梅毒螺旋體比較基因組學研究進展
2023-06-06 01:51:30趙思思趙飛駿
中南醫學科學雜志 2023年3期
趙思思,趙飛駿
南華大學衡陽醫學院病原生物學研究所,湖南衡陽 421001
梅毒螺旋體(Treponema pallidum,Tp)是蒼白密螺旋體屬中的蒼白亞種,其引起的梅毒是一種嚴重危害人類健康的性傳染性疾病。梅毒發病率不斷升高,中國每年報告的梅毒診斷數呈逐年遞增趨勢[1-2]。隨著基因測序技術日趨完善,基因組學得到了巨大的發展。比較基因組學通過比較分析Tp與其他致病性螺旋體的基因組數據,揭示Tp與其他致病性螺旋體的相似性與差異性,以幫助了解Tp的生物特性和致病機制等。因此,本文綜述Tp基因組、Tp和其他致病性螺旋體基因組比較,以探討Tp的致病機制。
1 Tp基因組
Tp基因組為環狀染色體,有最小的螺旋體基因組,大小約為1.14 Mb,G+C的平均含量為52.8%[3]。預測Tp蛋白質分子量為3 235~72 869 Da,等電點為3.9~12.3[3]。Nichols株是第一個完成全基因組測序的Tp菌株,目前已相繼完成了標準株SS14株、Chicago株、DAL-1株、Mexico A株、Sea81-4株的全基因組測序[3-8]。Tp菌株間基因序列相似性水平>99.8%[9]。
1.1 Nichols株
1912年,Nichols株從華盛頓特區的一名神經梅毒患者的腦脊液中被分離出來,并于1998年3月完成了首次測序工作[3]。2012年,科學家使用新的測序方法對Nichols株進行了第二次測序[4]。兩種測序方法得到的數據有些微差異[4]。NCBI最新數據顯示Nichols株基因組大小為1 138 011 bp,有1 010個蛋白質編碼區(coding sequence,CDS)(https://www.ncbi.nlm.nih.gov/nuccore/NC_000919.1)。根據Riley的分類方案,Nichols株中有577個承擔預測生物學功能的開放閱讀框(open reading frame,ORF),177個可與其他物種假定蛋白質相匹配的ORF,還有287個沒有數據庫匹配的ORF[3]。Nichols株中可以發生tprK變異,但其變異速度較低[10]。
1.2 SS14株
SS14株從亞特蘭大的一名二期梅毒患者身上被分離出來,該患者對青霉素過敏且對紅霉素治療無效[4]。SS14株對紅霉素表現出高水平耐藥性,這是人們第一次在致病性密螺旋體中發現與臨床相關的抗生素耐藥性[11]。2007年8月,SS14株進行了首次全基因組測序工作,2012年12月,研究者使用新的測序技術更新了基因組測序結果。NCBI最新數據顯示SS14株基因組大小為1 139 457 bp,有1 014個CDS(https://www.ncbi.nlm.nih.gov/nuccore/NC_021508.1)。SS14株的基因組測序結果首次揭示了Tp的全基因組變異性[12]。
Matejkova等[12]發現,SS14株與Nichols株相比有327個單核苷酸變化、14個缺失和18個插入,這其中不包括高度可變的tprK基因變化[12]。使用新的測序方法校正Nichols株和SS14株的全基因組數據后,這兩種菌株的蛋白質組發生了顯著變化,提示Nichols株和SS14株代表了不同的Tp菌株亞群[4]。SS14株和Nichols株基因組之間的重要差異主要存在于23S rRNA、tp0326、tp0868和tp0127基因中[5]。在SS14株的23S rRNA基因中的A2058G點突變,是SS14株對大環內酯類藥物耐藥的原因[11]。體外測試表明,SS14株對各種抗生素的敏感性均低于Nichols株[4]。此外,SS14株基因組中的tprK基因變異性比Nichols株基因組中的tprK變異性高得多,SS14株基因組鑒定的菌株內異質性也顯著高于Nichols株[4]。SS14株和Nichols株之間的多樣性比SS14株和Mexico A株基因組之間的多樣性大很多[4]。
