呼倫貝爾沙化草地不同生態修復階段植物群落特征研究
2023-07-29 13:55:28郝顥晶關瀟曹明李俊生
環境工程技術學報 2023年4期
關鍵詞:物種
郝顥晶,關瀟,曹明,李俊生,2*
1.中國環境科學研究院
2.中國地質調查局自然資源綜合調查指揮中心
草地資源作為我國面積最大的陸地生態系統,對水土保持、水源涵養、氣候調節等生態系統服務的維持及生物多樣性保護均具有重要意義[1]。呼倫貝爾草原作為我國北方主要自然資源及生態防護屏障,具有獨特的地理位置和多樣的生態功能。隨著全球氣候變化[2]和人類活動強度的持續加深[3-4],呼倫貝爾草原面臨日益嚴峻的沙化問題[5],威脅著草地生態系統的可持續發展[6]。近年來,呼倫貝爾市加大沙地植被修復治理力度,開展了人工植被修復等一系列草地修復和管理保護措施[7],草地生態環境得到初步修復。甘珠爾沙地位于呼倫貝爾市西南部,地處呼倫貝爾典型沙帶之一,其生態區位重要,生態敏感性等級為脆弱區。自2005年呼倫貝爾市采用“圍欄封育+建立草方格+補播耐旱植物”的林草、灌草結合的方式對甘珠爾沙地的流動沙丘開展修復工程后,流動沙丘被固定,植被蓋度得到明顯改善[8-9]。基于此,為促進沙化草地治理進程和提高治理成效,對甘珠爾不同修復年限的沙地開展植物群落研究,具有重要的現實意義。
不同立地條件下,沙化草地人工修復植物群落表現出不同的時間梯度特征。如錢洲等[10]對毛烏素沙地不同修復年限的植被修復特征的研究發現,密度、蓋度及生物量有明顯增長,同時植被和土壤性質之間存在極為密切的相互促進的相關性;慕宗杰[11]發現,渾善達克沙地不同治理年份的植物群落中灌木層與草本層高度、蓋度、多樣性的變化趨勢和波動幅度存在差異;王明明等[12]對長期封育的科爾沁沙地的植物研究中發現,隨著修復年限的增加植被蓋度顯著提高,生物量和多樣性在年際間波動變化無顯著趨勢;栗忠飛等[13]在呼倫貝爾南部沙帶植被修復過程中發現,隨植被修復年限的增加地上植被生物量、蓋度顯著增加;金曉明等[14]對呼倫貝爾市陳巴爾虎旗人工修復后的沙化草地的研究發現,植物群落在修復初期的1~4 a 仍處于不穩定狀態。綜上,沙地的植被修復的動態變化受到沙地地理位置、修復年限、土壤條件及區域氣候等因素的影響,在植被物種組成、結構和多樣性等方面表現出差異。
目前關于不同修復年限下呼倫貝爾市甘珠爾沙地的植被研究較為鮮見,針對甘珠爾沙地在長期修復過程中植物群落特征變化、多樣性變化及其影響因素的探究還較缺乏。因此,筆者采用以空代時法,以甘珠爾沙地修復區內修復年限分別為9、11、13、15 a 的沙化草地作為研究對象,結合同期氣象數據與土壤養分測定數據,探討分析不同修復年限沙地植物群落特征、多樣性變化及其影響因素,有助于深入理解沙化草地植物群落對修復過程的響應,以期為沙化草地的植被修復和生態環境改善提供科學指導。
1 材料與方法
1.1 研究區概況
甘珠爾沙地位于內蒙古自治區呼倫貝爾市新巴爾虎左旗阿木古郎鎮(117°55′E~119°22′E,47°59′N~48°43′N)(圖1),土壤類型為沙質栗鈣土[15]。平均海拔700 m,屬于溫帶半干旱大陸性季風氣候,年均氣溫0.3 ℃,年日照時數為3 076 h,無霜期100 d,積雪期140 d,年主導風向為西北風,年均風速2.8 m/s,年均降水量約287 mm,主要集中在7—8 月,年均蒸發量2 650 mm,雨熱同期,有利于夏季植物生長[16]。
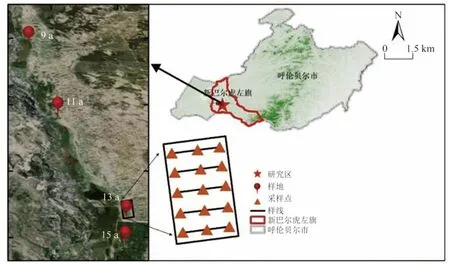
圖1 研究區采樣點示意Fig.1 Location map of the study area and sampling points
