豌豆酸漿中淀粉絮凝菌的篩選及其絮凝性質(zhì)分析
2023-07-29 10:19:54劉嬌李源王佳瑤張莉力顧英許云賀
食品工業(yè) 2023年7期
劉嬌,李源,王佳瑤,張莉力,顧英,許云賀
錦州醫(yī)科大學(xué)食品與健康學(xué)院(錦州 121000)
酸漿法生產(chǎn)淀粉在我國已有數(shù)百年的歷史,酸漿是將豆類磨碎成淀粉乳后放置一段時(shí)間經(jīng)微生物自然發(fā)酵,形成的一種淡黃色酸性液體[1]。酸漿中微生物的絮凝作用致使淀粉顆粒迅速形成大的絮凝團(tuán),從而加快淀粉與蛋白質(zhì)、纖維素等雜質(zhì)的分離。用酸漿法生產(chǎn)的淀粉被認(rèn)為比用離心等其他方法生產(chǎn)的淀粉更適合制作傳統(tǒng)的東方食品粉絲[2]。這種傳統(tǒng)的酸漿發(fā)酵方法仍在中國淀粉生產(chǎn)中占主導(dǎo)地位[3]。豌豆淀粉由于來源廣泛,價(jià)格便宜且成膠能力強(qiáng)、凝膠制品色澤好、持水性好等其他淀粉無法比擬的優(yōu)點(diǎn),被用于制作粉絲、粉皮、涼粉等傳統(tǒng)食品[4]。酸漿中的微生物類群對有效提升酸漿法的生產(chǎn)效率起關(guān)鍵性作用。但是很多因素限制了酸漿法在實(shí)際生產(chǎn)中的應(yīng)用,如環(huán)境因素會(huì)導(dǎo)致酸漿的酸度不易控制,酸漿中的腐敗雜菌也會(huì)致使不同批次淀粉產(chǎn)品的質(zhì)量各不相同。所以若將酸漿的生產(chǎn)由自然發(fā)酵改變?yōu)榧兎N發(fā)酵,將會(huì)解決眾多豌豆淀粉生產(chǎn)企業(yè)在酸漿發(fā)酵方面的技術(shù)問題。
絮凝作用菌的絮凝活性高低是其能否應(yīng)用到工業(yè)化生產(chǎn)中的重要評價(jià)指標(biāo)之一[5]。因此,試驗(yàn)利用平板分離法從豌豆酸漿中分離篩選出一株生長穩(wěn)定且具有高絮凝淀粉活性的菌株并對其絮凝性質(zhì)進(jìn)行分析,從而改善淀粉生產(chǎn)廢液排放導(dǎo)致的環(huán)境污染問題,減少淀粉的生產(chǎn)時(shí)間和酸漿的使用量,為豌豆酸漿接種發(fā)酵提供備選菌種及純種發(fā)酵、穩(wěn)定生產(chǎn)豌豆淀粉并研發(fā)淀粉專用微生物絮凝劑提供理論依據(jù)。
1 材料與方法
1.1 材料、試劑與培養(yǎng)基
新鮮豌豆(川南特產(chǎn)城網(wǎng)絡(luò)商店);豌豆淀粉(新良新鄉(xiāng)網(wǎng)絡(luò)商店);綠豆酸漿(微生物課題組自然發(fā)酵獲得);細(xì)菌基因組DNA提取試劑盒(沈陽立信生物技術(shù)有限公司);PCR引物(派森諾生物公司);葡萄糖、大豆蛋白胨、蔗糖、酵母浸粉、H2O2(天津市科密歐化學(xué)試劑有限公司);溶菌酶(Sigma-Aldrich公司);胃蛋白酶(雙匯實(shí)業(yè)集團(tuán)有限公司);胰蛋白酶(南寧龐博生物工程有限公司);纖維素酶(山東隆科特酶制劑有限公司);糖化酶(博立生物制品有限公司)。
