基于生物信息學分析MicroRNA-199靶基因的預測及在肺發育中的作用
2023-08-03 14:12:50喻玢黃棟
貴州醫藥 2023年7期
喻玢 黃棟
(1.貴州醫科大學,貴州 貴陽 550004;2.貴州省人民醫院,貴州 貴陽 550001)
目前利用干細胞移植來促使組織修復的治療手段已成為前沿科學,羊水干細胞因其取材來源為分娩廢棄物,倫理束縛較小,來源廣泛,取材較為容易,故在前臨床研究中越來越受到重視,羊水是胎兒生長的保護液,在胚胎發生期間提供機械支持和必要的營養。它主要由水、細胞和化學元素構成[1],人羊水干細胞(human amniotic fluid stem cells,hAFS)做為羊水中的提取物已被廣泛研究。hAFS已被證明在體外和體內具有廣泛的多能性[2],有助于激活內源性修復反應,同時還具有免疫調節功能等。目前已有研究表明人羊水干細胞(hAFS,human amniotic fluid stem cells)成梭形細胞形態[3],其細胞表達間充質干細胞(Mesenchymal stem cells,MSCs)的表面標志物如CD90,CD105,而不表達造血干細胞的表面標志物CD34、CD45及人類白細胞抗原基因HLA-dr[4],以上細胞學特點均提示人羊水干細胞為間充質干細胞。間充質干細胞的組織修復作用主要依賴于營養因子、抗炎蛋白和外泌體的旁分泌途徑[5],外泌體含有多種細胞因子、生長因子、代謝產物以及間充質干細胞自身產生的microRNA(miRNAs),當外泌體與相應靶細胞膜融合后,外泌體內容物釋放,以microRNA(miRNAs)對受體細胞進行信號調節。目前已有學者證明hAFS對各類肺發育不良模型中促進了胎兒肺再生[6],實驗中證明在支氣管肺發育不良模型中,氣管內給予AFS后可促進肺泡發育,減弱了血管重塑和肺動脈高壓,降低了肺促炎細胞因子表達,減少了巨噬細胞浸潤。最新研究顯示,microRNAs(miRNAs)在多種生物過程中扮演了重要角色,如細胞的增殖、凋亡、分化等[7],可以通過直接降解靶基因或者抑制其翻譯來沉默靶基因,參與大多數細胞機制的調節功能,目前miRNAs在細胞分化和器官再生中的功能逐漸凸顯,已經有多個miRNA被證明在肺發育中發揮了重要作用。發掘并研究新的miRNAs在肺發育中的作用對開拓治療肺疾患、促進肺組織再生及修復有著重要意義。
1 資料與方法
1.1差異性miRNA分析 在對hAFS的研究,通過對圍產期hAFS(perinatal human amniotic fluid stem cells,p-hAFS)和胎兒期hAFS(fetal human amniotic fluid stem cells,f-hAFS)miRNAs的差異性比較研究(常規妊娠Ⅱ期獲得的羊水干細胞為胚胎期,從妊娠Ⅲ期分離出來的羊水干細胞為圍產期),通過對p-hAFS和f-hAFS的差異miRNA進行生物信息學分析從數據集GSE168152獲取p-hAFS(GSM5129985,GSM5129987,GSM5129991,GSM5129994,GSM5129983)和f-hAFS(GSM5129977,GSM5129979,GSM5129981,GSM5129989)的miRNA測序數據,使用DESeq包比較了5個p-hAFS樣本和4個f-hAFS樣本之間的差異miRNA(篩選條件為:P<0.05,logFC>1)。
1.2靶基因預測及功能富集分析 通過主流的miRNA-mRNA互作數據庫對3個目標miRNA(hsa-miR-199b-5p,hsa-miR-199b-3p,hsa-miR-199a-3p)進行預測分析,這里我們主要選取了miRDB,miRWalk,Starbase進行預測分析;理想狀況下我們取3個數據集的交集作為miRNA的靶基因。但經過這樣的處理后hsa-miR-199b-5p 在KEGG富集中只能找到2條通路是顯著的。因此我們對取交集的數據集進行了變更,經過觀察發現,miRDB數據庫預測得到的靶基因數目最少,因為我們決定只取miRWalk和Starbase數據庫預測的結果。通過上述方式在miRWalk和 Starbase數據庫中共預測得到291個hsa-miR-199a-3p的交集靶基因;同時預測得到363個hsa-miR-199b-5p的交集靶基因。為了說明差異表達基因參與的生物學功能和信號通路,利用R/Bioconductor package clusterProfiler、org.Mm.eg.db、enrichplot、ggplot2包對差異表達基因進行GO和KEGG富集分析。