進行人工植被修復前,甘珠爾沙地的地貌特征為流動沙丘。自2005年起,甘珠爾沙地開展人工植被修復工作,以灌草混播的方式為主,輔以少量樟子松(Pinus sylvestris)固定流動沙丘并在人工播種后對甘珠爾沙地修復區實施圍欄封育。各樣地均保持獨立封育,期間可排除家畜采食、踐踏、施肥等人工干擾,使土壤內養分貯存從而促進修復區內植被自然更新和生產力恢復。開展本研究時,修復區已分布著大量不同修復年限的人工植被,灌木有羊柴(Hedysarum laeve)、小葉錦雞兒(Caragana microphylla)、鹽蒿(Artemisia halodendron),其下草本以羊草(Leymus chinensis)、 披堿草(Elymus dahuricus)、糙隱子草(Cleistogenes squarrosa)、地稍瓜(Cynanchum thesioides)、 蟲實(Corispermum hyssopifolium)等植物為主。
1.2 試驗設計與數據采集
1.2.1 樣地設置
通過實地調查,分別選取甘珠爾沙地修復區內地形地貌、成土母質等土壤環境條件相近,修復起始年限不同的4 個修復樣地(2006年、2008年、2010年和2012年),開展植物群落調查及土壤樣品的采集。按照修復年限的不同將樣地命名為15、13、11、9 a。為克服邊緣效應對結果的影響,各樣地均距離圍欄邊界20 m 以上,各樣地內自西向東沿植被種植時樣帶走向,均勻設置5 條平行樣帶(相鄰樣帶間隔50 m 以上),每條樣帶上設置3 個重復,每個重復包含5 m × 5 m 灌木樣方和1 m × 1 m 草本樣方各1 個,共計草本植物樣方60 個,灌木樣方60 個。
1.2.2 植被調查與土壤采集
2021年8 月下旬,在沙地植被生長旺盛期分別對15、13、11、9 a 人工修復的植被進行采樣調查,包括植物群落特征調查和土壤樣品采集。野外調查與數據計算參考任繼周[17]草原生態調查方法,利用鋼卷尺進行測量并記錄樣方內出現的草本及灌木植物種類、株數、株高、多度、基徑、冠幅、分蓋度、群落總蓋度、頻度。齊地刈割樣方內的草樣,分裝后帶回室內105 ℃殺青30 min,65 ℃烘干至恒重再稱取其地上生物量。計算相對密度(RD)、相對蓋度(RV)、相對頻度(RF)及重要值(IV)。在刈割后的草本樣方內,用直徑3 cm 的土鉆采集0~20 cm 深度的土壤樣品,每個樣方內采用對角線五點取樣法采集5 個土壤樣品并混合均勻,帶回室內風干磨碎并過篩。
1.2.3 氣象數據
研究區2006—2012年的年均降水量、蒸散發、風速和溫度等氣象數據來源于國家地球系統科學數據中心(http://www.geodata.cn)。
1.3 測定方法
1.3.1 主要植物群落結構特征值測定
根據已有研究[18-21]得到的生物量模型計算灌木物種地上生物量:
式中:W1、W2 、W3、W4 分別 為羊柴、黃柳、小葉錦雞兒及鹽蒿物種的地上生物量,kg/hm2; DH為斷面周長,m;V為冠幅體積,m3;A為冠幅面積,m2。
式中:IV 為各物種重要值;RD 為相對密度,%;RV為相對蓋度,%;RF 為相對頻度,%。
物種多樣性指數主要包括Shannon-Wiener 指數( Sw)、Simpson 指數(D)、Margalef 指數(M)以及Pielou 指數(J),計算公式如下:
式中:Pi為第i物種密度占群落總密度的比例;S為樣方物種總數;T為樣方中所有物種的個體總數。
1.3.2 土壤測定