豌豆汁制備方法:200 g新鮮豌豆加1 L蒸餾水加熱煮沸后,轉(zhuǎn)小火煮30 min,用0.125 mm孔徑(120目)篩子加8層紗布過濾,用蒸餾水定容至1 L備用。
基礎(chǔ)碗豆汁培養(yǎng)基:20 g葡萄糖、1 g大豆蛋白胨,碗豆汁1 L,調(diào)pH至6.5,于121 ℃滅菌30 min。
改良碗豆汁培養(yǎng)基:30 g蔗糖、0.5 g酵母浸粉,碗豆汁1 L,調(diào)pH至6.0,于121 ℃滅菌30 min。
UV-1600紫外可見分光光度計(jì)(北京瑞利分析儀器公司);T20MM立式高速冷凍離心機(jī)(湖南赫西儀器裝備有限公司);ABI-2720PCR儀(美國Applied Biosystems公司);Mini Pro 300V Power Supply電泳儀(major science USA);ABI 3730XL測序儀(美國Applied Biosystems公司)。
1.2 試驗(yàn)方法
1.2.1 豌豆乳的制備
稱取40 g豌豆洗凈,加入1 L自來水浸泡過夜(12~18 h),利用破壁機(jī)調(diào)節(jié)至4檔打漿10 s,使用0.125 mm孔徑(120目)篩子加8層紗布過濾后得到新鮮豌豆乳。
1.2.2 豌豆酸漿的制備
1.2.2.1 發(fā)酵豌豆酸漿的工藝
豌豆浸泡→打漿→新鮮豌豆乳→加老漿→靜置棄上清液→定容至1 L→密封培養(yǎng)(20~25 ℃)
1.2.2.2 酸漿的制備
新鮮豌豆乳中加入30%的綠豆發(fā)酵酸漿。棄2/3含有蛋白的上清液,用涼白開定容至1 L,置于室溫下發(fā)酵24 h。
1.2.2.3 酸漿的活化
將上述酸漿加入新制備的豌豆乳中。持續(xù)活化3個(gè)月,當(dāng)酸漿pH 4~5且漿液呈青白色并有少量水沫時(shí),結(jié)束發(fā)酵并檢測絮凝率,此時(shí)自然發(fā)酵的豌豆酸漿微生物區(qū)系趨于穩(wěn)定。
1.2.3 DNA的提取與高通量測序分析
取絮凝率最高時(shí)的豌豆酸漿,分為酸漿組(SQ1,SQ2和SQ3)和沉淀淀粉組(CD1,CD2和CD3),酸漿組是指豌豆全漿中的上清液,沉淀淀粉組是指豌豆全漿中的沉淀淀粉,對2組樣品進(jìn)行DNA的提取。PCR擴(kuò)增正向引物為520F,反向引物為802R。將合格的序列進(jìn)行文庫的擴(kuò)建并上機(jī)進(jìn)行高通量測序[6]。
1.2.4 ASV聚類及物種組成分析
利用Uparse軟件做ASVs聚類,代表序列為出現(xiàn)頻數(shù)最高的序列,利用分類數(shù)據(jù)庫在門和屬水平上統(tǒng)計(jì)樣品的物種組成。