1.3 Chicago株
1951年,Chicago株從梅毒患者的原發性硬下疳中被分離出來,其可以在兔睪丸內生長良好[6]。2009年9月,研究者成功完成了對Chicago株基因組的測序。NCBI最新數據顯示Chicago株基因組大小為1 139 281 bp,編碼989個CDS(https://www.ncbi.nlm.nih.gov/nuccore/NC_017268.1)。與Nichols株基因組相比,Chicago株有44個核苷酸替換、21個缺失和75個插入[6]。Chicago株tprK序列具有高度多樣性,而Nichols株的tprK基因變異較慢[10]。
1.4 DAL-1株
1991年,DAL-1株從達拉斯的一名二期梅毒孕婦的羊水中被分離出來;2011年11月,美國研究者組合使用454測序、Solexa測序和Sanger測序確定完整DAL-1基因組序列[5]。NCBI數據表明其基因組大小為1 139 971 bp,包含986個CDS(https://www.ncbi.nlm.nih.gov/nuccore/NC_016844.1)。
1.5 Mexico A株
1953年,Mexico A株從居住在墨西哥的一名患有原發性梅毒的18歲男性患者體內被分離出來[7]。NCBI最新數據顯示,Mexico A株基因組大小為1 140 038 bp,編碼986個CDS(https://www.ncbi.nlm.nih.gov/nuccore/NC_018722.1)。與Nichols株相比,體外培養的Mexico A株生長速率和運動性降低[13]。Mexico A株在目前研究的所有Tp菌株中擁有最大的基因組,其基因組有1 140 038個堿基。Mexico A株的兩個基因tp0326和tp0488結合了Tp和蒼白密螺旋體極細亞種的特異性核苷酸序列,這可能是因為單個宿主同時感染兩種菌株時菌株間進行了基因重組,這證明密螺旋體亞種之間可以進行水平基因轉移[7]。對Mexico A株的G+C含量、密碼子和氨基酸使用以及基因位置進行分析,預測其有77個基因能水平轉移[7]。Mexico A株基因組與SS14株基因組親緣關系最近。Mexico A基因組與SS14基因組相比只有175個替換、85個插入和28個缺失;與Chicago株基因組相比有419個替換、18個插入和20個缺失,而與Nichols基因組相比有438個替換、94個插入和38個缺失[7]。
1.6 Sea81-4株
Sea81-4株于1980年從西雅圖的一名患者的原發性硬下疳中被分離出來,在2012年完成了測序[8]。NCBI最新數據顯示,Sea81-4株基因組大小為1 139 203 bp,有1 000個CDS(https://www.ncbi.nlm.nih.gov/nuccore/NZ_CP003679.1)。與其他5種Tp分離株不同,Sea81-4株靜脈接種感染兔模型后會導致中樞神經系統持續感染[8,14]。
編碼假定毒力因子的基因、參與細胞結構和過程的基因和編碼DNA復制、修復和重組的基因,是Tp不同菌株間基因差異的主要發生位點。相比之下,編碼一般代謝、轉錄、翻譯、基因調控和轉運成分的Tp基因是保守的[7]。Tp菌株之間的遺傳差異首先在tprD基因和tp0126~tp0127基因間的區域中被發現[5]。tprK基因在Tp各菌株之間具有高度變異,這種變異使得Tp可以逃脫免疫清除,導致宿主的慢性感染[15]。來自不同實驗室的Nichols株的tprK基因在經過幾十年的獨立傳代后仍只有很小的序列多樣性[10]。但除Nichols株外,其他菌株均已鑒定出基因序列多樣性高的tprK基因[10]。tprK序列的多樣性位于7個可變區域(V1~V7)和保守區域的兩側[16]。在Tp感染期間,T細胞免疫反應針對tprK的保守區,而抗體反應針對V區[17]。tprK的多樣性隨著Tp在兔睪丸中的連續傳代而積累,其中V6是V區中變化最大的[16]。