參考鮑士旦[22]的土壤分析方法,測定土壤含水量、容重、pH、電導率、總氮含量、總鉀含量、總磷含量和有機碳含量。采用烘干稱重法測定土壤含水量;環刀法測定土壤容重;電極電位法測定土壤pH 和電導率;凱氏定氮法測定土壤總氮含量;NaOH熔融-原子吸收法測定土壤總鉀含量;NaOH 熔融-鉬銻抗顯色-紫外分光光度法測定土壤總磷含量;重鉻酸鉀容量法-外加熱法測定土壤有機碳含量。
1.4 數據處理
采用Excel 軟件對不同修復年限的植物調查數據、環境因子數據進行預處理,基于R 4.1.2 語言進行多樣性指數計算及單因素方差分析,并基于Tukey HSD 檢驗對其進行多重比較。利用R 4.1.2 語言的randomForest 包對物種多樣性與環境因子之間的相互關系進行隨機森林模型回歸分析,基于rfPermute包檢驗模型中各環境因子的顯著性[23],A3 包檢驗整個模型的顯著性[24],通過部分依賴圖展現具有顯著性的環境因子與生物多樣性指數之間的關系。采用Origin 8.0 軟件制圖。
2 結果與分析
2.1 不同物種組成及優勢度
2.1.1 植物科、屬、種和生活型組成
研究區域內不同修復階段的植物群落中,共發現42 種植物,隸屬于17 科35 屬。其中灌木物種共3 種,分別為羊柴(Hedysarum fruticosum)、小葉錦雞兒(Caragana microphylla)和鹽蒿(Artemisia halodendron);草本植物39 種,隸屬于17 科33 屬,物種組成以菊科(Asteraceae)、禾本科(Poaceae)、薔薇科(Rosaceae)和豆科(Leguminosae)為主,占總數的56.4%。藜科(Chenopodiaceae)、蓼科(Polygonaceae)、石竹科(Caryophyllaceae) 及莧科(Amaranthaceae)共8 個物種,占總數的20.5%。單屬種9 種,占總數的23.1%。按生活型劃分,可分為一年生草本9 種、一/二年生草本2 種和多年生草本28 種。多年生草本物種數明顯高于一年生和一/二年生,占比為43.8%~71.4%。
不同修復階段,群落物種數存在一定差異。草本物種數整體呈現先下降后上升的趨勢,表現為26 種(15 a)、23 種(13 a)、16 種(9 a)、13 種(11 a),在各修復年限中,一/二年生草本占草本物種總數的占比與其他生活型相比最小;灌木物種數以豆科為主,在不同修復階段均有分布。菊科的鹽蒿物種僅在9 和11 a 中出現。
2.1.2 物種重要值變化
植物物種的重要值可反映出各修復年限草地植被的整體情況。結果顯示,各物種重要值在年際間波動劇烈(圖2、圖3)。草本植物中地梢瓜(Cynanchum thesioides)、蟲實(Corispermum hyssopifolium)、披堿草(Elymus dahuricus)、 糙隱子草(Cleistogenes squarrosa)、 羊草(Leymus chinensis)與狗尾草(Setaria viridis)的重要值明顯高于其他物種。9 a 時草本優勢度較高的物種依次為地梢瓜(18.3%)、蟲實(14.8%)、披堿草(11.9%) 和羊草(11.0%)。同期糙隱子草(3.69%)并未表現出優勢。11 a 時地梢瓜(29.4%)與披堿草(22.0%)重要值顯著高于其他草本物種。13 a 時地梢瓜被糙隱子草(15.6%)和狗尾草(15.1%)取代,與此同時黃花蒿(Artemisia annua)、羊草與蟲實的重要值均超過10.0%,與糙隱子草和狗尾草共同組成該年的優勢群落。15 a 時披堿草與狗尾草成為草本最優勢物種,重要值分別為18.1%和10.5%。
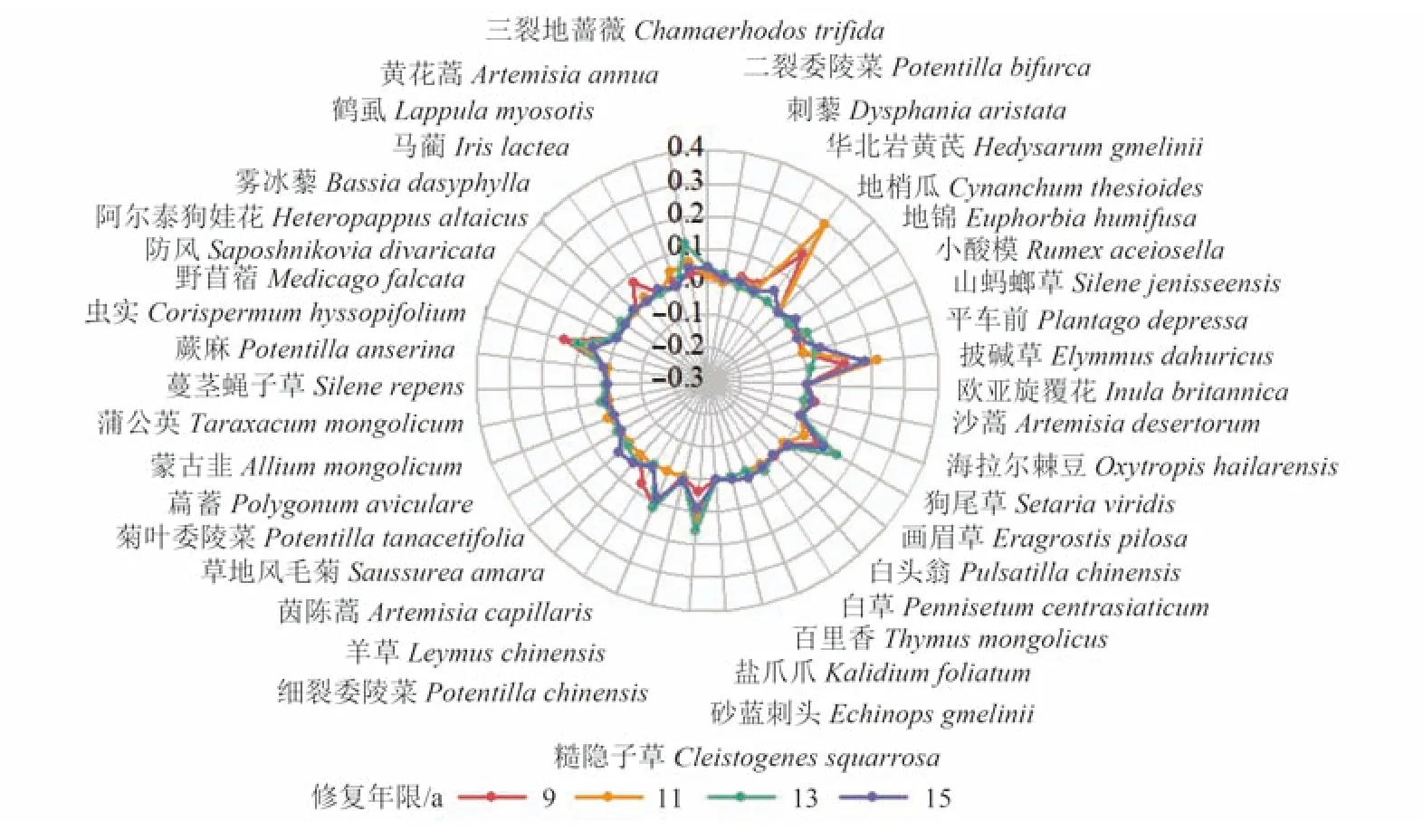
