OTUD3敲除對神經干細胞中縫隙連接蛋白43表達的影響
2023-06-08 09:51:08徐同迎焦倩姜宏
精準醫學雜志 2023年2期
關鍵詞:小鼠
徐同迎 焦倩 姜宏
(青島大學基礎醫學院,山東省神經相關疾病的機制與重點防治實驗室,山東省沿海地區神經退變疾病協同創新中心,山東 青島 266071)
神經干細胞(NSCs)存在于哺乳動物胚胎的腦組織以及成年人側腦室室管膜下區和海馬齒狀回顆粒下層中,具有自我增殖和分化的潛能[1-3]。中樞神經系統受到損傷以后,靜止的NSCs被激活并參與到中樞神經系統的損傷修復過程中[4]。去泛素化酶(DUBs)可以逆轉蛋白質泛素化降解過程,進而影響多種生物學過程,包括細胞凋亡和自噬、調節細胞周期和干細胞的發育等[5-8]。卵巢腫瘤結構域蛋白(OTUs)是DUBs家族成員,具有內源性連鎖特異性[9-10]。OTUD3是OTUs亞家族中的一員,與腫瘤的發生發展和神經退行性疾病關系密切,其底物包括PTEN、GRP78、p53和IRP2蛋白[8,11-14]。縫隙連接蛋白(Cx)通道是機體大多數組織中相鄰細胞間及細胞與細胞外區域進行物質交換和信號交流主要通道[15-16]。其中Cx43是Cx中分布最廣泛家族成員[17],與基因轉錄、發育、自噬調節、細胞內運輸和細胞外小泡介導遠程通訊有關[18-20]。OTUD3是否調節Cx43而參與NSCs增殖分化等生物學行為,目前尚未見相關報道。本研究利用OTUD3敲除(OTUD3-/-)的胚胎小鼠,于體外分離并且培養OTUD3-/-和野生型(WT)胚胎小鼠NSCs,檢測OTUD3對NSCs中Cx43表達影響,同時提取胚胎小鼠腦皮質蛋白,驗證OTUD3對Cx43表達影響。
1 材料和方法
1.1 實驗試劑
表皮生長因子(EGF)、堿性成纖維細胞生長因子(bFGF)、N-2 supplement、B-27 supplement購自美國Gibco公司;GAPDH抗體和Cx43抗體購自美國CST公司;OTUD3抗體購自美國Abcam公司;Ki67抗體、Nestin抗體、β-tubulin Ⅲ抗體購自美國Millipore公司;羊抗兔IgG-HRP購自中國愛博泰克生物公司;ECL發光液購自美國Millipore公司。
1.2 實驗動物及其飼養
