[7],即水稻根際(rhizosphere)。該區(qū)域根系活躍且相對獨立,不僅影響水稻對營養(yǎng)元素的吸收,還會通過累積的代謝產物導致根際效應[8],進而影響周圍環(huán)境與作物生長。因此,輪作變更導致的根際土壤代謝物組成與功能轉變可能是解析冬季稻草覆蓋種植馬鈴薯–水稻輪作模式穩(wěn)產高效的關鍵。
水稻根際,一般指水稻根系周圍直徑約4 mm、受根系活動影響的土壤區(qū)域。該區(qū)域代謝物組分包含根系分泌物和經根際土壤微生物代謝后產生的各類小分子物質,富含糖類、有機酸等化合物,對周圍土壤、根際微生物、作物本身的生長發(fā)育及其抗病和抗逆能力等均會產生一定影響,還會觸發(fā)根際土壤有機質的分解與轉化[9]。水稻生長中后期(尤其是穗發(fā)育期)根系活性特征,例如根系生長能力、根長、根體積、根部生物量及根際微生物益生菌群等方面對于產量形成具有重大意義[10],而根際的活性有機代謝物則可能是影響以上根系活力的重要因素。因此,探究水稻穗發(fā)育期根際土壤活性有機代謝物特征至關重要。但是根際土壤中代謝物組分和豐度除受病原真菌侵害[11]、外界脅迫[12]、植物生長調節(jié)劑[13]等因素的調控外,還在較大程度上與作物輪作模式密切相關[14]。近年來隨著科學技術的發(fā)展,有學者發(fā)現(xiàn)輪作可通過變更土壤環(huán)境對作物根際土壤代謝物組成與豐度產生影響。例如Lu等[15]利用氣相色譜-質譜法(GC-MS)發(fā)現(xiàn)水稻-水稻-休耕和水稻-水稻-油菜輪作模式下根際土壤代謝物如烷烴、有機酸和苯類等物質豐度顯著不同;而康益晨等[16]對馬鈴薯研究發(fā)現(xiàn)與馬鈴薯連作相比,輪作蠶豆可減少根系分泌物中酯類物質,增加烴類、酮類、羧酸及胺類等物質。由此可見,不同輪作模式下作物根際土壤代謝物種類及豐度存在較大差異,了解不同輪作條件下根際土壤代謝物的表現(xiàn)特征對于維持穩(wěn)定的輪作制度至關重要。然而目前有關根際土壤中各種功能性代謝物對輪作模式的響應的報道較少,其中對水稻-秸稈覆蓋種植馬鈴薯輪作模式下水稻根際土壤代謝物變化的研究尚未見報道。
根際土壤代謝組是近年來新興的聚焦根際土壤代謝物的高通量組學篩選技術,利用液相色譜-質譜法(LC-MS)可對系統(tǒng)內所含的小分子代謝物進行半定量分析,探究根際代謝物與作物生理-土壤養(yǎng)分變化間的關系,揭示系統(tǒng)內生理生態(tài)功能的狀態(tài),已廣泛應用于微生物研究等領域[17]。本研究以2003年在浙江富陽開始的水旱輪作模式長期定位試驗為基礎,以水稻-冬季空閑(CK)模式為對照、水稻-馬鈴薯(稻草覆蓋種植并還田,RP)輪作模式為處理,采用“根袋法”采集水稻齊穗期根際土壤,利用LC-MS法研究長期RP輪作模式下,水稻齊穗期根際土壤差異代謝物的表現(xiàn)特征及其與水稻產量的潛在關系,以為水稻-馬鈴薯輪作模式穩(wěn)產高效種植提供理論依據(jù)。
1 材料與方法
1.1 試驗地概況
試驗于2020年在浙江省富陽市中國水稻研究所試驗農場(120.2°E,30.3°N,海拔11 m)進行。試驗地屬亞熱帶季風氣候區(qū),年均溫約18 ℃,年日照時數(shù)1 500 h以上,年均降水量1 700 mm左右。2020年移栽前CK模式土壤有機質32.10 g/kg,全氮2.30 g/kg,堿解氮125.22 mg/kg,有效磷30.90 mg/kg,速效鉀145.08 mg/kg,pH 5.88;RP輪作模式土壤有機質42.40 g/kg,全氮2.50 g/kg,堿解氮137.07 mg/kg,有效磷102.56 mg/kg,速效鉀320.84 mg/kg,pH 5.79。