圖2 不同修復年限研究區草本植物重要值雷達圖Fig.2 Radar map of important values of herbaceous plants in the study area with different recovery years

圖3 不同修復年限研究區灌木植物重要值雷達圖Fig.3 Radar map of important values of shrub plants in the study area with different recovery years
灌木物種小葉錦雞兒在各修復年限間整體表現出具有較高的優勢度,其在9 a 時重要值最大為74.8%。11 a 時鹽蒿物種分別超過羊柴(26.2%)和小葉錦雞兒(21.0%)達到最大值(52.8%)。13 a 時小葉錦雞兒重新成為最優勢物種,重要值達到73.3%。15 a 時,未發現鹽蒿物種存在,2 種豆科植物羊柴(51.7%)和小葉錦雞兒(48.3%)共同構成當年的灌木群落。
2.2 蓋度、生物量變化
隨著修復年限的增加,草本植被蓋度整體呈波動增加的趨勢,但差異不顯著(P>0.05) 〔圖4(a)〕。其中修復年限最短的(9 a)總蓋度最小(30.5%),在11 a 時植被蓋度升至43.0%,在13 a 時降至34.9%,在修復年限最長(15 a)時達到最大值(43.3%)。與修復9 a 相比,11、13 和15 a 草本總蓋度分別高出41.2%、14.4%和42.2%。草本地上生物量在不同修復年限之間無顯著差異(P>0.05),但與蓋度呈現相似的波動趨勢。其最大值出現在15 a(33.9 g/m2) ,與9 a的最小值(18.1 g/m2)相比增加了87.0%〔圖4(b)〕。

圖4 甘珠爾沙地不同修復年限植被蓋度、地上生物量比較Fig.4 Comparison of vegetation coverage and aboveground biomass in different restoration years in Ganzhuer sandy land
灌木總蓋度隨著修復年限的增加而增加,在修復年限最短的9 a(38.7%)和最長的15 a(72.6%)之間存在顯著性差異(P<0.05),在其他修復年限之間差異不顯著〔圖4(a)〕。灌木地上生物量在修復9 a 與其他修復年限(11、13、15 a)之間均存在顯著性差異(P<0.05),而11、13、15 a 之間差異不顯著(P>0.05)〔圖4(b)〕。總體來看,灌木地上生物量隨修復年限的增加呈現持續增加的趨勢。9 a 為最小值(6.75 kg/m2),11、13 、15 a 相比于9 a 分別顯著增加了1.81 倍、1.88 倍和2.10 倍(P<0.05),15 a 為最大值20.9 kg/m2〔圖4(b)〕。
2.3 物種多樣性變化
不同修復年限的沙地草本植物多樣性指數無顯著差異(P>0.05)(圖5)。草本植物的4 個多樣性指數均在11 a 為最小值。Shannon-Wiener指數在9 a 為1.11,隨后在11 a 降至最小(0.570)。與9 a 相比,13、15 a 分別高出0.14% 和0.710%〔圖5(a)〕。Simpson指數在9 a 時最大(0.570),在11 a 降為最小(0.300),13 a 與15 a 基本穩定在0.570 左右〔圖5(b)〕。Margalef 指數在9 a時為0.910,在11 a 時降至最小(0.500),13 a 時升至最大(0.930),15 a 時為0.860,比13 a降低了8.27%〔圖5(c)〕。Pielou指數與Margalef指數的變化趨勢相似,在9 a 時為0.710,11 a 時降至最小(0.440),13 a 時升至最大(0.760),15 a 為0.690,比13 a 降低了9.89%〔圖5(d)〕。