1.2.5 菌株的分離與篩選
初篩。酸漿組和沉淀淀粉組分別用滅菌后的生理鹽水進(jìn)行梯度稀釋,選取適宜稀釋度涂布于MRS固體培養(yǎng)基平板上,挑取不同形態(tài)的單個(gè)菌落進(jìn)行活化,劃線接種于MRS斜面培養(yǎng)基上。以pH為指標(biāo),篩選出pH低于3.8的產(chǎn)酸菌株,對其斜面進(jìn)行保存。
復(fù)篩。將產(chǎn)酸菌株接種至碗豆汁培養(yǎng)基中35 ℃培養(yǎng)24 h。以絮凝率為指標(biāo)復(fù)篩出絮凝率大于45%的菌株。將菌株活化后再次梯度稀釋,重復(fù)劃線3次以上,直至鏡檢下菌落形態(tài)一致。
1.2.6 絮凝率(FR)測定
將5 g豌豆淀粉、100 mL蒸餾水和5 mL待測液依次加入100 mL量杯中,快速攪拌3 min后,靜置沉降3 min。在上清液面下10 mL處取樣。用紫外可見分光光度計(jì)測定其在550 nm波長處的吸光度,以空白豌豆汁培養(yǎng)基作對照試驗(yàn)[7-8],并用式(1)計(jì)算絮凝率。
絮凝率(FR)=(A-B)/A×100% (1)式中:A為對照組在550 nm波長處的吸光度;B為試驗(yàn)組在550 nm波長處的吸光度。
1.2.7 菌種鑒定
1.2.7.1 形態(tài)特征觀察
對絮凝率最高的菌株LJ5平板上的菌落形態(tài)進(jìn)行觀察并進(jìn)行革蘭氏染色試驗(yàn),使用光學(xué)顯微鏡對其個(gè)體形態(tài)進(jìn)行觀察。
1.2.7.2 菌種生理生化試驗(yàn)
對篩選出的絮凝優(yōu)勢菌LJ5進(jìn)行生理生化試驗(yàn),以鑒定細(xì)菌種屬。
1.2.7.3 16S rDNA的基因序列測定以及系統(tǒng)發(fā)育分析
細(xì)菌基因組PCR擴(kuò)增,擴(kuò)增正向引物為P1(27F)和反向引物為P2(1492R)。測序結(jié)果在GenBank中進(jìn)行BLAST比對,使用MEGA 10.0構(gòu)建絮凝優(yōu)勢菌的系統(tǒng)發(fā)育樹。
1.2.8 生長曲線繪制及培養(yǎng)時(shí)間確定
將S.harbinensisLJ5接種于改良碗豆汁培養(yǎng)基中,擴(kuò)大培養(yǎng)至第3代時(shí),間隔2 h取樣。用紫外可見分光光度計(jì)測定在600 nm波長處的吸光度,繪制生長曲線并確定最佳生長時(shí)間。采用平板計(jì)數(shù)法測定最佳生長時(shí)間的活菌數(shù)。
1.2.9 S.harbinensisLJ5絮凝性質(zhì)分析
1.2.9.1 S.harbinensisLJ5絮凝活性分布