1.2 試驗設計
本研究基于以水稻為主體的多種輪作模式結合冬季作物殘體還田的長期定位試驗(2003年至今),該長期定位試驗采用裂區(qū)隨機區(qū)組設計,以6種不同輪作模式為主區(qū),水稻季不同施氮水平為副區(qū),主區(qū)面積為160 m2(8 m×20 m),副區(qū)面積40 m2(8 m×5 m),3個田間區(qū)組重復,小區(qū)面積及位置常年固定。供試品種為嘉58,6月中旬手工移栽,行株距20 cm×20 cm,每穴2 ~ 3株,移栽前使用小型耕田機將冬作及秸稈殘留還田。
本研究選擇以水稻-冬閑(CK)模式為對照、水稻–稻草覆蓋種植馬鈴薯(RP)輪作模式作為研究對象,其2018—2020年水稻產量及還田秸稈特性見表1,可以看出,RP輪作模式下水稻產量平均較CK模式升高18.01%。在前期研究的基礎上,本研究再選擇以水稻生育期不施氮(N0)處理作為研究對象,試驗處理具體如下:在CK處理中,上茬水稻收割后移除全部秸稈,冬季閑田放置;在RP處理中,1月下旬田面擺放種薯后,覆蓋稻草8 ~ 10 cm厚,施復合肥(N︰P2O5︰K2O=15︰15︰15)600 kg/hm2,5月中旬收獲塊莖后將腐爛的稻草及馬鈴薯莖稈和根系殘留翻耕還田。水稻生育期間不施氮肥,磷、鉀肥分別以過磷酸鈣和氯化鉀為主,施用量分別為P2O597.5 kg/hm2和K2O 165 kg/hm2,磷肥全部基施,鉀肥基︰穗肥為5︰5。其他田間栽培措施與當?shù)馗弋a栽培一致。
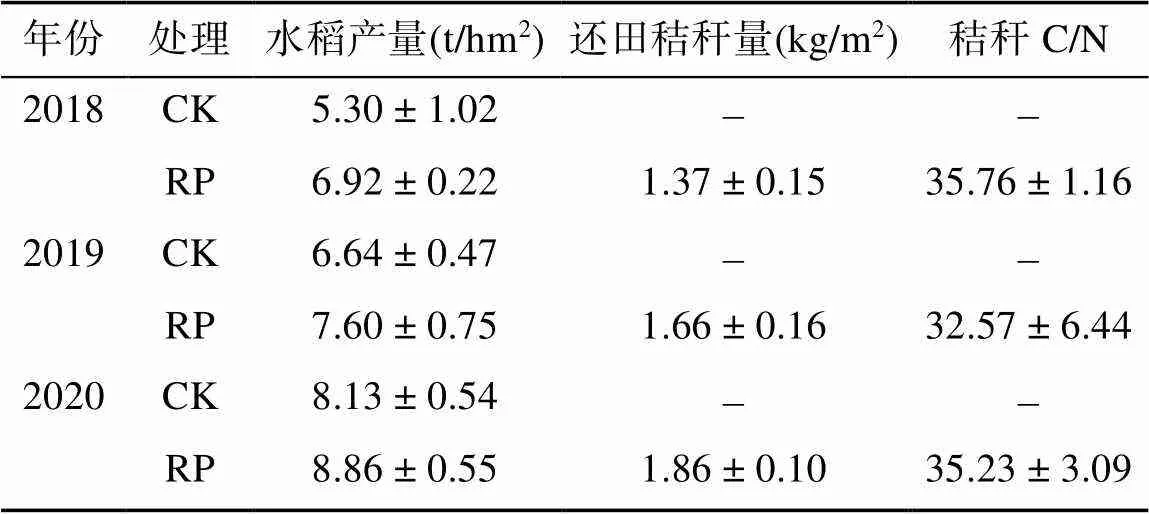
表1 近3年水稻產量及還田秸稈特性
1.3 樣品采集與測定分析
1.3.1根際土壤的采集 于水稻移栽一周后使用PVC柱(直徑20 cm)將水稻植株及根區(qū)(0 ~ 20 cm)土壤挖出放入根袋(200目,直徑20 cm,深30 cm),再次埋入田中原位培養(yǎng)。至齊穗期挖出根袋及水稻,小心撕下根袋維持水稻根系完整,用力抖動水稻去除根部大部分土壤,刮取附著于根表的土壤即根際土,去除雜質后保存于–80 ℃冰箱待測。每個處理設3個田間重復,2個實驗重復,共計6個重復。
1.3.2 根際土壤代謝物測定 樣本處理如下:樣本解凍后稱取1 000 mg于2 mL離心管,加入6 mm研磨珠和1 000 μL甲醇/水(4︰1,/)提取液,經冷凍組織研磨儀研磨6 min(–10℃,50 Hz)后低溫超聲提取30 min(5 ℃,40 kHz),于–20℃靜置30 min,離心15 min(13 000 g,4℃)后取上清液氮氣吹干。在分析代謝物之前,取100 μL的乙腈/水(1︰1,/)重新溶解樣本,重復上述低溫超聲提取和離心步驟后取上清液上機分析。每個樣本分別移取20 μL上清液,混合后作為質控樣本(QC)。本試驗采用AB SCIEX公司的超高效液相色譜串聯(lián)飛行時間質譜UHPLC- Triple TOF系統(tǒng)對CK和RP處理根際代謝物進行分析。色譜條件:色譜柱為ACQUITY UPLC HSS T3(100 mm × 2.1 mm i.d.,1.8 μm;Waters,Milford,USA);流動相A為95% 水+5% 乙腈(含0.1% 甲酸),流動相B為47.5% 乙腈+ 47.5% 異丙醇+ 5 %水(含0.1% 甲酸);流速為0.40 mL/min,進樣量為10 μL,柱溫為40 ℃。在儀器分析的過程中,每5個分析樣本中插入一個QC樣本,以考察整個檢測過程的穩(wěn)定性。采用電噴霧電離法(ESI)分別采用正、負離子掃描模式采集質譜信號。質譜參數(shù)見表2。