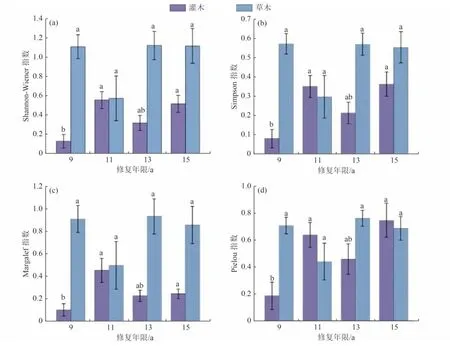
圖5 甘珠爾沙地不同修復年限植被多樣性指數比較Fig.5 Comparison of diversity index of different restoration years in Ganzhuer sandy land
灌木層的多樣性指數隨修復年限的增加表現出的差異不同(圖5),4 個多樣性指數在9 a 與11、15 a均呈現出顯著差異(P<0.05);而13 a 與其他修復年限差異均不顯著(P>0.05)。整體上看,各多樣性指數變化趨勢一致,即上升—下降—上升。Shannon-Wiener指數在9 a 為最小(0.130),11 a 顯著增加了3.34 倍后達到最大(0.560)(P<0.05)。而13 a 降至0.320,在15 a 有所回升(0.520)。修復年限最長(15 a)與修復年限最短(9 a)相比,顯著提高了3.03 倍(P<0.05)〔圖5(a)〕。Simpson 指數9 a 為最小(0.080 0),15 a 達到峰值0.360 且顯著高于9 a(P<0.05)〔圖5(b)〕。Margalef 指數在9 a 為最小(0.100),顯著低于11 a 的最大值0.450(P< 0.05)。11、13、15 a 與9 a 相比,均提高了1.24 倍以上〔圖5(c)〕。Pielou 指數在9 a 為最小(0.190),但隨著修復年限的增加出現明顯升高,11 和15 a 分別提高了2.44 倍和3.03 倍,差異達到顯著水平(P<0.05)。13 a 與9 a 相比提高了1.48 倍,但差異不顯著(P>0.05)〔圖5(d)〕。
2.4 草本多樣性與環境因子的關系
為直觀表達環境因子與植被修復之間的相互關系,找出沙地生態系統修復過程中影響植物多樣性變化的潛在驅動因素,并量化環境因子對植被多樣性的貢獻程度。本研究選用隨機森林模型(RF)方法,將選取的環境因子土壤酸堿度(pH)、電導(EC)、含水量(SMC)、有機質含量(SOC)、總氮含量(TN)、總磷含量(TP)、總鉀含量(TK)、年降水量(AP)、年蒸散量(AE)、年均溫度(AAT)、年均風速(MAWS)、容重(UW))作為解釋變量,草本植物多樣性指數作為響應變量,進行篩選及模型回歸。分析確定包含土壤養分因子與環境因子在內的相對重要性排序圖(圖6)。隨機森林模型的結果顯示,并非所有環境因子對多樣性指數做出了相似的貢獻。其中土壤總氮含量、總鉀含量、有機質含量、年降水量和年均風速為影響沙地植被Shannon-Wiener 和Simpson 指數的關鍵性因子(P<0.01) 〔圖6(a)、(b)〕。同時對于Shannon-Wiener 指數來說,pH 與土壤含水量也對其具有較高的解釋度(P<0.01) 〔圖6(b)〕。土壤總氮含量、有機質含量、土壤含水量和年降水量依次為顯著影響Margalef 指數的環境因子(P<0.01)〔圖6(c)〕。盡管環境因子對Pielou 指數影響均不存在顯著性〔圖6(d)〕,但總鉀含量、土壤含水量、有機質含量與總氮含量仍為影響Pielou 指數的重要指標。部分依賴圖(圖7)結果表明,土壤總氮含量、總鉀含量、有機質含量,土壤含水量及年降水量等環境因子與植被多樣性指數呈顯著正相關,且隨著環境因子的提高而逐漸增加,后穩定在較高的水平。年均風速與植物多樣性呈顯著負相關,當風速超過2.27 m/s 時,沙地生境Shannon-Wiener 和Simpson 指數將會出現顯著下降。