為確定S.harbinensisLJ5起絮凝作用的是菌體分泌物還是菌體細(xì)胞本身,利用光學(xué)顯微鏡和掃描電子顯微鏡觀察加入改良碗豆汁發(fā)酵液前后豌豆淀粉顆粒的分布狀態(tài)。對S.harbinensisLJ5發(fā)酵液、上清液、未洗滌菌懸液和洗滌菌懸液的絮凝率進(jìn)行測定。發(fā)酵液為S.harbinensisLJ5在35 ℃培養(yǎng)36 h的碗豆汁發(fā)酵液;上清液為碗豆汁發(fā)酵液在10 000 r/min條件下離心10 min后的上清液;未洗滌菌懸液為碗豆汁發(fā)酵液離心后的菌體細(xì)胞加入等體積的無菌蒸餾水;洗滌菌懸液為碗豆汁發(fā)酵液離心后洗滌菌體細(xì)胞2次,再次加入等體積的無菌蒸餾水。
1.2.9.2 S.harbinensisLJ5絮凝活性的熱穩(wěn)定性分析
分別測定30,40,50,60和70 ℃條件下水浴30 min后的S.harbinensisLJ5發(fā)酵液對豌豆淀粉懸濁液的絮凝率。
1.2.9.3 酶處理對S.harbinensisLJ5發(fā)酵液絮凝活性的影響
發(fā)酵液在4 ℃條件下以8 000 r/min離心20 min,收集菌體,用pH 7.0的磷酸鹽緩沖液(0.2 mol/L NaH2PO4與0.2 mol/L Na2HPO4一定比例混合)離心洗滌菌體數(shù)次,至菌體呈潔凈白色為止,收集菌體備用[9]。準(zhǔn)確稱取1.0 g菌體,用磷酸鹽緩沖液懸浮菌體,分別加入0.3%酶質(zhì)量濃度1 mg/L的溶菌酶、胰蛋白酶、胃蛋白酶、纖維素酶和糖化酶,酶解pH分別為7.8,8.0,2.5,4.8和4.5,酶解溫度分別為52,37,37,50和60℃,放在200 r/min的搖床上酶解2 h[10]。以未處理的發(fā)酵液為對照組,分別測定各試驗(yàn)組的絮凝率。
1.2.9.4 金屬離子對S.harbinensisLJ5發(fā)酵液絮凝活性的影響
淀粉懸濁液本身帶負(fù)電性,所以選擇陽離子進(jìn)行絮凝試驗(yàn)。配制1%濃度的各鹽溶液:KCl、NaCl、CaCl2和MgCl2。將發(fā)酵液與1 mL各鹽溶液一起加入100 mL的豌豆淀粉懸濁液,分別測定各試驗(yàn)組的絮凝率[11]。
1.3 數(shù)據(jù)處理
利用SPSS 26.0、MEGA 10.0和Origin 2021a軟件進(jìn)行數(shù)據(jù)分析和繪圖。
2 結(jié)果與分析
2.1 豌豆酸漿樣品中細(xì)菌群落的稀疏曲線
稀疏曲線可以判斷測序數(shù)量能否反映樣本中物種的組成以及測序深度和物種的豐富度程度[12-13]。隨著測序數(shù)量不斷增加,曲線呈快速上升趨勢,表明還有大量物種未被發(fā)現(xiàn),而曲線平穩(wěn)時(shí)表明測序深度已經(jīng)飽和且基本覆蓋到樣本中所有的物種及絕大部分的微生物信息[14-15]。如圖1所示,測序數(shù)量超過20 000時(shí),2個(gè)樣本顯示出趨于平坦的稀疏曲線,表明隨著測序深度不斷增加幾乎不會(huì)有新的物種產(chǎn)生,因此試驗(yàn)的測序量合理[16-17]。