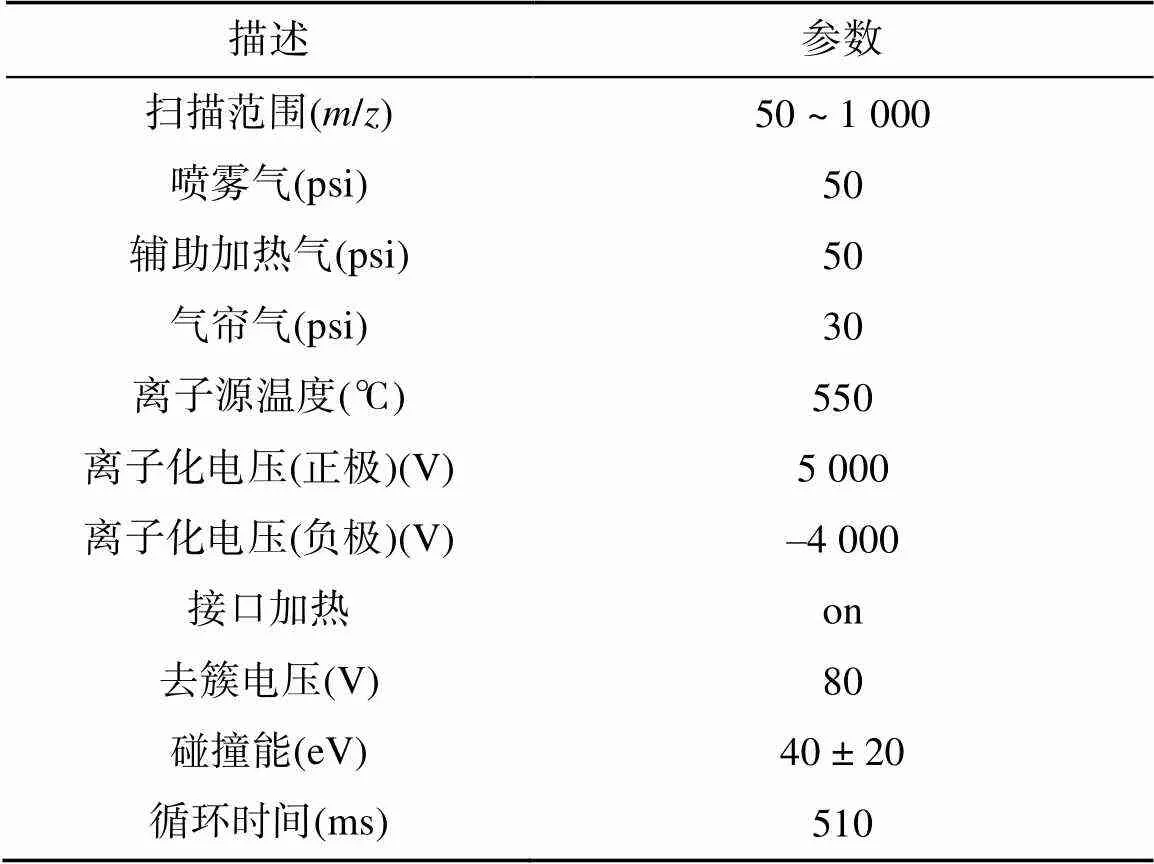
表2 質譜參數(shù)
1.4 數(shù)據(jù)整理與分析
本試驗所有數(shù)據(jù)均使用Microsoft Excel 2010軟件進行整理,采用代謝組學軟件Progenesis QI進行基線過濾、峰識別、積分、保留時間校正、峰對齊,最終得到一個含保留時間、峰強度等的數(shù)據(jù)矩陣,保留至少一組樣品中非零值80% 以上的變量,再用原始矩陣中最小值填補空缺值,并利用總和歸一化法對樣本質譜峰的響應強度進行歸一化處理;同時刪除QC樣本相對標準偏差(RSD)>30% 的變量,并進行對數(shù)化(lg)處理,得到最終用于后續(xù)分析的數(shù)據(jù)矩陣;通過特征峰搜庫鑒定,將MS和MS/MS質譜信息與HMDB 4.0(https://hmdb.ca/)和KEGG Release 2017- 05-01(https://www.genome.jp/kegg/)代謝數(shù)據(jù)庫進行匹配,MS質量誤差設置為小于10–6,同時根據(jù)二級質譜匹配得分鑒定代謝物;采用R語言(1.6.2)進行多元統(tǒng)計分析,并對數(shù)據(jù)進行主成分分析(PCA)和正交偏最小二乘法分析(OPLS-DA),PCA和OPLS- DA置信度0.95,OPLS-DA置換次數(shù)為200;單因素方差分析(ANOVA,value)繪制火山圖;基于HMDB和KEGG數(shù)據(jù)庫,鑒定、篩選、歸類組間差異(<0.05)代謝物,并采用OriginPro 2021對其繪圖;基于KEGG Pathway Database(https://www.genome. jp/kegg/pathway.htmL)進行通路富集分析并繪圖(https://www.chiplot.online/)。
2 結果
2.1 根際土壤代謝物的質譜圖譜
正、負離子模式下12個土壤樣本分別鑒定出6 041和5 544個有效峰,經處理后最終保留5 641和5 260個有效峰(圖1)。由圖1可以看出,總離子色譜峰形良好,分布相對均勻。基于HMDB和KEGG數(shù)據(jù)庫,共鑒定到可命名代謝物661個,其中HMDB數(shù)據(jù)庫可鑒定到598個,KEGG數(shù)據(jù)庫172個。
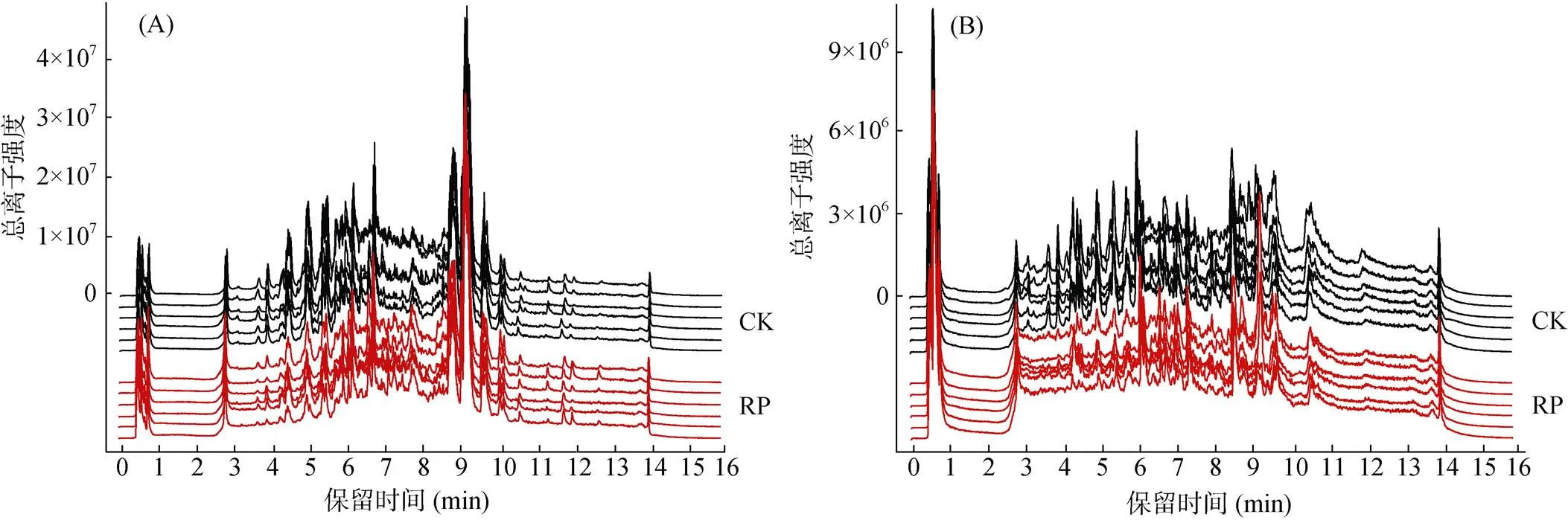
圖1 正離子(A)和負離子(B)模式下12個土樣的重疊總離子流色譜圖
2.2 基于土壤代謝組的根際土壤代謝物主成分分析
對正、負離子模式下CK和RP處理代謝物進行主成分分析(圖2A、2B),可見,CK和RP處理組間差異明顯,樣本間坐標點較為聚集,易于區(qū)分,所有坐標點均位于置信橢圓內,表明各樣本均處于95% 置信度下,且組內樣本相似性較高,數(shù)據(jù)處理可信度高。
2.3 根際土壤組間差異代謝物的篩選