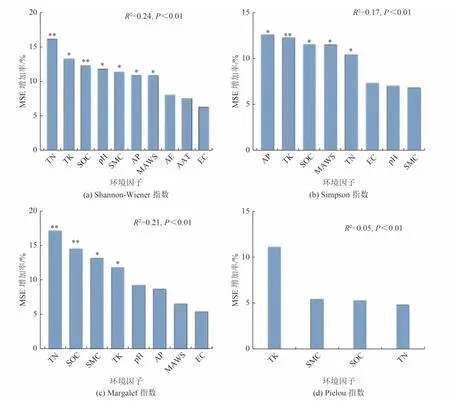
圖6 環境因子與草本植物多樣性指數隨機森林模型重要性排序Fig.6 Importance ranking chart of random forest model of environmental factors and herb diversity index
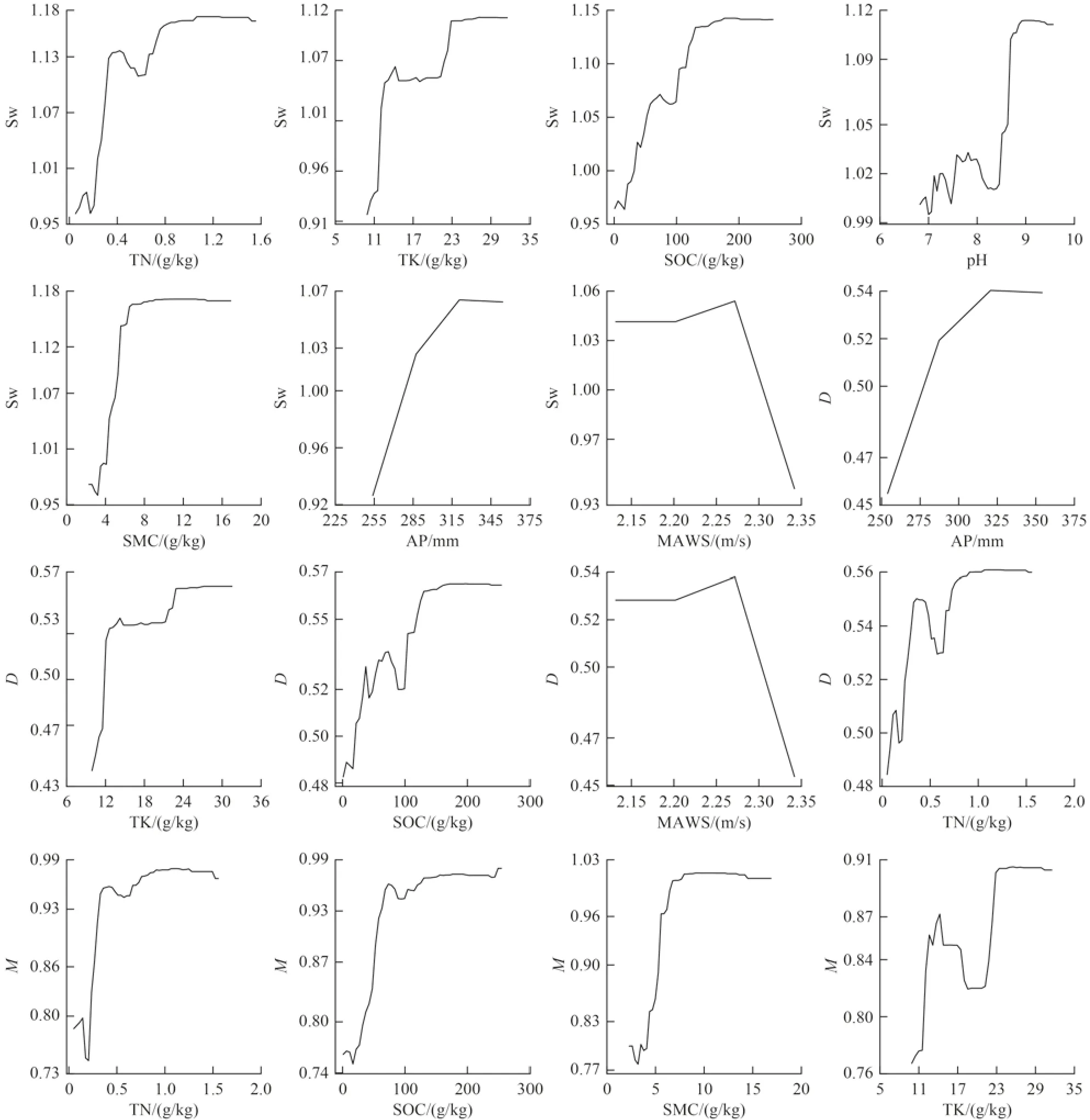
圖7 環境因子與草本植物多樣性指數部分依賴圖Fig.7 Partial dependence of environmental factors on herb diversity index
3 討論
3.1 植物群落特征及優勢度變化
群落的結構特征是研究生態系統的動態變化規律及其維持機制的基礎。群落物種組成對群落動態變化規律以及植被恢復策略等方面的研究具有重要作用[25]。本研究中,不同修復年限的沙地植物群落的組成存在一定差異,草本植物種數呈現先減少后增加的趨勢,這與一些學者在修復跡地的研究結果相似[26-27]。研究發現,草本群落主要集中在菊科和禾本兩大科,其中對草地沙漠化環境具有一定忍耐性的菊科植物物種數最大,這與草地植被退化的逆向演替趨勢恰恰相反[28]。以披堿草、狗尾草、糙隱子草和羊草為主的禾本科植物的數量較穩定,有較高的優勢度,且隨著修復年限的增加逐漸突顯其在群落中的作用和功能。從生活型組成來看,在各修復年限的草本層組成中多年生草本占優勢,且隨著植被修復的進行,抗逆性強的多年生植物種類增加,并且優勢度明顯超過一年生及一/二年生植物,對群落生態功能的維持起主要作用。灌木群落中的優勢科豆科的優勢度及多年生草本優勢度隨著修復過程逐漸上升,這與張繼義等[29]的研究結果相同。多年生植物具有比一年生植物更強的抵抗環境擾動和保持種群穩定的能力,因此物種生活型組成上的這種變化也反映了植被修復過程生態系統結構與功能的變化特征,群落結構趨于穩定,生態功能增強。群落總蓋度和生物量從側面反映了植物光合能力強弱及繁茂程度,是草地植物重要的數量特征。植被群落總蓋度的增加能夠顯著減少水土流失和改善地表小氣候,是植被修復的關鍵性指標。趙嘯龍等[30]在科爾沁沙地自然修復過程中發現修復20 a 的草本植物群落總蓋度和生物量顯著大于修復12 與17 a。而本研究并未發現9~15 a 草本植物群落總蓋度和生物量存在顯著差異,與趙嘯龍等[30]的研究結果并不一致,推測與修復地位置、修復年限、修復措施以及灌草植物的相互作用有關。
本研究發現,在9 a 時灌木群落保持著較低的群落總蓋度與生物量,根據實地調查也發現該年限樣地中灌木植株相較其他年限的灌木更矮小,根系相對不發達,所以根際對養分的截留能力較弱。根際是土壤水分和礦物質進入根系參與生物循環的門戶[31],在半干旱荒漠區,土壤養分含量較低,因此根際微域內的有效養分決定著植物養分實際吸收的養分量,其截留效應的影響相比于農田和森林更明顯[32]。由于受到養分可獲得性的影響,該年限樣地中草本與灌木植物的群落總蓋度和生物量均處于較低水平。修復11 與9 a 相比,草本植物物種數出現降低,為恢復年限中的最小值。同年的樣地中鹽蒿物種成為最優勢種,灌木總蓋度大幅度提高,生物量也出現顯著增加。研究表明,鹽蒿同小葉錦雞兒相比,具有更強的抗旱能力及適應性[33],因此鹽蒿物種的優勢度變化也反映了該修復階段可能受到極端天氣的影響使土壤中養分含量波動劇烈,使競爭能力較強的鹽蒿物種脫穎而出。