不同含油量花生品種中二酰甘油酰基轉移酶基因的表達分析
2023-07-17 07:36:20潘麗娟許靜王秀貞姜驍陳娜王通殷祥貞楊偉強遲曉元
山東農業科學 2023年6期
關鍵詞:油脂
潘麗娟,許靜,王秀貞,姜驍,陳娜,王通,殷祥貞,楊偉強,遲曉元
(山東省花生研究所,山東青島 266100)
在油料作物中,油脂主要以三酰甘油(triacylgycerol,TAG)的形式貯存在種子中。 二酰甘油酰基轉移酶(diacylglycerol acyltransferase,DGAT)是催化二酰甘油合成TAG 的關鍵限速酶[1,2]。 目前報道的DGAT 存在四種類型——DGAT1、DGAT2、DGAT3 和DGAT/WS,彼此之間基因序列同源性很低,具有不同的亞細胞定位和表達模式[3-5],但都具有催化在二酰甘油加上酰基脂肪酸生成甘油三酯的作用[6-8]。
過表達DGAT1基因可促進種子中油脂的積累。 擬南芥DGAT1在煙草中過表達可使轉基因煙草種子中TAG 含量顯著上升[9]。 擬南芥突變體AS11的種子中DGAT1 活性降低,TAG 含量降低,脂肪酸組成發生變化,種子皺縮,發育遲緩,平均重量降低[10];而在野生型擬南芥中過量表達AtDGAT1后,種子含油量和千粒重得到明顯提高[11]。 過量表達高油玉米DGAT1的轉基因玉米種子含油量和油酸含量分別提高26.1% 和84.5%[12]。DGAT2在油料作物中過量表達,也能調控種子油脂的含量和組成[13,14],如蓖麻DGAT2過表達可促進蓖麻油中蓖麻油酸含量的增加[15]。DGAT3 和DGAT/WS 發現較晚,目前研究僅限于少數植物或微生物中,DGAT3 在TAG 合成中的功能已經在花生和產油酵母(oleaginous yeast)中被驗證[16]。
在花生中,Saha 等(2006)從未成熟種子的子葉中首先克隆到AhDGAT3基因[8]; 王龍龍(2010)從花生中克隆獲得了AhDGAT2的全長序列,在葉和花中表達量都較高[17];Chi 等(2014)通過構建的花生種子全長cDNA 文庫,首次克隆獲得AhDGAT1基因[18]。 目前花生中發現的這三種酶,AhDGAT1 和AhDGAT2 均有跨膜結構,屬于膜蛋白,而AhDGAT3 為可溶性蛋白;基因序列同源性很低;AhDGAT1和AhDGAT2在種子發育前中期表達量較高,而AhDGAT3則主要在種子發育后期表達[18]。 說明它們在種子生長發育、儲藏物質積累等方面所起的作用并不相同,可能在特定組織或發育階段由主效酶發揮作用[7,17]。
花生是我國重要的油料作物,近年來我國花生總產中約52%用于榨油,年產花生油近300 萬噸,占國產植物油產量的25%以上,是國產植物油的第二大來源[19]。 目前提高花生品種的含油量及改善其脂肪酸組成已成為育種專家努力的方向。 隨著分子生物學和基因工程技術的發展,對花生油脂合成代謝途徑及關鍵酶基因的認識也在不斷深入。 DGAT 作為Kennedy 途徑中唯一的限速酶[4],深入研究其在TAG 合成和積累過程中的作用對于闡明花生油脂及其脂肪酸組成的形成機理具有重要意義。 本研究分析了前期從花生中分離的3 個DGAT 基因AhDGAT1-1、AhDGAT1-2和AhDGAT3-3在含油量和油酸含量不同的3 個花生品種中的表達情況,以期為深入探究花生油脂合成和培育高油、高油酸新品種提供理論依據。