3月齡OTUD3-/-小鼠(中國軍事醫學科學院提供)和3月齡WT小鼠(北京維通利華實驗動物技術有限公司提供)置于恒溫(21±2)℃、恒濕(50±10)%、12-12 h晝夜交替光照下飼養,自由飲水與取食。同種基因型小鼠之間進行交配,待妊娠第14天時,異氟烷氣體麻醉孕鼠,以體積分數0.75的乙醇消毒腹部后,沿腹中線剪開,使用鑷子取出串珠樣OTUD3-/-小鼠和WT胚胎小鼠各3只,置于預冷的PBS中,用于后續實驗。
1.3 實驗方法
1.3.1胚胎小鼠腦皮質組織的提取 剝離出所有OTUD3-/-和WT胚胎小鼠的全部腦組織,剔除腦膜后分離腦皮質組織。取出每只胚胎小鼠的一部分腦皮質組織置入1.5 mL EP管內,加入配置好的蛋白裂解液,組織研磨儀研磨1 min,冰上裂解30 min后,4 ℃下12 000 r/min離心20 min,提取腦皮質組織蛋白用于后續實驗。
1.3.2NSCs的分離和傳代 將OTUD3-/-和WT胚胎小鼠剩余部分腦皮質組織,剪碎至1 mm×1 mm×1 mm,使用1 mL槍頭反復吹打組織直至形成細胞懸液,400目篩網過濾后置于大離心管中,以1 000 r/min離心5 min,棄去上清液;加入完全培養液重懸細胞,細胞計數后將其接種于75 cm2培養瓶中,接種密度約為1×108個/L,并分別標記為OTUD3-/--NSCs和WT-NSCs,置于37 ℃、含體積分數0.05的CO2細胞培養箱中進行培養;隔天添加完全培養基,培養4~6 d時,根據細胞狀態(細胞生長所形成神經球聚集增多時)傳代,傳至第3代時,于倒置顯微鏡下觀察第1、3天時OTUD3-/--NSCs和WT-NSCs的生長狀態。
1.3.3NSCs的培養 將高壓滅菌后的玻片置于24孔板中,加入多聚賴氨酸溶液室溫下孵育5 min,0.01 mol/L PBS洗2次后,超凈臺晾干;將傳代至第3代的神經球消化為單細胞后,調整細胞密度為1×109個/L,接種細胞懸液至上面制備好的玻片上,置于37 ℃、含體積分數為0.05的CO2細胞培養箱中培養。取OTUD3-/--NSCs和WT-NSCs各3個玻片,使用完全培養基貼壁培養3 d以后,用于檢測Ki67;取OTUD3-/--NSCs和WT-NSCs各6個玻片,使用分化培養基貼壁培養7 d以后,用于檢測Nestin和β-tubulin Ⅲ。