13 a 樣地中灌木群落維持著較高的群落總蓋度與生物量,且保持著持續穩定增長的趨勢,其灌叢下分布的草本植物豐富度也有顯著增加。較多的草本植物的出現可能與土壤中的養分富集有關。灌木群落在13 a 呈現出以豆科的小葉錦雞兒為優勢種,該旱生灌木的根瘤固氮作用可為植株生長提供氮元素,在缺水、貧瘠、沒有人工干預的沙化草地上具有強烈的適應性[34],因此更多草本植物出現、定居。本研究發現15 a 樣地中無鹽蒿物種分布,調查時僅有小葉錦雞兒與羊柴2 種豆科植物。考慮到鹽蒿是一種好氧植物,適宜在修復年限早期較疏松的半固定沙地上生長,優勢的鹽蒿種群很快使土壤堅實化并向淡栗鈣土化發展,使土壤通氣性降低,造成生境條件轉變從而抑制了自身生長[35]。同年的樣地中由于缺少強抗旱的鹽蒿作為競爭對手,小葉錦雞兒與羊柴灌木保持較高的群落總蓋度和地上生物量且隨修復年限表現出顯著性上升,其生態優勢度逐漸突顯,且朝著高大植物的趨勢發展。在整個修復年限中,物種數、群落總蓋度與地上生物量表現出波動增加,這與其他地區植被修復過程中變化規律一致[36-37]。
3.2 物種多樣性變化
植物多樣性是維持草地生態系統發展和生產力的最關鍵因素之一[38]。時間梯度上植物群落物種多樣性的變化,在一定程度上可以反映出植被的演替變化[39]。本研究中,草本植物各項多樣性指數隨著修復時間的增加整體表現出增加的趨勢,具體為先下降后上升的不顯著波動。這種變化可以一定程度上反映出草本植物群落逐漸達到一個相對穩定的階段,物種豐富程度和復雜程度相似,沙地生態系統保持著穩定的生態效應。草本植物Shannon-Wiener 指數、Simpson 指數、Margalef 指數與Pielou 指數在11 a均低于其他修復年限,反映出11 a 草本植物群落中物種數目少,群落中各物種分布較均勻,最優勢種地梢瓜與披堿草的優勢地位不突出,推測可能由于灌木群落在11 a 表現出較高的優勢度與豐富度,其復雜的群落結構導致其下草本群落結構傾向于簡單化發展。隨著灌木群落郁閉度的持續增加以及其根系越來越發達,所吸收的養分隨之增加,草本植物物種受到光資源與灌木競爭資源空間能力的逐漸減弱,同時優勢灌木物種占據的生態位也對下層草本植物的生長起到一定限制作用[40]。本研究中15 a 草本植物與灌木植物的豐富度與多樣性指數與9 a 相比有顯著提高,15 a 樣地中灌木的均勻度較高,這與單桂蓮等[37]認為14 a 是較適宜的封育年限的結果有所不同,可能與不同物種對沙化生態系統的適應能力及程度的差別有關。鹽蒿物種的退出使得沙地僅存在小葉錦雞兒和羊柴2 種固沙能力強的灌木,由于競爭作用讓二者及對土壤水分、養分的吸收處于相對較穩定的狀態。
3.3 物種多樣性與環境因子的關系
草本植物相較于灌木植物更新速度較快,對環境變化的響應更敏感。其多樣性是生物與環境因子長期作用和適應的結果[41],體現了生物資源的豐富性與環境之間的復雜關系[42-43]。自然因素對沙化土地變化的影響是通過土壤、植被、降水、溫度、風速等多方面共同造成的,而植物多樣性[44]和植物物種分布[45]與氣候、土壤養分密切相關,直接影響著生物多樣性格局[46]。土壤是植物生長發育的基質[47],沙地土壤環境的改善直接影響植被演替進程[48-49],其養分的積累是影響植被蓋度、植物群落組成和植物多樣性的重要因素[50-56]。喀斯特地區[57]、黃山丘陵[58]和毛烏素沙地[59]等土壤養分匱乏的地區,全氮、有機質等土壤養分成為群落分布的決定性因子;在干旱地區水分也是植被恢復的限制因子,土壤含水量為調控森林-草原連續體及凍原生態系統的物種多樣性的主要因子[60-61];pH 是濕地和半干旱草原植被分布的調控因子[62-63];全氮、pH 和全鉀是綠洲植被結構的主要影響因子[64];有機質、全氮、土壤質地和濕度是荒漠生態系統植被分布的制約因子[65];全鹽量、Cl-、K+、Na+、Mg2+等的含量是綠洲荒漠過渡帶群落多樣性的主要影響因子[66]。此外,對于同一的生態系統不同研究區而言影響植被群落分布與多樣性的影響因子也有所不同。