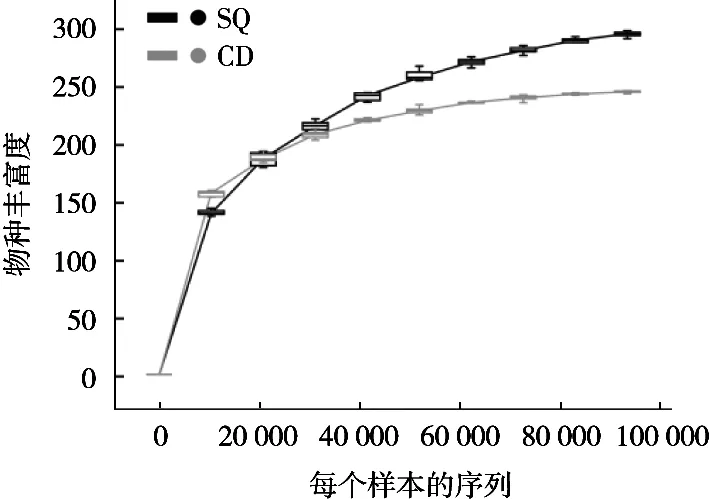
圖1 細(xì)菌群落稀疏曲線分析
2.2 豌豆酸漿樣品中細(xì)菌多樣性分析
由圖2可知,韋恩圖結(jié)果共得到1 088個(gè)ASVs,酸漿組中含637個(gè)ASVs,沉淀淀粉組中含451個(gè)ASVs,其中2組共有的ASVs數(shù)為88個(gè),2組特有的ASVs分別為549和363個(gè),表明酸漿組中微生物群落較沉淀淀粉組豐富。2組樣品多樣性指數(shù)分析見表1。酸漿組和沉淀淀粉組的細(xì)菌區(qū)系,分別獲得168 902和125 097條有效序列。Chao1指數(shù)可估計(jì)樣本群落中包含的物種總數(shù),物種總數(shù)越多值越大[18]。因此,酸漿組中的物種總數(shù)較沉淀淀粉組的物種總數(shù)多。酸漿組的Shannon、Simpson指數(shù)顯著(P<0.05)高于沉淀淀粉組。結(jié)果表明,酸漿組的細(xì)菌豐富度及多樣性更高。

表1 2種樣品中細(xì)菌Alpha多樣性指數(shù)

圖2 2種樣品的ASVs聚類
2.3 豌豆酸漿樣品中細(xì)菌群落結(jié)構(gòu)差異分析
2.3.1 門水平下豌豆酸漿樣品中細(xì)菌群落結(jié)構(gòu)分析
由圖3可知,在門水平下,酸漿組與沉淀淀粉組共鑒定出8個(gè)門類細(xì)菌。酸漿組中變形菌門(Pro-teobacteria)、厚壁菌門(Firmicutes)、擬桿菌門(Bacteroidetes)和放線菌門(Actinobacteria),所占比例依次為56.92%,38.35%,3.80%和0.91%;沉淀淀粉組中厚壁菌門(Firmicutes)、放線菌門(Actinobacteria)、變形菌門(Proteobacteria)和藍(lán)藻菌門(Cyanobacteria),所占比例依次為55.84%,41.03%,1.76%和0.98%。酸漿組中的優(yōu)勢菌門為變形菌門,沉淀淀粉組中的優(yōu)勢菌門為厚壁菌門。
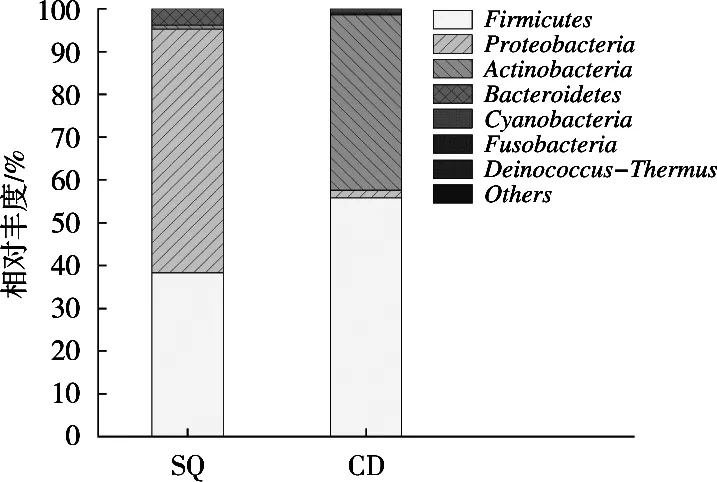
圖3 基于門水平的細(xì)菌結(jié)構(gòu)
2.3.2 屬水平下豌豆酸漿樣品中細(xì)菌群落結(jié)構(gòu)分析
由圖4可知,在屬水平下,酸漿組與沉淀淀粉組的細(xì)菌隸屬于8個(gè)屬,酸漿組中醋酸桿菌屬(Acetobacter)、乳酸桿菌屬(Lactobacillus)和乳球菌屬(Lactococcus)所占比例依次為55.70%,25.12%和10.13%;沉淀淀粉組中乳酸桿菌屬(Lactobacillus)、雙歧桿菌屬(Bifidobacterium)和醋酸桿菌屬(Acetobacter)所占比例依次為55.60%,41.03%和1.52%。酸漿組中的優(yōu)勢菌屬為醋酸桿菌屬,沉淀淀粉組中的優(yōu)勢菌屬為乳酸桿菌屬。