采用OPLS-DA方法結合VIP值(>1)和差異表達倍數(shù)(fold change,F(xiàn)C >1或< 1),基于HMDB數(shù)據(jù)庫對處理間差異代謝物進行篩選,共鑒定出201個具有顯著性差異(<0.05)的代謝物,相對CK處理,RP處理中表達差異上調的代謝物112個,表達差異下調的代謝物89個(圖3)。差異代謝物分屬于脂質和類脂分子、有機酸及衍生物、有機氧化合物、苯丙烷和聚酮、有機雜環(huán)化合物、苯環(huán)型化合物、核苷/核苷酸和類似物等11個父類(圖4A)。

圖2 正離子(A)和負離子(B)模式下CK和RP處理組的PCA得分圖
輪作處理后根際代謝物中變化最多的是脂質和類脂分子,共含85個差異代謝物,但其豐度提高和降低約各占一半(圖4A),其中豐度下降的代謝物主要集中在脂肪酰糖苷、脂肪醇、脂肪醇酯、甘油單油酸酯、膽汁酸、醇和衍生物等子類;豐度提高的丙烯醇脂質以子類代謝物三萜類、萜苷和二萜類為主,另外甾體內酯子類代謝物豐度也明顯提高(圖4C)。有機酸及衍生物中共有24個差異代謝物,僅次于脂質和類脂分子,其中豐度增加的代謝物主要集中在羧酸及其衍生物中的氨基酸/肽和類似物,而下調代謝物則包括酮酸及其衍生物、有機硫酸及其衍生物和有機硫代磷酸及其衍生物(圖4B)。在差異性有機氧代謝物中,碳水化合物及其結合物豐度明顯增加(圖4C)。此外,豐度明顯提高的還包括苯丙烷和聚酮類化合物(包括類黃酮、異黃酮類、苯丙酸、茋類化合物、肉桂酸及其衍生物等)、有機雜環(huán)化合物(吡啶及其衍生物等)和苯及其取代衍生物等;而豐度下降的主要是嘌呤核苷酸糖類代謝物(圖4)。
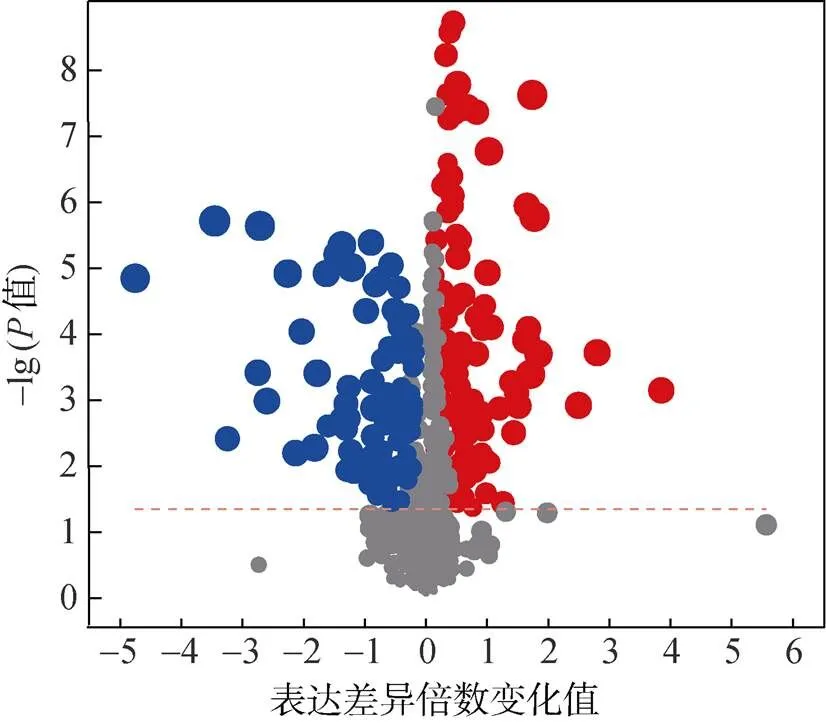
(每個點代表一個特定的代謝物;點的大小表示VIP值;紅色表示表達差異上調;藍色表示表達差異下調;灰色表示無顯著差異)
2.4 根際土壤差異代謝物代謝通路
將輪作處理后的201個差異代謝物對比KEGG Pathway數(shù)據(jù)庫,發(fā)現(xiàn)有27個差異代謝物分別注釋在28條可能的通路上(表3)。其中,有7個差異代謝物參與各種脂肪酸代謝途徑,如花生四烯酸代謝、亞油酸代謝、α-亞麻酸代謝和丁酸代謝,其富集代謝物豐度存在差異。4個差異性表達的核苷/核苷酸和類似物類代謝物中,有3個除參與嘌呤或嘧啶代謝有關的途徑外,還參與信號轉導相關的途徑,如ABC轉運子、cGMP-PKG信號通路和cAMP信號通路途徑;此外,除參與果糖和甘露糖代謝以及氨基糖和核苷酸糖代謝的GDP-4-Dehydro-6-deoxy-D-mannose豐度下調外,其他代謝物豐度均表現(xiàn)為提高。另有3、3和2個代謝物分別參與到類固醇合成降解代謝(脂質和類脂分子及其他)、戊糖和葡萄糖醛酸轉化途徑(有機氧化合物以及脂質和類脂分子)、葉酸生物合成以及煙酸和煙酰胺代謝等代謝途徑(有機雜環(huán)化合物),且后3條代謝通路涉及的5個代謝物均表現(xiàn)為豐度提高。此外,差異性表達代謝物還與萜類代謝、谷氨酰胺與谷氨酰胺代謝、氨基苯甲酸降解、甘油磷脂代謝等相關。利用拓撲學算法“Relative-betweeness centrality”對相關差異代謝物進行代謝通路富集分析,發(fā)現(xiàn)共計9條代謝途徑顯著富集(<0.05,圖5),包括信號傳導途徑和嘌呤類代謝、脂肪酸(丁酸、亞油酸、亞麻酸)代謝、類固醇激素生物合成等。

表3 KEGG代謝物涉及功能通路匯總

(富集因子為富集在該通路的代謝物個數(shù)與富集在28條通路中總代謝物個數(shù)27的比值)
總體而言,水稻-馬鈴薯輪作后水稻根際土壤中差異代謝物涉及的代謝通路中脂肪酸類代謝最多,且富集到各通路的代謝物個數(shù)及豐度不同。參與嘌呤和嘧啶類代謝、ABC轉運子、戊糖和葡萄糖醛酸轉化等途徑的代謝物均上調,而參與生物堿的生物合成等途徑的代謝物則下調。