1 材料與方法
1.1 試驗材料
試驗所用3 個花生栽培品種分別為低油低油酸品種花育17 號、高油高油酸品種花育910 和高油低油酸品種花育918(表1),均為本課題組培育。 2020 年5 月初種植于山東省花生研究所萊西試驗站大田,田間水肥管理等同于常規大田,及時進行病蟲害防治。
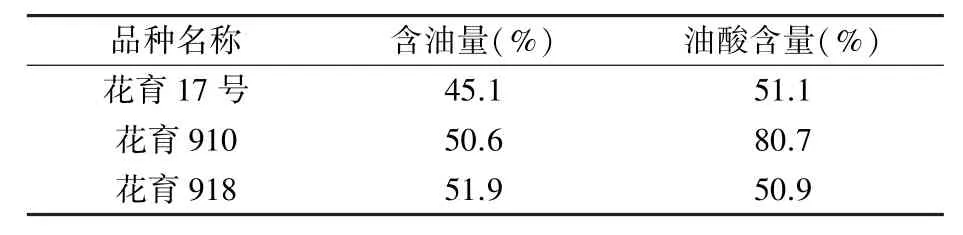
表1 供試花生品種油脂和油酸含量
1.2 取樣方法
花生開花下針后,分別采集下針后30、40、50、60 d 的莢果,剝取籽仁(圖1),用鋁箔紙包裹,經液氮速凍后立即放入-80℃冰箱保存備用。

圖1 供試材料4 個發育時期的籽仁
1.3 RNA 提取與cDNA 合成
采用北京天根生化科技有限公司試劑盒(RNeasy Mini Kit)提取樣品總RNA,詳細方法參考使用說明。 用DNaseⅠ處理提取的RNA 以去除DNA 污染。 利用Promega 的M-MLV 反轉錄酶進行cDNA 合成,每25 μL 反應體系中加入2 μg RNA;反轉錄體系于42℃反應1 h,之后置于冰上冷卻5 min,然后將反轉錄產物放于-20℃備用。
1.4 實時熒光定量PCR 分析
用SYBR Green PreMix 試劑盒(購自MDBio臺灣生工)按使用說明進行。 先將反轉錄所得的cDNA 稀釋到8 ng/μL,每反應體系中加2 μL 稀釋后的cDNA。 熒光定量PCR 儀采用Roche 的LightCycler 2.0。 反應程序:95℃10 s;95℃5 s,60℃30 s,72℃10 s,40 個循環;繪制熔解曲線,溫度每10 s 升高0. 5℃。 所用內參基因為Actin11,所用引物見表2。每個樣品設置3 次生物學重復。 采用delta-delta Cp 方法分析數據,誤差線為3 次重復的標準偏差。
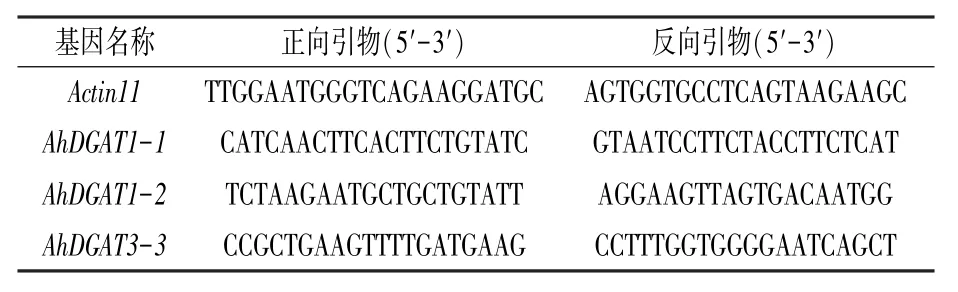
表2 熒光定量PCR 所用引物
2 結果與分析
2.1 AhDGAT1-1 基因的表達分析
隨著莢果發育,AhDGAT1-1基因在3 個花生品種籽仁中的表達模式各有不同(圖2)。 花育17 號與花育910 的AhDGAT1-1基因表達量均在下針后30 天(莢果發育初期)的籽仁中最高,在下針后40 天的籽仁中最低,下針后50 天又有小幅回升。 花育918 中,AhDGAT1-1基因表達量隨莢果發育顯著升高,在莢果發育中期(下針后40天)的籽仁中表達量最高,之后表達量逐漸降低。
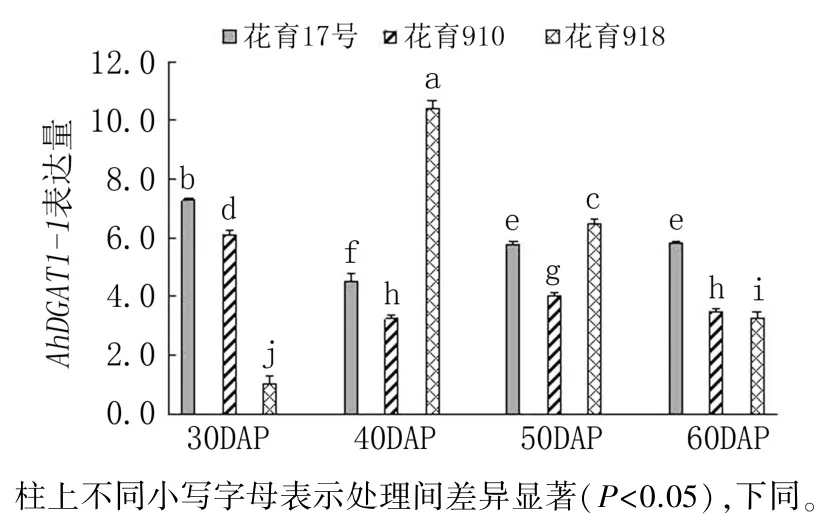
圖2 花生品種間AhDGAT1-1 基因表達差異分析