通過對影響沙地修復區草本植物多樣性的關鍵因子的研究,發現人工植被修復9~15 a 的沙地生態系統中,土壤總氮含量、總鉀含量、有機質含量、年降水量、土壤含水量及年均風速等環境因子與植物多樣性顯著相關,其中土壤養分(如土壤總氮含量、總鉀含量和有機質含量)與該地區的物種多樣性顯著正相關,這一結果與以往研究中發現由于土壤養分含量低導致植物物種多樣性下降的結果相似[67-68],另外推測與半干旱地區典型的灌叢“肥島”現象有關[69-71],灌叢肥島的發育有利于根系養分的利用,產生較多的根際沉積,而根系的活動又會促進肥島的發育[32],進一步加劇土壤養分脅迫。除土壤養分的影響,本研究還表明風速和降水成為制約植物的主要因素,其年際間的變化對物種多樣性也有顯著影響。對于荒漠程度下的沙化草地,年均風速的增加與植物多樣性的顯著負相關關系,表明更高的風速帶來土壤表面更大的風蝕,使植物多樣性降低。這與基于全球環境變化的長期監測研究發現風速、溫度與降水影響著多年生物種多樣性變化的結論一致[72]。與土壤養分因子相比,本研究中甘珠爾修復區的植被受到水分的影響表現出弱于土壤總氮含量、總鉀含量和有機質含量等因子。推測其原因為研究區曾分布有舊河道,因此地下水對土壤仍存在一定的補給作用。另外,在沙質草地多為草本植物,密布的根系層對降水存在滯留作用[73],草本植物相比于灌木耗水相對較少,且較高的植被蓋度產生的地表遮蔭效果也能有效減弱土壤蒸發[74],這些因素的綜合作用使其土壤水分含量提高,進而使得水分對植物多樣性的限制作用弱于土壤養分。同時本研究發現,在本研究區中沙地土壤含水量對植被生長存在重要影響,這是因為在固定沙丘時的優勢物種主要為根系較深、耗水強烈的鹽蒿、小葉錦雞兒與楊柴3 種灌木,使得土壤水分消耗強烈,因而導致草本植物受到一定程度的土壤水分的限制。綜上所述,甘珠爾沙地修復區植被多樣性受到氣候、水分、土壤養分特征等因素的共同影響,因此需要對不同修復階段的沙化草地植被群落進行調查與研究,并根據養分需求對其實施合理的修復措施。
4 結論
(1)甘珠爾沙地調查共發現植物42 種,以菊科、禾本科、薔薇科、豆科為主。草本群落在不同修復年限樣地中表現為菊科及禾本植物數量穩定且占有高優勢度,群落總蓋度和生物量變化不顯著,但隨修復年限的增加呈現先增后減再增加的趨勢。物種分布均勻性良好,豐富度指數和多樣性指數在各修復年限間出現小幅波動,但無顯著差異,年際間變化穩定。灌木群落整體呈現出以豆科為優勢種,隨修復年限保持著較高的群落總蓋度和地上生物量。灌木物種組成較單一,但多樣性指數整體上隨著修復年限的延長存在顯著增加的趨勢。
(2)甘珠爾沙地修復區草本植物多樣性主要受土壤養分含量(土壤總氮、總鉀、有機質)和修復當年的自然條件(年降水量、年均風速)影響。在沙化草地植被修復過程中,適當補充土壤養分含量(土壤總氮、總鉀、有機質)和水分,有助于干旱區域荒漠草原生態系統的群落結構的重建和生物多樣性的恢復。
(3)未來應持續開展沙地修復后的植被生態調查,掌握修復后生態系統群落結構及生物多樣性情況,分析生態系統修復的影響因素,以了解沙化地區植被修復成效,為沙化區域植被修復策略的制定提供科學參考。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