3 討論
3.1 秸稈覆蓋種植馬鈴薯–水稻輪作處理下根際土壤代謝物差異的基本特征
根際土壤代謝物是植物根系-土壤交流的過程中,植物及與之互作的土壤微生物分泌的各種有機無機化合物和信號分子,包含各種光合產物中的初級和次級代謝物[18],在不同作物栽培措施下會表現(xiàn)出不同的反應[16]。Lu等[15]和康益晨等[16]發(fā)現(xiàn)作物輪作可顯著影響有機酸、苯類、酯類、烴類、酮類及胺類等代謝物;Li等[19]在研究秸稈還田時,利用1H-NMR技術發(fā)現(xiàn)秸稈還田后玉米根際土壤氨基酸、糖、低分子有機酸等發(fā)生明顯變化;陳軍等[20]采用GC-MS分析不同氮肥運籌對水稻根際土壤代謝物的影響,發(fā)現(xiàn)氮肥調控的根際差異代謝物主要為醇類、酚類、醚類、脂類、酸類以及酮類。本研究利用LC-MS分析RP處理后水稻根際土壤代謝物的差異性變化,發(fā)現(xiàn)與CK處理相比,RP處理下的差異性代謝物主要集中在脂質和類脂分子、有機酸及衍生物和有機氧化合物等11個父類,對比HMDB庫,主要以苯丙烷和聚酮、丙烯醇脂質、苯及其取代衍生物、甾體內酯、氨基酸/肽和類似物、碳水化合物及其結合物和脂肪酰糖苷物質等為主,而土壤有機酸等變化不大。這些差異可能與所采用的種植制度及其所搭配的栽培措施有關。相較于CK處理,RP處理中水稻與馬鈴薯輪作顯著改變了根際土壤代謝物的種類及豐度;其次,該輪作系統(tǒng)中所涉及的馬鈴薯季秸稈覆蓋與還田、施肥及馬鈴薯殘茬還田等措施可為土壤及水稻提供一定的氮素養(yǎng)分,從而使脂類等受氮肥影響的土壤代謝物發(fā)生變化[20]。因此,苯丙烷和聚酮等差異性代謝物是RP模式下水稻根際土壤的特征指標之一,可為鑒定與區(qū)分RP輪作提供一定參考。
3.2 根際土壤代謝物在稻田綜合生產力中的潛在功能
前期研究表明,周年水稻-馬鈴薯秸稈覆蓋輪作模式有利于水稻高產潛力的實現(xiàn),是一種穩(wěn)產高效的輪作模式[2,21]。本研究中,通過對CK和RP處理間差異代謝物豐度與養(yǎng)分差異進行相關性分析發(fā)現(xiàn),差異代謝物苯丙烷和聚酮、有機氧化合物、有機酸及衍生物等的豐度與土壤有機質、全鉀、堿解氮、速效鉀含量差異呈顯著正相關(=0.816 ~ 0.938,表4)。基于該結果,本研究認為根際土壤代謝物對稻田綜合生產力的作用可能源于:①根際土壤代謝物可直接為作物提供營養(yǎng)物質。前人在研究秸稈還田和外源物添加時,發(fā)現(xiàn)土壤根際的氨基酸、糖、低分子有機酸以及銨態(tài)氮含量等均發(fā)生不同程度的變化[19,22],與之類似,本研究也發(fā)現(xiàn)RP輪作體系下水稻根際土壤中從屬于苯丙烷和聚酮、有機氧化合物、有機酸及衍生物的類黃酮、碳水化合物、氨基酸/肽等豐度顯著提高(圖4),而小分子氨基酸/多肽等可作為養(yǎng)分被作物直接吸收,進而提高作物生產力;②通過調控土壤自養(yǎng)型氮礦化微生物,提高土壤供肥能力。源于光合作用的根際碳被認為是土壤養(yǎng)分和作物生產間的重要媒介,作為外源碳,可激發(fā)土壤固有碳氮礦化分解,促進土壤養(yǎng)分的供給[23]。因此,RP輪作后水稻根際土壤糖類豐度增加,有助于調控相關養(yǎng)分挖掘型微生物的增加,從而誘導土壤固有有機質的激發(fā),最終導致稻田養(yǎng)分供給能力的提高。