1.3.4細胞免疫熒光染色 將前面培養3 d或7 d的OTUD3-/--NSCs和WT-NSCs玻片從24孔板取出,吸棄培養基,用4 ℃預冷的4% PFA溶液室溫固定30 min,0.01 mol/L的PBS清洗3次,每次5 min,加入封閉液,室溫下封閉1 h;向使用完全培養基培養3 d的NSCs中加入Ki67一抗(1∶400),向使用分化培養基培養7 d的NSCs中加入Nestin一抗(1∶100)和β-tubulin Ⅲ一抗(1∶500),室溫孵育1 h后,4 ℃搖床孵育過夜,0.01 mol/L PBS洗3次,每次5 min;加入AF555或AF488標記的熒光二抗(1∶500),室溫避光孵育1 h;將含DAPI封片液滴于載玻片上,將生長細胞的玻片面緩慢接觸封片液,避光保存,倒置熒光顯微鏡下觀察并拍照。
1.3.5Western blotting方法檢測腦皮質和NSCs中OTUD3和Cx43表達水平 NSCs培養至第3代時,傳代后接種于6孔板(2 mL/孔)中,每組設置3個復孔,調整密度為1×108個細胞/L;繼續培養至細胞融合度約90%時,加入已配置好的蛋白裂解液,組織研磨儀研磨1 min,冰上裂解30 min后,4 ℃下12 000 r/min離心20 min,提取NSCs蛋白用于后續實驗。配制聚丙酰胺凝膠,向凝膠孔加入1.3.1中制備好的腦皮質組織蛋白樣品或NSCs蛋白樣品,電壓調至80 V進行SDS蛋白電泳,待樣品進入分離膠后,將電壓調為120 V,PVDF轉膜,使用100 g/L脫脂奶粉室溫封閉2 h以后,分別加入OTUD3一抗(1∶1 000)和Cx43一抗(1∶1 000),4 ℃搖床孵育過夜,次日使用TBST洗條帶3次,每次10 min;根據一抗種屬加入HRP偶聯的山羊抗兔二抗(1∶10 000),于室溫下孵育1 h后,TBST漂洗3次,每次洗10 min,ECL化學發光液避光孵育1 min顯影。使用Image J軟件分析蛋白條帶灰度值,以GAPDH條帶結果作為內參照,計算目的蛋白的相對表達量。
2 結 果
2.1 WT和OTUD3-/-胚胎小鼠NSCs的鑒定