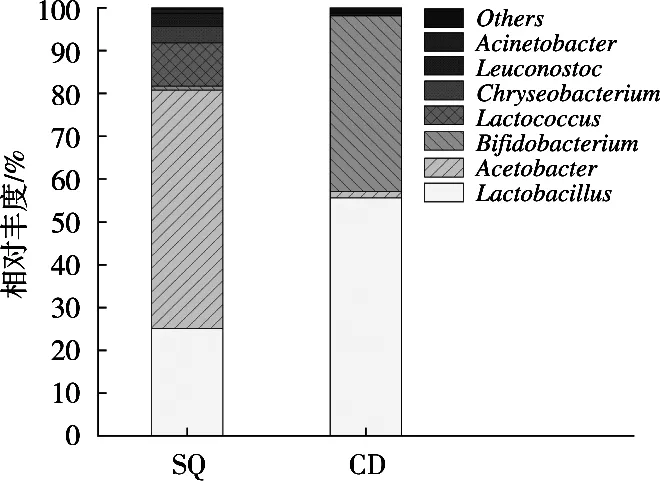
圖4 基于屬水平的細(xì)菌結(jié)構(gòu)
2.4 菌株的分離與篩選
以pH為指標(biāo)從自然發(fā)酵豌豆酸漿樣品中初篩共分離出36株pH低于3.8的菌株,以絮凝率為指標(biāo)復(fù)篩出6株絮凝率大于45%的菌株,結(jié)果見表2。選擇絮凝率最高達(dá)71.2%±2.0%的菌株LJ5作為后續(xù)試驗(yàn)的研究對象。

表2 由豌豆酸漿分離出的6株菌對淀粉的絮凝率
2.5 菌種鑒定
對絮凝活性較高的菌株LJ5進(jìn)行菌種鑒定。菌株LJ5的菌落形態(tài)見圖5(A),光學(xué)顯微鏡下形態(tài)見圖5(B)。
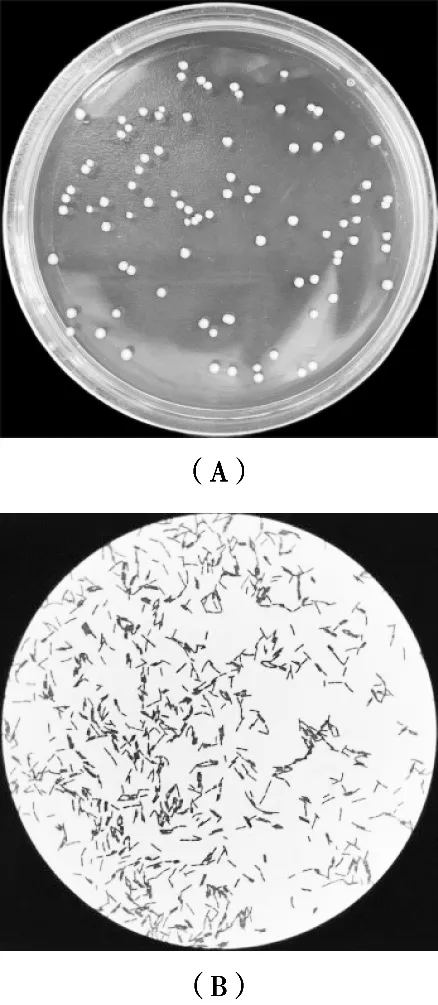
圖5 菌株LJ5菌落形態(tài)及其革蘭氏染色菌體形態(tài)
2.5.1 形態(tài)特征觀察
通過光學(xué)顯微鏡觀察絮凝優(yōu)勢菌LJ5在MRS培養(yǎng)基中的形態(tài)和菌落特征。菌株LJ5的菌體大小為0.5 μm×2.5 μm,桿狀,鏈狀排布方式、革蘭氏陽性、無芽孢、不運(yùn)動(dòng)。菌株LJ5的菌落大小為1~3 mm,菌落形態(tài)為灰白色圓形、不透明、低凸隆起有光澤、邊緣不整齊、表面不光滑。
2.5.2 生理生化試驗(yàn)
試驗(yàn)結(jié)果見表3,初步判斷絮凝優(yōu)勢菌LJ5為乳酸桿菌屬。

表3 菌株LJ5生理生化試驗(yàn)結(jié)果
2.5.3 16 S rDNA序列分析結(jié)果及系統(tǒng)發(fā)育樹的構(gòu)建
6株絮凝菌純化后的16S rDNA的PCR產(chǎn)物瓊脂糖凝膠電泳結(jié)果如圖6所示。使用測序儀ABI 3730XL進(jìn)行DNA測序。用NCBI中的BLAST程序?qū)⑵唇雍蟮男蛄形募cNCBI中16S rDNA數(shù)據(jù)庫中的數(shù)據(jù)進(jìn)行比對,選擇相似度最大的序列作為物種鑒定的結(jié)果[19]。
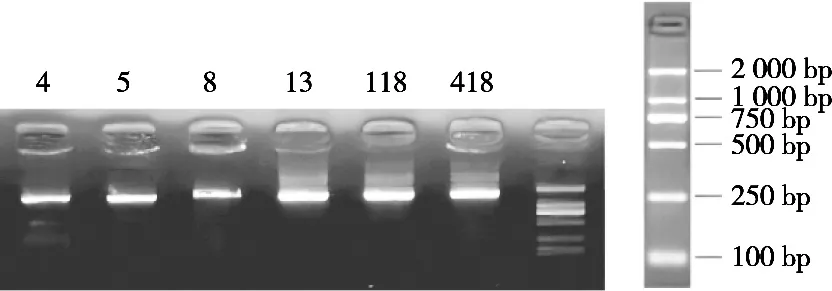
圖6 菌株LJ5的16S rDNA電泳圖譜
經(jīng)16S rDNA鑒定,菌株LJ5的16S rDNA序列全長為1 451 bp,經(jīng)GenBank比對,與Schleiferilactobacillus harbinensis相似性達(dá)到100%。利用MEGA 10.0軟件構(gòu)建系統(tǒng)發(fā)育樹,菌株LJ5與Schleiferilactobacillus harbinensisOK271739.1親緣關(guān)系最近,結(jié)果如圖7所示。結(jié)合其群體、個(gè)體形態(tài)特征、生理生化試驗(yàn)及16S rDNA序列比對,確定菌株LJ5為哈爾濱施萊弗乳桿菌,命名為Schleiferilactobacillus harbinensisLJ5。