AhDGAT1-1基因表達量在下針后30 天和60天表現為花育17 號>花育910>花育918,而下針后40 天和50 天則表現為花育918>花育17 號>花育910。AhDGAT1-1在花育918 下針后40 天的籽仁中表達量最高,顯著高于其他時期各品種中的表達量;在花育17 號和花育910 下針后30天的籽仁中表達量也較高,與花育918 下針后50天的表達量相差較小。 說明AhDGAT1-1可能主要在低油低油酸品種和高油高油酸品種的莢果發育初期起作用,但在高油低油酸品種中則主要在籽仁油脂積累快速期。
2.2 花生AhDGAT1-2 基因的表達分析
由圖3 可見,隨著莢果發育,花育17 號中Ah-DGAT1-2基因的表達量呈降低-升高-降低的變化趨勢,最高值出現在下針后30 天。 花育910 中AhDGAT1-2基因的表達峰值出現在下針后50天,其次為下針后30 天,下針后40 天的表達量最低。 花育918 中AhDGAT1-2基因的表達量隨莢果發育先逐漸升高,下針后50 天達到峰值,之后急劇下降。 3 個品種間,下針后30 天和60 天,Ah-DGAT1-2基因的表達量均表現為花育17 號>花育910>花育918,品種間差異顯著;下針后40 天和50 天,AhDGAT1-2基因均在花育17 號中的表達量最高,其次為花育918,花育910 中的表達量最低,品種間差異也達顯著水平。 綜合來看,Ah-DGAT1-2基因的表達峰值出現在低油低油酸品種的籽仁充實初期,而出現在高油高油酸品種和高油低油酸品種的籽仁油脂積累快速期。
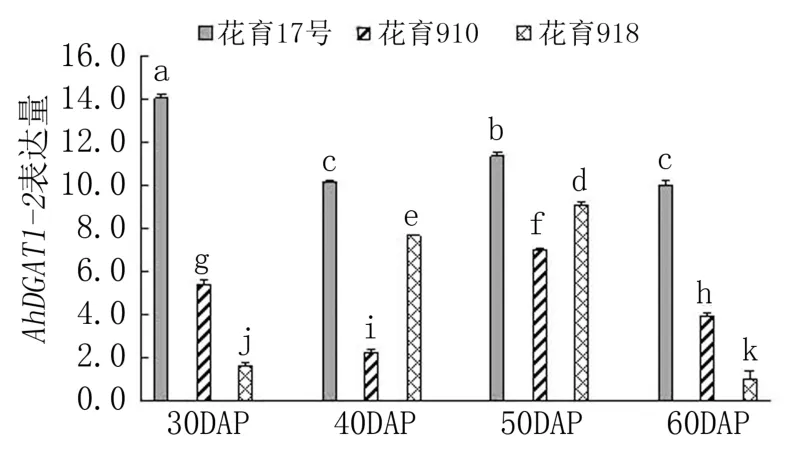
圖3 花生品種間AhDGAT1-2 基因表達差異分析
2.3 花生AhDGAT3-3 基因的表達分析
由圖4 可見,下針后30 天,AhDGAT3-3基因在花育910 中表達量最高,依次高于花育17 號和花育918;下針后40 天和50 天,花育17 號中Ah-DGAT3-3基因的表達量最高,花育918 中最低;下針后60 天,花育918 中AhDGAT3-3基因的表達量急劇升高,分別是花育17 號和花育910 表達量的4.65 倍和7.56 倍。 隨著莢果發育,AhDGAT3-3基因表達量在花育17 號和花育910 中均呈先升高后降低趨勢,均在下針后50 天達到峰值;而在花育918 中則呈持續升高趨勢,在下針后60 天達到峰值。 可見,AhDGAT3-3基因主要在3 個花生品種莢果發育中后期表達,說明其可能在種子儲藏物質積累方面起作用。
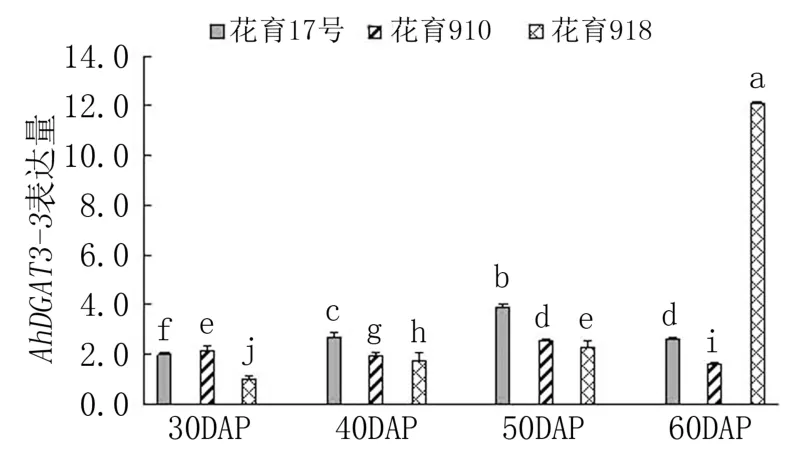
圖4 花生品種間AhDGAT3-3 基因表達差異分析
3 討論與結論
花生是我國重要的油料作物與經濟作物,提高含油量是花生品種改良的主攻方向之一。 由于花生基因組測序研究起步較晚,目前從分子水平研究花生油脂合成代謝機制的還較少。 越來越多的研究表明,DGAT 在真核生物TAG 生物合成的最后一步和限速步驟中起著至關重要的作用[20-22]。 對DGAT基因的研究近幾年來在一些油料作物如高油玉米、大豆、油菜、油桐(Vernicial fordii)和向日葵(Helianthus annuus) 中相繼開展[12,23-26]。