但相關機理與研究存在很大爭議,仍需進一步探索與驗證;③根際促生菌(plant growth-promoting rhizobacteria,PGPR)的間接作用。根際促生菌是一類存在于植物根系附近的調節(jié)作物生長、緩解逆境脅迫的微生物,其在土壤中的作用與根際土壤代謝物組分密切相關[24]。其中,碳水化合物與氨基酸是作物和PGPR之間共生關系的媒介[25],共同作用于PGPR對植物的調控過程。此外,黃酮類等次級代謝物也是PGPR誘導作物促進生長的抵御病原菌攻擊的重要物質[26]。因此,RP輪作后,苯丙烷和聚酮、有機氧化合物、有機酸及衍生物等土壤代謝物不僅直接或間接為水稻提供養(yǎng)分物質,還間接調控根際促生微生物的群落與結構,調節(jié)水稻生長,最終影響稻田綜合生產力。

表4 養(yǎng)分含量差異與差異代謝物間的相關性
注:*、**分別表示在<0.05、<0.01水平顯著相關,下同。
3.3 根際土壤代謝物在提高水稻抗逆能力方面的潛在功能
RP處理中的特異性根際土壤代謝物對水稻生長發(fā)育的影響,還可能以信號分子的形式增強作物抗逆能力。植物與外界的交流、互作依靠各種信號分子;在外界環(huán)境變化時,PGPR與植物依賴這些物質抵御外界脅迫。本研究鑒定到4種核苷/核苷酸和有機酸類代謝物顯著富集到3條信號轉導途徑,即ABC轉運子、cGMP-PKG和cAMP信號轉導途徑(圖5)。其中ABC轉運子主要存在于質膜和液泡膜,參與生物降解或糖類中間產物的運輸?shù)榷喾N生理過程,使植株適應不斷變化的外界環(huán)境以抵抗各種生物和非生物脅迫[27]。cAMP信號通路是環(huán)核苷酸系統(tǒng)的一種,可調節(jié)植物對生理和環(huán)境刺激的響應。cGMP-PKG信號通路可作用于生物體內一氧化氮(NO)信號轉導過程,通過調控NO提高水稻的耐鹽性、耐熱性以及水稻根系對重金屬毒害的抗性[28]。除信號分子外,還有一些脂質和類脂分子參與植物抗逆防御的代謝途徑,顯著富集于亞油酸/亞麻酸代謝、類固醇激素合成等。亞油酸代謝產物褪黑素可在水稻受到鹽脅迫時通過增強葉綠素競爭、維持膜穩(wěn)定性保障其正常生長[29];亞油酸和亞麻酸及其衍生物可進一步轉化為氧化脂質,在水稻抗病過程中作為信號分子和抗菌化合物抑制病原體生長[30]。此外,本研究還發(fā)現(xiàn)核苷/核苷酸及脂質和類脂分子等差異代謝物豐度與水稻產量差異呈顯著正相關(= 0.957 ~ 0.999,表5)。因此,長期RP輪作后水稻根際土壤中核苷/核苷酸及脂質和類脂分子等代謝物通過參與信號傳導及植物抗逆、防御等相關代謝途徑,提高水稻應對外界生物和非生物脅迫的能力,保障水稻產量的形成。

表5 水稻產量差異與差異代謝物間的相關性
4 結論
與水稻-冬閑模式相比,水稻-秸稈覆蓋種植馬鈴薯模式下水稻齊穗期根際土壤特異性代謝物主要以苯丙烷和聚酮、丙烯醇脂質、苯及其取代衍生物、甾體內酯、氨基酸/肽和類似物、碳水化合物及其結合物和脂肪酰糖苷物質等為主,這些差異性代謝物及其豐度可作為鑒別RP輪作的重要參考指標。RP輪作后,其特有的根際土壤代謝物不僅直接或間接為水稻提供養(yǎng)分物質,還通過間接調控根際土壤促生微生物促進水稻生長,影響水稻產量表現(xiàn);另一方面,相關代謝物通過參與信號傳導及植物抗逆、防御等相關代謝途徑,提高水稻應對外界生物和非生物脅迫的能力,從而保障水稻高產。
[1] 謝小燕. 稻田水旱輪作栽培模式效益淺析[J]. 現(xiàn)代農業(yè)研究, 2021, 27(2): 98–100.