倒置顯微鏡下觀察顯示,第3代OTUD3-/--NSCs和WT-NSCs培養1 d時均形成由多細胞組成的神經球,培養3 d時增殖形成的神經球均增多(圖1)。細胞免疫熒光染色檢測結果顯示,使用完全培養基培養3 d時的WT-NSCs和OTUD3-/--NSCs當中Ki67均染色陽性(圖2A),圖中綠色為Ki67;使用分化培養基培養至第7天WT-NSCs和OTUD3-/--NSCs中Nestin和β-tubulin Ⅲ均染色陽性(圖2B),其中綠色為Nestin,紅色為β-tubulin Ⅲ。通過以上結果可以鑒定本研究中原代提取培養的細胞為NSCs,具有增殖和分化能力。
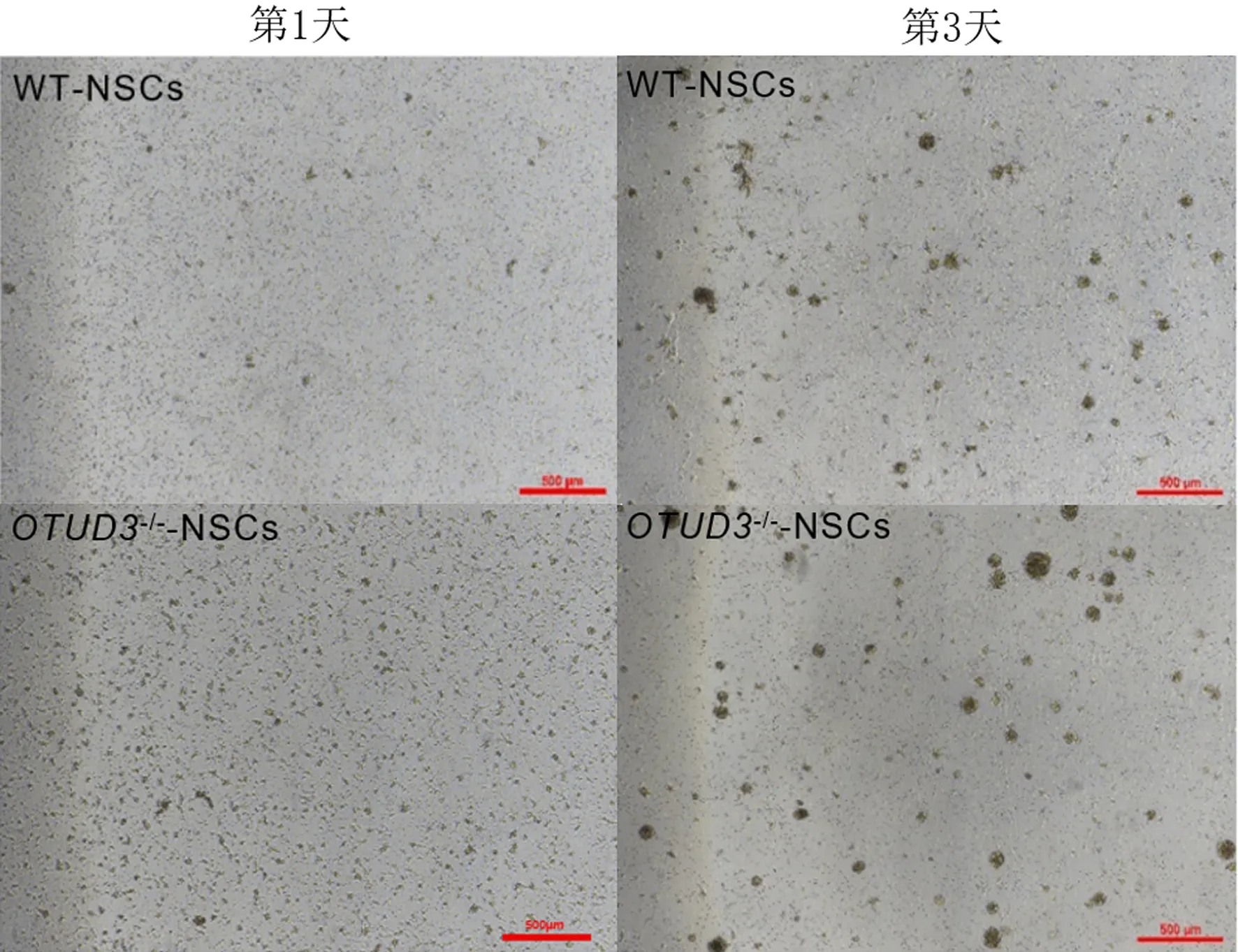
圖1 第3代WT-NSCs和OTUD3-/--NSCs培養第1、3天時神經球生長狀況(4倍)
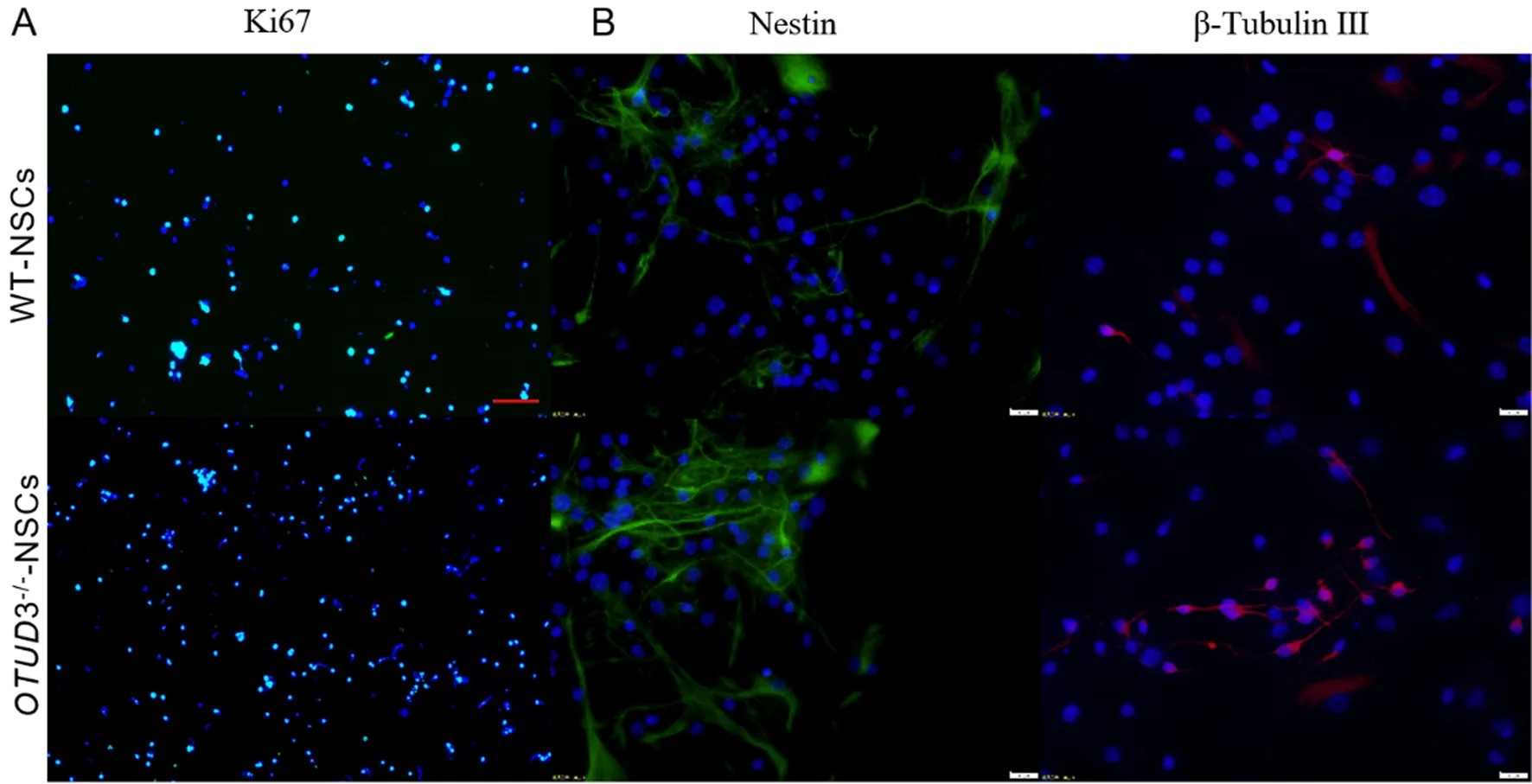
A:第3代WT-NSCs和OTUD3-/--NSCs培養第3天時Ki67染色呈陽性(細胞免疫熒光染色,10倍),B:第3代WT-NSCs和OTUD3-/--NSCs培養第7天時Nestin和β-Tubulin Ⅲ染色陽性(細胞免疫熒光染色,40倍)
2.2 WT和OTUD3-/-胚胎小鼠NSCs和腦皮質組織中OTUD3、Cx43的表達情況
Western blotting法檢測結果示,WT-NSCs當中OTUD3、Cx43條帶灰度值分別為1.023±0.062、1.035±0.093;OTUD3-/--NSCs中無OTUD3蛋白表達,Cx43條帶的灰度值為0.592±0.102。WT-NSCs和OTUD3-/--NSCs的上述兩種蛋白比較,差異均有顯著意義(t=16.82、3.21,P<0.05),見圖3A。WT胚胎小鼠腦皮質組織中OTUD3、Cx43條帶的灰度值分別為0.929±0.060、0.905±0.0149;OTUD3-/-胚胎小鼠腦皮質組織中無OTUD3蛋白表達,Cx43條帶灰度值為0.522±0.012,WT胚胎小鼠和OTUD3-/-胚胎小鼠腦皮質組織上述兩種蛋白比較,差異均具有顯著意義(t=10.04、20.41,P<0.05),見圖3B。
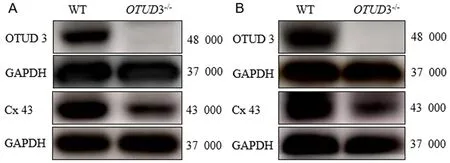