本研究對前期從花生中分離的3 個DGAT基因(AhDGAT1-1、AhDGAT1-2和AhDGAT3-3)在不同油脂含量的花生品種間的表達差異進行比較,結果表明,AhDGAT1-1基因表達峰值出現在低油低油酸品種和高油高油酸品種的莢果發育初期(下針后30 天),而出現在高油低油酸品種莢果發育中期(下針后40 天);AhDGAT1-2基因的表達峰值出現在低油低油酸品種的莢果發育初期及在高油高油酸品種和高油低油酸品種的籽仁油脂積累快速期;AhDGAT3-3基因則主要在3 個品種的莢果發育中后期表達。
研究發現不同植物的DGAT 對酰基脂肪酸底物選擇存在偏好性差異,致使不同物種的油脂成分不同,使特定脂肪酸在某些植物中富集,例如亞麻DGAT1 對亞麻酰CoA、油桐DGAT2 對桐酰CoA、可可(Theobroma cacao)DGAT1 對硬脂酰CoA 和花生DGAT3 對油酰基CoA(18 ∶1 CoA)的優先選擇[8,27-29]。 本研究結果顯示,在高油高油酸品種花育910 中AhDGAT1-1基因表達峰值出現在莢果發育初期,對促進花生籽仁油脂的合成有重要作用,但之后隨莢果發育表達量降低,這種降低是否與油酸含量升高有關,還需進一步的試驗驗證。AhDGAT1-2基因的表達峰值出現在低油低油酸品種花育17 號籽仁充實初期,而在高油高油酸品種花育910 和高油低油酸品種花育918的籽仁油脂積累快速期,說明該基因可能在種子發育中后期參與油脂與脂肪酸的合成與積累。DGAT3是一類組成型表達基因,于種子中優勢表達,在擬南芥、大豆、花生種子發育中期開始大量積累,一直延續到接近成熟階段[3,30]。 在本研究檢測的3 個花生品種中,AhDGAT3-3基因的表達峰值出現在莢果發育的中后期,也再次驗證了前人的試驗結果,說明該基因可能在種子儲藏物質積累方面起作用。
花生油脂合成途徑中關鍵酶基因的篩選及功能研究,對利用分子育種手段提高花生油脂含量意義重大。 本研究通過實時熒光定量PCR 檢測了3 個DGAT基因在高、低油花生品種中的表達,發現AhDGAT1-1主要在莢果發育早期起作用,而AhDGAT1-2和AhDGAT3-3主要在莢果發育中后期起作用。 本研究結果對深入理解花生油脂合成積累機制和調控網絡及選育高油花生品種具有一定的參考價值。
猜你喜歡
食品工業(2023年12期)2023-12-12 12:56:16
糧油食品科技(2022年6期)2022-11-25 12:18:34
糧油食品科技(2022年6期)2022-11-25 12:18:30
基層中醫藥(2020年8期)2020-11-16 00:55:20
環境衛生工程(2020年3期)2020-07-27 01:19:22
瘋狂英語·新讀寫(2020年3期)2020-06-06 09:06:14
塑料助劑(2019年3期)2019-07-24 08:51:22
中國糧油學報(2019年4期)2019-07-12 09:06:38
食品與機械(2018年11期)2019-01-03 06:15:34
中國糧油學報(2018年12期)2018-01-26 13:32:04