圖7 菌株LJ5基因序列系統(tǒng)發(fā)育樹
2.6 生長曲線的測定
生長曲線如圖8所示。接種0~4 h為延滯期,4~24 h為對數(shù)生長期,24~58 h為穩(wěn)定期,之后為衰亡期。結(jié)合S.harbinensisLJ5實(shí)際用途是用于豌豆酸漿的純種發(fā)酵,為獲得高絮凝淀粉活性的豌豆酸漿,對于菌株種子液的活菌數(shù)和活力均要求較高,所以選擇對數(shù)期末期、穩(wěn)定期前期為發(fā)酵終點(diǎn),最佳發(fā)酵時(shí)間為24 h。發(fā)酵24 h時(shí)菌液的活菌數(shù)為8.31±0.03 lg CFU/mL。

圖8 S.harbinensis LJ5生長曲線
2.7 S.harbinensis LJ5絮凝性質(zhì)分析
2.7.1 S.harbinensisLJ5絮凝活性物質(zhì)分布
普遍認(rèn)為微生物絮凝劑分為兩類:一類微生物分泌的胞外產(chǎn)物具有絮凝作用,如發(fā)酵乳桿菌的分泌物,其胞外產(chǎn)物具有絮凝作用,而菌體本身沒有絮凝作用[20];另一類是菌體本身具有絮凝作用,如副干酪乳桿菌副干酪亞種[21]。光學(xué)顯微鏡下觀察到加入發(fā)酵液前后豌豆淀粉顆粒的分布狀態(tài)如圖9(A和B)所示。加入發(fā)酵液前,淀粉顆粒在顯微鏡下均勻分布,加入發(fā)酵液后,眾多淀粉顆粒凝聚成大的絮凝體。如圖9(C和D)所示,通過掃描電子顯微鏡可以觀察到加入發(fā)酵液前后淀粉顆粒的變化,菌粘附在淀粉顆粒表面并且使淀粉顆粒粘結(jié)在一起,所以在絮凝淀粉過程中使淀粉顆粒凝集成大的絮凝體,正是淀粉顆粒粘結(jié)體積變大后重力的增加,從而加速淀粉的沉降。發(fā)酵液絮凝位置分析圖如圖9(E)所示。絮凝率大小順序?yàn)榘l(fā)酵液>未洗滌菌懸液>洗滌菌懸液>上清液。其中,未洗滌菌懸液和洗滌菌懸液與發(fā)酵液的絮凝率差異較小,都具有很強(qiáng)的絮凝活性,可能是由于離心分離過程損失一部分的菌體細(xì)胞,使菌體細(xì)胞數(shù)量減少,導(dǎo)致絮凝活性的稍有下降。而上清液與發(fā)酵液的絮凝率差異較大,去細(xì)胞后的上清液基本不具有絮凝活性,表明S.harbinensisLJ5的絮凝活性物質(zhì)主要存在于菌體細(xì)胞本身,而不是分泌到胞外的代謝產(chǎn)物。結(jié)合光學(xué)顯微鏡和電子顯微鏡下S.harbinensisLJ5粘附淀粉顆粒的特性與絮凝活性物質(zhì)存在于菌體細(xì)胞本身,表明絮凝因子可能在菌體細(xì)胞的表面。