A:WT-NSCs和OTUD3-/--NSCs中OTUD3、Cx43表達,B:WT和OTUD3-/-胚胎小鼠腦皮質中OTUD3、Cx43表達
3 討 論
NSCs是一類具有自我增殖和分化潛能的細胞,在神經系統發育和修復受損神經組織中發揮重要作用,具有修復和代替受損腦組織的功能,并可以重建部分神經功能環路[3,21-22]。近年研究發現神經系統疾病的發生發展與縫隙連接密切相關,縫隙連接在細胞間信息傳遞、物質交換和維持內環境穩態方面都發揮重要作用[23]。Cx43在神經元與星形膠質細胞、星形膠質細胞與少突膠質細胞間可以傳遞死亡信號以及鈣超載等危險信號[24]。也有研究表明Cx43可以通過氧化應激和內質網應激調控腎小管上皮細胞損傷,抑制Cx43后可以促進細胞生長并提升細胞對于應激情況的抵抗能力[25],但目前具體機制尚不清楚。NSCs存在于哺乳動物胚胎期的大腦,胚胎小鼠大腦皮質中有大量的NSCs,Cx43在胚胎發育期的腦組織中廣泛表達,本研究選擇胚胎期第14天小鼠的腦皮質區,體外培養NSCs,探討Cx43調控NSCs生物行為學的具體機制。
泛素化信號通路在神經發育和神經元生理功能中發揮重要的調控作用,而DUBs作為調節蛋白泛素化的中心分子,其對神經系統的調控受到越來越多的關注。OTUD3屬于DUBs家族,是OTUs亞家族中的一員[11],其功能具有組織特異性和時間特異性,可通過調控不同靶蛋白的穩定性,調控細胞周期和細胞增殖[26]。OTUD3可以穩定PTEN蛋白表達水平,PTEN的缺乏可以調控細胞周期相關基因cyclinB2和cyclinD1,進而縮短細胞周期,影響NSCs增殖分化[11]。本研究中使用異氟烷氣體麻醉孕鼠以后取出串珠樣胚胎小鼠,分離提取胚胎WT和OTUD3-/-小鼠NSCs,分別標記為WT-NSCs和OTUD3-/--NSCs,在完全培養基中進行培養,倒置顯微鏡下觀察其生長狀態良好。Ki67是一種表達于細胞周期G1、S、G2和M期的核抗原,其表達水平能夠客觀地反映細胞的增殖狀態。本研究應用細胞免疫熒光染色對第3代培養了3 d的WT-NSCs和OTUD3-/--NSCs進行Ki67檢測,結果為陽性;應用細胞免疫熒光染色對第3代培養了7 d的WT-NSCs和OTUD3-/--NSCs進行Nestin和β-tubulin Ⅲ檢測,結果為陽性,提示本研究中原代提取培養的細胞為NSCs,并有增殖和分化能力。