2 Tp和其他螺旋體基因組比較
2.1 Tp和其他蒼白密螺旋體基因組比較
近年來全基因組測序結果表明,密螺旋體是高度克隆的生物,其基因組的細微差異可導致疾病的臨床表現和宿主范圍的深層次差異[5]。對人致病的密螺旋體有蒼白密螺旋體和品他密螺旋體。蒼白密螺旋體又分為3個亞種,分別為蒼白亞種、極細亞種和地方亞種。除此之外,還有一種未分類的類人猿密螺旋體分離株(Fribourg-Blanc)。梅毒螺旋體、雅司螺旋體和Fribourg-Blanc螺旋體關系密切,這三者無法通過形態學、蛋白質電泳、細菌生理學或宿主免疫反應來區分[18]。
2.1.1 Tp和雅司螺旋體基因組比較 雅司螺旋體是雅司病的病原體。雅司病是一種熱帶病,主要以皮膚、關節、軟組織和骨骼影響為特征。一般來說,雅司螺旋體的毒性比Tp小[9]。盡管Tp和雅司螺旋體會引起完全不同的疾病,但這兩個亞種的基因組總差異僅為0.36%~0.37%;提示Tp和雅司螺旋體之間存在極大的序列相似性[5,18]。使用下一代測序技術確定了3種雅司螺旋體菌株的完整基因組序列,其基因組長度為1 139 330~1 139 744 bp[9]。
Tp和雅司螺旋體之間的大部分遺傳差異主要累積在6個基因組區域中,包括第122位的tp0138(tpF1)、16S rRNA、tpp15(tp0171)的5′側翼區域和3′側翼區域、第579位的gpd(Tp0257)、tp0326(Tp92)以及tprI和tprC,這些基因位點的差異可能導致了Tp和雅司螺旋體之間致病性的差異[9]。在tpp15的5′側翼區域和3′側翼區域中發現的序列變化可以將Tp與其他密螺旋體區分開來,包括雅司螺旋體、地方密螺旋體和Fribourg-Blanc密螺旋體[18]。
2.1.2 Tp和Fribourg-Blanc株基因組比較 Fribourg-Blanc株是1962年從非洲幾內亞的一只狒狒中分離出來,能夠感染倉鼠及人類[18]。Fribourg-Blanc株在所有不可培養的密螺旋體中有最大的基因組(1 140.4 kb)[9]。Fribourg-Blanc分離株在tp0696~tp0697之間的基因間區域中鑒定出了DNA重復,因此Fribourg-Blanc分離株不太可能包含Tp菌株中缺失的任何獨特DNA區域[9]。在tprK供體位點所在的基因組區域中,蒼白密螺旋體和Fribourg-Blanc分離株之間存在高程度的相似性和序列保守性(99.57%),其進化關系非常密切[9]。此外,Fribourg-Blanc分離株與雅司螺旋體的關系比與Tp的關系更密切[9,19]。
2.2 Tp和其他致病性螺旋體基因組比較
通過比較Tp與其他不同屬的致病性螺旋體間的基因差異(表1),可以進一步揭示Tp的生物學特性。
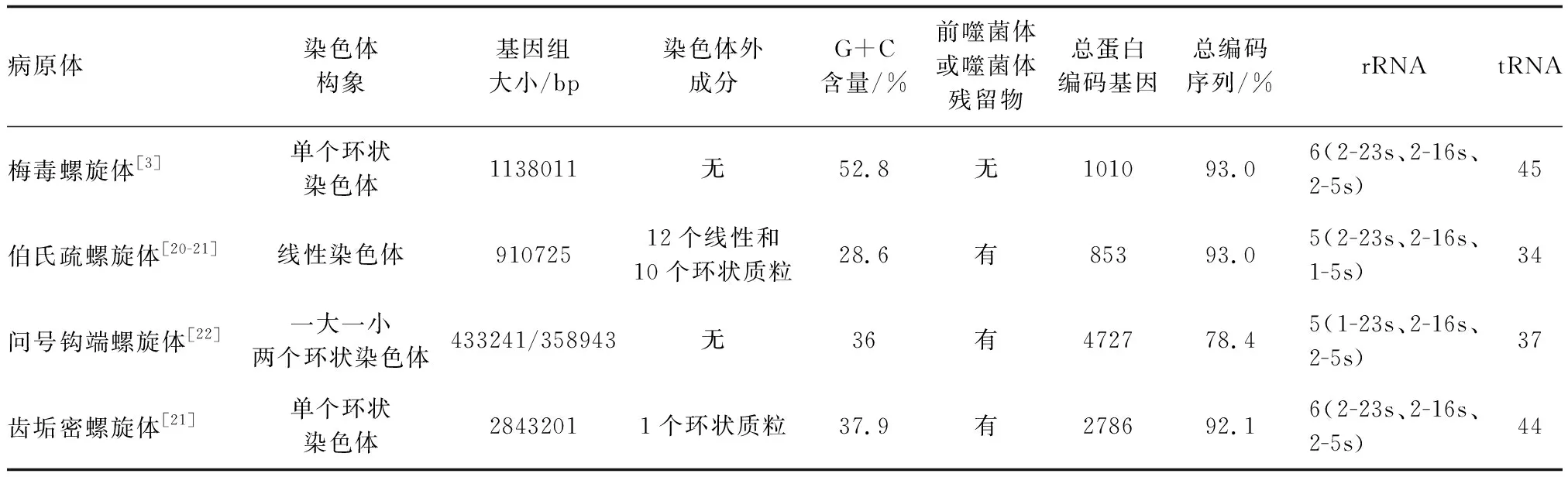
表1 螺旋體基因組的特征
2.2.1 Tp與問號鉤端螺旋體基因組比較 問號鉤端螺旋體的基因組由兩條環狀染色體組成,其基因組大小遠大于Tp和Bb[22]。問號鉤端螺旋體共有4 727個CDS,4 360個位于CⅠ上,367個位于CⅡ上,所有rRNA和tRNA基因都位于CⅠ上。問號鉤端螺旋體、Tp和Bb有315個同源基因[22]。與Tp不同的是,問號鉤端螺旋體缺乏編碼己糖激酶的基因,但其有Tp中不存在的編碼完整長鏈脂肪酸利用系統、三羧酸循環(TCA)和呼吸電子傳遞鏈的基因[22]。因此,Tp的腺苷三磷酸(adenosine triphosphate,ATP)產生方式與問號鉤端螺旋體不同,Tp通過糖酵解途徑生成ATP,而問號鉤端螺旋體通過氧化磷酸化產生ATP[22]。除此之外,問號鉤端螺旋體通過編碼完整的氨基酸和核苷酸生物合成代謝系統來獲得營養[23],而Tp編碼廣泛的轉運體從宿主獲取營養[24]。與Tp一樣的是,問號鉤端螺旋體使用FlaA鞘蛋白和FlaB核心蛋白作為其內鞭毛細絲的基本成分[22]。