[2] Chen S, Xu C M, Yan J X, et al. The influence of the type of crop residue on soil organic carbon fractions: An 11-year field study of rice-based cropping systems in southeast China[J]. Agriculture, Ecosystems & Environment, 2016, 223: 261–269.
[3] 王虎, 王旭東, 田宵鴻. 秸稈還田對土壤有機碳不同活性組分儲量及分配的影響[J]. 應用生態(tài)學報, 2014, 25(12): 3491–3498.
[4] 張維理, Kolbe H, 張認連. 土壤有機碳作用及轉化機制研究進展[J]. 中國農業(yè)科學, 2020, 53(2): 317–331.
[5] Kuzyakov Y, Friedel J K, Stahr K. Review of mechanisms and quantification of priming effects[J]. Soil Biology and Biochemistry, 2000, 32(11/12): 1485–1498.
[6] Kemmitt S J, Lanyon C V, Waite I S, et al. Mineralization of native soil organic matter is not regulated by the size, activity or composition of the soil microbial biomass—a new perspective[J]. Soil Biology and Biochemistry, 2008, 40(1): 61–73.
[7] 孫悅, 徐興良, Yakov K. 根際激發(fā)效應的發(fā)生機制及其生態(tài)重要性[J]. 植物生態(tài)學報, 2014, 38(1): 62–75.
[8] 徐國偉, 常二華, 陳明燦, 等. 根系分泌物對水稻及其他作物生長與品質影響[J]. 湖北農業(yè)科學, 2011, 50(23): 4757–4760.
[9] Blagodatskaya E, Littschwager J, Lauerer M, et al. Plant traits regulating N capture define microbial competition in the rhizosphere[J]. European Journal of Soil Biology, 2014, 61: 41–48.
[10] 劉桃菊, 戚昌瀚, 唐建軍. 水稻根系建成與產量及其構成關系的研究[J]. 中國農業(yè)科學, 2002, 35(11): 1416–1419.
[11] 趙利, 鈔建賓, 郭捷, 等. 基于代謝組學技術的植物抗病相關代謝物研究進展[J]. 西北植物學報, 2021, 41(6): 1071–1078.
[12] 趙培培, 王國棟. 植物活性特異性代謝物合成生物學應用研究進展[J]. 植物生理學報, 2020, 56(11): 2296–2307.
[13] 馬琳, 郜玉鋼, 臧埔, 等. 植物生長調節(jié)劑對藥用植物次生代謝物積累的影響[J]. 中南藥學, 2016, 14(8): 834–837.
[14] 李崇瑋, 柏新富, 陳國忠, 等. 不同恢復年限老參地土壤養(yǎng)分以及酚酸類代謝物含量差異[J]. 植物生態(tài)學報, 2021, 45(11): 1263–1274.
[15] Lu S, Song H X, Guan C Y, et al. Long-term rice-rice-rape rotation optimizes 1, 2-benzenediol concentration in rhizosphere soil and improves nitrogen-use efficiency and rice growth[J]. Plant and Soil, 2019, 445(1/2): 23–37.
[16] 康益晨, 楊昕宇, 張俊蓮, 等. 覆膜壟播及輪作蠶豆對馬鈴薯連作田根系分泌物及化感效應的影響[J]. 西北農業(yè)學報, 2020, 29(8): 1148–1158.
[17] 袁進成, 孟亞軒, 孫穎琦, 等. 基于全基因關聯(lián)分析的代謝組學在植物中的應用[J]. 中國農業(yè)科技導報, 2021, 23(9): 12–18.
[18] 劉美麗. 植物間相互作用及其林學意義分析[J]. 現(xiàn)代農業(yè)研究, 2018(6): 77–78.
[19] Li R, Liu J, Li J N, et al. Straw input can parallelly influence the bacterial and chemical characteristics of maize rhizosphere[J]. Environmental Pollutants and Bioavailability, 2020, 32(1): 1–11.
[20] 陳軍, 黃珊瑜, 劉冰, 等. 不同氮肥運籌對水稻根際土壤理化性質及代謝物質的影響[J]. 福建農業(yè)學報, 2015, 30(11): 1082–1089.
[21] Chen S, Liu S W, Zheng X, et al. Effect of various crop rotations on rice yield and nitrogen use efficiency in paddy-upland systems in southeastern China[J]. The Crop Journal, 2018, 6(6): 576–588.