接下來,本研究應用Western blotting技術檢測敲除OTUD3后胚胎小鼠NSCs和腦皮質組織中OTUD3蛋白的表達情況,結果顯示,與WT-NSCs相比,OTUD3-/--NSCs無OTUD3蛋白的表達;與WT胚胎小鼠腦皮質組織相比,OTUD3-/-胚胎小鼠腦皮質組織也無OTUD3蛋白的表達,提示胚胎小鼠NSCs中和腦皮質組織中OTUD3敲除成功。
Cx43參與細胞增殖過程,通過控制cyclinD1-CDK4-p27復合體的核轉位,進而阻斷細胞周期進程[27]。NSCs發育的同時,Cx亞型的表達模式也會發生變化,Cx43在胚胎發育期的腦組織中廣泛表達[28]。隨著NSCs在體內分化為神經元,Cx43表達水平下降,但是Cx43在NSCs表達調控仍需進一步研究。為明確Cx43在OTUD3調控NSCs生物學功能中的作用,本研究采用Western blotting技術檢測敲除OTUD3后胚胎小鼠腦皮質和NSCs中Cx43的表達情況,結果顯示OTUD3-/--NSCs中Cx43表達水平明顯降低,同時在動物水平進一步驗證,顯示OTUD3-/-后Cx43表達明顯降低,由于Cx43是NSCs增殖的抑制蛋白,通過改變細胞周期進展參與調控NSCs的增殖行為。而OTUD3可以通過調控下游靶蛋白,調控細胞增殖和分化。由此推測OTUD3可能通過調控Cx43影響NSCs的增殖和分化等生物學行為,進而影響神經發生。
綜上所述,敲除OTUD3后體外培養的NSCs中Cx43表達下降,可能通過影響細胞間的連接,進而促進NSCs的增殖,在神經發育過程發揮著重要作用。但OTUD3是否通過影響其泛素化水平影響Cx43表達,具體調節機制仍需進一步探討。
倫理批準和動物權利聲明:本研究涉及的所有動物實驗均已通過青島大學醫學部倫理委員會的審核批準(文件號QDU-AEC-20233-47)。所有實驗過程均遵照《倫理委員會標準/守則》的條例進行。
作者聲明:所有作者均參與了研究的設計、論文的寫作和修改。所有作者均閱讀并同意發表該論文,且均聲明不存在利益沖突。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