2.2.2 Tp與伯氏疏螺旋體基因組比較 Bb最顯著的特征之一是有一個不尋常的基因組,其中包含一個線性染色體和許多線性/環狀質粒[25]。Bb的線性染色體包含了853個基因,編碼一組用于DNA復制、轉錄、翻譯、物質轉運和能量代謝的基本蛋白質[20]。同時,Bb的11個質粒上的30個基因中,大多數沒有已知的生物學功能[20]。
Tp和Bb之間差異有顯著性,兩個病原體基因組的整體直系同源系數值約為0.43[26]。參與核心生物學功能的基因同源性很高,而參與特定過程的基因存在顯著的變異性[26]。Tp和Bb有476個ORF同源,其中有76%具有預測的生物學功能,24%編碼功能未知蛋白質[3]。編碼功能未知蛋白的ORF中有近50%是螺旋體獨有的,這組螺旋體屬獨有的蛋白質可能決定了螺旋體結構和生理的特殊性,可能與Tp和Bb感染人類并引起慢性傳播性疾病的能力有關。此外,Tp中有90個功能未知的ORF與Bb染色體上的編碼基因相匹配,沒有ORF與Bb質粒上的編碼基因匹配,表明質粒是Bb所特有的[3,20]。Tp的第三密碼子位置有G或C偏向,Bb在該位置有A或T偏向,這使得Tp基因組中的G+C含量幾乎是Bb基因組中的兩倍[3]。Tp與Bb基因組之間不同的G+C含量會造成總體密碼子使用的偏差,是預測編碼序列中的氨基酸組成不同[20]。
2.2.3 Tp和齒垢密螺旋體基因組比較 齒垢密螺旋體是一種與牙周病相關的口腔螺旋體。齒垢密螺旋體的基因組構象與Tp一致,但其基因組大小比Tp大得多[21]。齒垢密螺旋體預計編碼2 786個CDS,其中734個CDS是獨特的[21]。盡管Tp和齒垢密螺旋體基因組大小顯著不同,但這兩種病原體間存在的穩定RNA的數量幾乎相同[21]。有觀點認為,1.14 Mb的Tp基因組是由齒垢螺旋體缺失和(或)分化而來[27]。Tp基因組與齒垢密螺旋體基因組可以共享有限的核苷酸相似性,大約25%的齒垢密螺旋體基因與Tp基因組中68%的CDS最匹配[27]。除了編碼核糖體和鞭毛蛋白的高度保守操縱子外,Tp基因組與齒垢密螺旋體基因組之間基本上不存在同線性[27]。與齒垢密螺旋體相比,Tp沒有公認的限制修飾系統、插入序列元件或噬菌體[21]。Tp基因組編碼一個有12個成員家族(TprA-L)的假定膜蛋白,這個家族蛋白具有免疫保護性和調理活性,編碼這些蛋白的基因家族在不同亞種和菌株之間表現出異質性[28]。齒垢密螺旋體中也擁有一個與該基因家族相關的成員,即TDE0405,其編碼齒垢密螺旋體的主要外鞘蛋白[28]。同時,Tp和齒垢密螺旋體都具有磷酸轉移酶系統(phosphotransferase system,PTS)的HPr、酶Ⅰ和酶ⅡA,但沒有PTS轉運蛋白復合物,表明這些蛋白質在Tp和齒垢密螺旋體體內只起著調節作用[21]。齒垢密螺旋體中存在糖酵解和TCA循環,表明其產生ATP的方式與Tp相似都是由糖發酵產生。但與Tp不同的是,齒垢密螺旋體可以從頭合成脂肪酸、輔因子和核苷酸[21]。
3 展 望
比較基因組學應用于Tp,使Tp的生物特性、進化方向以及致病機制等方面更進一步得到了解。目前已經有許多Tp標準株和臨床株的全基因組測序工作已經完成。通過Tp菌株間的比較,Tp的主要突變區域和保守區域被明確,這為梅毒疫苗的開發奠定了堅實的基礎。比較Tp基因組與其他致病性螺旋體基因組的差異為研究Tp的致病機制提供了新的方向。