[22] Cheng N, Peng Y J, Kong Y L, et al. Combined effects of biochar addition and nitrogen fertilizer reduction on the rhizosphere metabolomics of maize (L.) seedlings[J]. Plant and Soil, 2018, 433(1/2): 19–35.
[23] Kirkby C A, Richardson A E, Wade L J, et al. Nutrient availability limits carbon sequestration in arable soils[J]. Soil Biology and Biochemistry, 2014, 68: 402–409.
[24] 穆文強, 康慎敏, 李平蘭. 根際促生菌對植物的生長促進作用及機制研究進展[J]. 生命科學, 2022, 34(2): 118–127.
[25] Fang W G, St Leger R J. Mrt, a gene unique to fungi, encodes an oligosaccharide transporter and facilitates rhizosphere competency in[J]. Plant Physiology, 2010, 154(3): 1549–1557.
[26] Abdel-Lateif K, Bogusz D, Hocher V. The role of flavonoids in the establishment of plant roots endosymbioses with arbuscular mycorrhiza fungi, rhizobia andbacteria[J]. Plant Signaling & Behavior, 2012, 7(6): 636–641.
[27] Choi H, Jin J Y, Choi S, et al. An ABCG/WBC-type ABC transporter is essential for transport of sporopollenin precursors for exine formation in developing pollen[J]. The Plant Journal: for Cell and Molecular Biology, 2011, 65(2): 181–193.
[28] Singh H P, Kaur S, Batish D R, et al. Nitric oxide alleviates arsenic toxicity by reducing oxidative damage in the roots of(rice)[J]. Nitric Oxide, 2009, 20(4): 289–297.
[29] Xie Z Y, Wang J, Wang W S, et al. Integrated analysis of the transcriptome and metabolome revealed the molecular mechanisms underlying the enhanced salt tolerance of rice due to the application of exogenous melatonin[J]. Frontiers in Plant Science, 2021, 11: 618680.
[30] Yara A, Yaeno T, Montillet J L, et al. Enhancement of disease resistance toin rice by accumulation of hydroxy linoleic acid[J]. Biochemical and Biophysical Research Communications, 2008, 370(2): 344–347.
Effects of Rice-potato with Straw Mulch in Winter on Soil Metabolite Expression in Rice Rhizosphere at Full-heading Stage
WANG Mengjia1, FENG Xiangqian1, 2, MA Hengyu1, LIU Yuanhui1, CHU Guang1, XU Chunmei1, ZHANG Xiufu1, WANG Danying1, CHEN Song1*
(1 State Key Laboratory of Rice Biology, China National Rice Research Institute, Hangzhou 311400, China; 2 College of Agriculture, Yangtze University, Jingzhou, Hubei 434025, China)
Rice-potato with straw mulch in winter is an economical and high-yield paddy-upland rotation system in southern China, and its effects on soil metabolites in rhizosphere may be the key to achieve the rice yield potential but it is still unclear. Based on a long-term experiment of paddy-upland rotation (since 2003 Fuyang of Zhejiang), the rice-fallow (CK) was set as control and composition and possible function of soil rhizosphere metabolites of rice-potato (straw mulch and return, RP) were evaluated. The rhizosphere soil samples were collected at full heading stage using “root bags” and evaluated by Liquid Chromatography-Mass Spectrometry (LC-MS). The results show that: 1) By using the OPLS-DA model, 201 different expressed rhizosphere soil metabolites (VIP >1 and<0.05) are screened out under the RP rotation, as compared to that under CK, and these metabolites are attributed to 11 super-class, such as lipids and lipid-like molecules, organic acids and derivatives, organic oxygen compounds, etc. (based on HMDB database). In addition, the abundances of phenylpropanoids and polyketides, prenol lipids, benzene and substituted derivatives, steroid lactones, amino acids/peptides, carbohydrates and fatty acyl glycosides are changed significantly in response to the change of rotation from CK to RP, which indicate that these metabolites could be used as characteristics traits of rhizosphere soil metabolites of rice to distinguish the RP rotation from CK. 2) Organic oxygen compounds, organic acids and derivatives, phenylpropanoids and polyketides have significant positive correlation with the differences of soil organic matter, total potassium, available nitrogen and available potassium (=0.816–0.938), which not only provide the nutrient matter directly or indirectly, but also assist in promoting the rice growth. 3) Phenylpropanoids and polyketides, nucleosides/nucleotides and analogues, lipids and lipid-like molecules, which have significant positive correlation with rice grain yield (= 0.957–0.999), might improve the stress resistance of rice by participating in a series of metabolic pathways.
Rice rhizosphere; Metabolites; Rotation; Straw returning
S210.50
A
10.13758/j.cnki.tr.2023.02.009
王孟佳, 馮向前, 馬橫宇, 等. 水稻–冬季秸稈覆蓋種植馬鈴薯模式對水稻齊穗期根際土壤代謝組表達特征的影響. 土壤, 2023, 55(2): 295–304.
浙江省自然科學基金項目(LY22C130001)、浙江省重點研發(fā)計劃項目(2022C02008)和國家自然科學基金項目(32172106)資助。
(chensong02@caas.cn)
王孟佳(1997—),女,河南盧氏人,碩士研究生,主要從事稻田土壤根際活性有機碳組分研究。E-mail:wmj15837605820@163.com
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
音樂探索(2022年2期)2022-05-30 21:01:37
今日農業(yè)(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(yè)(2021年14期)2021-10-14 08:35:40
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
小天使·一年級語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