圖9 豌豆淀粉顆粒分布及絮凝位置分析圖
2.7.2 S.harbinensisLJ5絮凝活性的熱穩(wěn)定性分析
從圖10可以看出,溫度對S.harbinensisLJ5的絮凝活性影響很大,在30~40 ℃范圍內(nèi),發(fā)酵液的絮凝活性從71.5%降至68.5%,而溫度大于50 ℃時(shí),發(fā)酵液基本不具有絮凝活性,表明S.harbinensisLJ5發(fā)酵液的絮凝因子不具有熱穩(wěn)定性,對溫度較為敏感。

圖10 發(fā)酵液絮凝活性的熱穩(wěn)定性分析圖
2.7.3 酶處理對S.harbinensisLJ5發(fā)酵液絮凝活性的影響
由圖11可知,空白組的絮凝率最高,為71.1%。溶菌酶、胰蛋白酶與胃蛋白酶3種酶對發(fā)酵液的絮凝活性都有不同程度的抑制作用,其中胰蛋白酶和胃蛋白酶的抑制作用較大,而用纖維素酶和糖化酶作用后的絮凝率基本沒有變化,表明S.harbinensisLJ5絮凝因子可能是蛋白類物質(zhì)。結(jié)合上述試驗(yàn)結(jié)果推測絮凝因子可能為菌體細(xì)胞表面的蛋白類物質(zhì)。
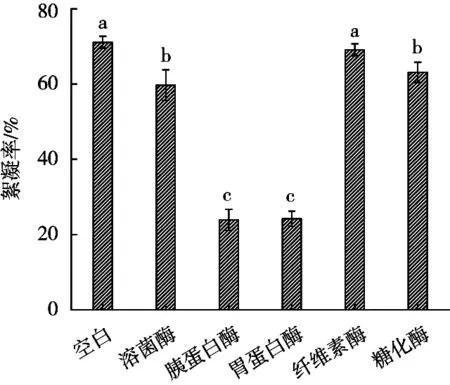
圖11 酶處理對發(fā)酵液絮凝活性的影響
2.7.4 金屬離子對S.harbinensisLJ5發(fā)酵液絮凝活性的影響
膠體理論表明陽離子可以和帶負(fù)電性膠體微粒的表面電荷結(jié)合,從而克服靜電排斥力使膠體顆粒脫穩(wěn)形成細(xì)小的凝聚體并與微生物絮凝劑反應(yīng)形成大絮凝團(tuán)快速下沉。不同金屬陽離子對發(fā)酵液絮凝活性的影響如圖12所示。Na+、K+對發(fā)酵液的絮凝活性均有增強(qiáng)作用,Ca2+、Mg2+對絮凝活性也有不同程度的促進(jìn)作用。其中,Na+的促進(jìn)作用最強(qiáng),所以可選Na+作為助凝劑。
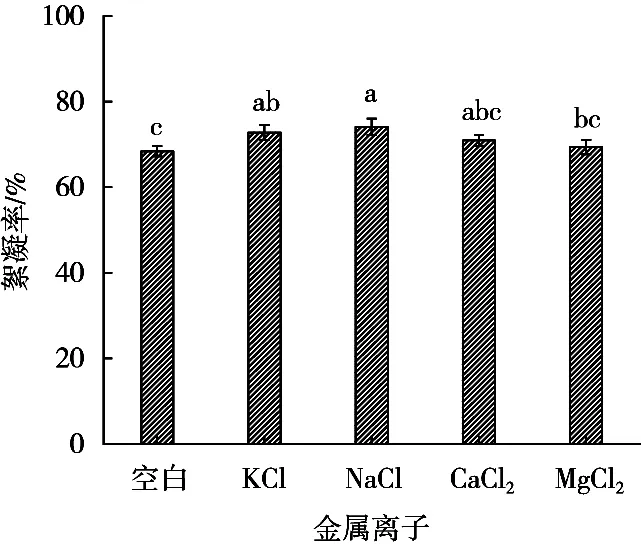
圖12 金屬離子對發(fā)酵液絮凝活性的影響
3 結(jié)論
通過高通量測序技術(shù)分析酸漿組和沉淀淀粉組中的微生物多樣性,從中分離篩選出一株在碗豆汁培養(yǎng)基中生長穩(wěn)定且具有高絮凝淀粉活性的S.harbinensisLJ5,為豌豆酸漿接種發(fā)酵提供備選菌種。通過絮凝活性位置分析確定絮凝活性物質(zhì)主要存在于菌體細(xì)胞本身。菌株發(fā)酵液的絮凝活性受溫度影響顯著,絮凝活性在30 ℃時(shí)最高,而溫度在50 ℃以上時(shí),發(fā)酵液就基本不具有絮凝活性,絮凝因子對溫度比較敏感。胰蛋白酶和胃蛋白酶對發(fā)酵液的抑制作用較強(qiáng),說明絮凝活性物質(zhì)可能是蛋白類的物質(zhì)。Na+對發(fā)酵液的絮凝活性促進(jìn)作用最強(qiáng),可選Na+作為助凝劑。