2 結 果
2.1篩選差異表達的miRNA 通過對p-hAFS和f-hAFS的差異miRNA進行生物信息學分析出p-hAFS及f-hAFS中差異表達的16個基因,同時注意到在p-hAFS中高表達的6個miRNA即miRNAhsa-miR-3919,hsa-miR-199a-3p,hsa-miR-199b-3p,hsa-miR-199b-5p,hsa-miR-875-3p,hsa-miR-424-5p中有3個基因來自同一族系,即hsa-miR-199a-3p,hsa-miR-199b-3p,hsa-miR-199b-5p。使用heatmap繪制熱圖(圖1)來展示差異表達情況。基于熱圖可以看到,在p-hAFS中hsa-miR-199a-3p,hsa-miR-199b-3p,hsa-miR-199b-5p均為hsa-miR-199族系miRNA,且在p-hAFS中的表達顯著,在一篇胎兒和圍產期人羊水干細胞的分泌組配方的全面概況[8]中亦檢測到p-hAFS中的hsa-miR-199b-3p的序列。差異表達的micRNA可能在28周后胎肺加速發育中發揮重要作用,繼而為肺發育的相關研究提供新的思路。在目前對miR-199基因的前臨床研究中可以了解到miR-199可通過靶向SP1的表達負性調控成纖維細胞增殖和纖維化[9],miR-199a/b-3p通過下調PAK4/MEK/ERK通路可抑制胃癌細胞MGC-803的增殖[10],miR-199a-3p可抑制mTOR的表達引起細胞自噬從而促進細胞凋亡降低細胞增殖[11],但與肺發育的相關性尚不明確。通過生物信息學分析對其靶基因及靶基因功能進行富集分析,為深入研究hsa-miR-199a-3p,hsa-miR-199b-3p,hsa-miR-199b-5p在肺發育中的生物功能及調控機制提供理論指導。
2.2靶基因預測及miRNA靶基因功能富集分析 我們分別使用miRDB,miRWalk,Starbase等3個數據庫進行對hsa-miR-199b-5p,hsa-miR-199b-3p,hsa-miR-199a-3p預測可能結合的靶基因。研究結果顯示hsa-miR-199a-3p和hsa-miR-199b-3p在數據庫中預測的結果一致,于是我們選取hsa-miR-199a-3p的預測靶基因結果。每個miRNA的靶基因取交集處理,在數據庫中共預測得到291個交集靶基因。hsa-miR-199b-5p的miRDB數據庫預測得到的靶基因數目較少,因此只選取miRWalk和Starbase數據庫預測的結果,共預測到363個交集靶基因。基于上述結果,我們對靶基因進行功能富集分析,利用R包clusterprofile進行通路富集分析。在hsa-miR-199a-3p相關靶基因的KEGG富集分析中,我們共鑒定出37條富集通路,在GO中共鑒定出505通路被顯著富集,其中包括371條BP,56條CC,78條MF。在KEGG富集結果中,顯示前30個KEGG通路(圖2)。對GO通路進行富集分析,如圖為最顯著差異的前30個GO通路(圖3)。在hsa-miR-199b-5p相關靶基因的KEGG富集分析中,我們共鑒定出21條通路被顯著富集,在GO中共鑒定出370通路被顯著富集,其中包括268條BP,40條CC,62條MF。在KEGG富集結果中,我們展示了21條顯著的KEGG通路。(圖4)。hsa-miR-199b-5p相關靶基因的GO通路展示差異最顯著的前30個富集分析結果(圖5)。
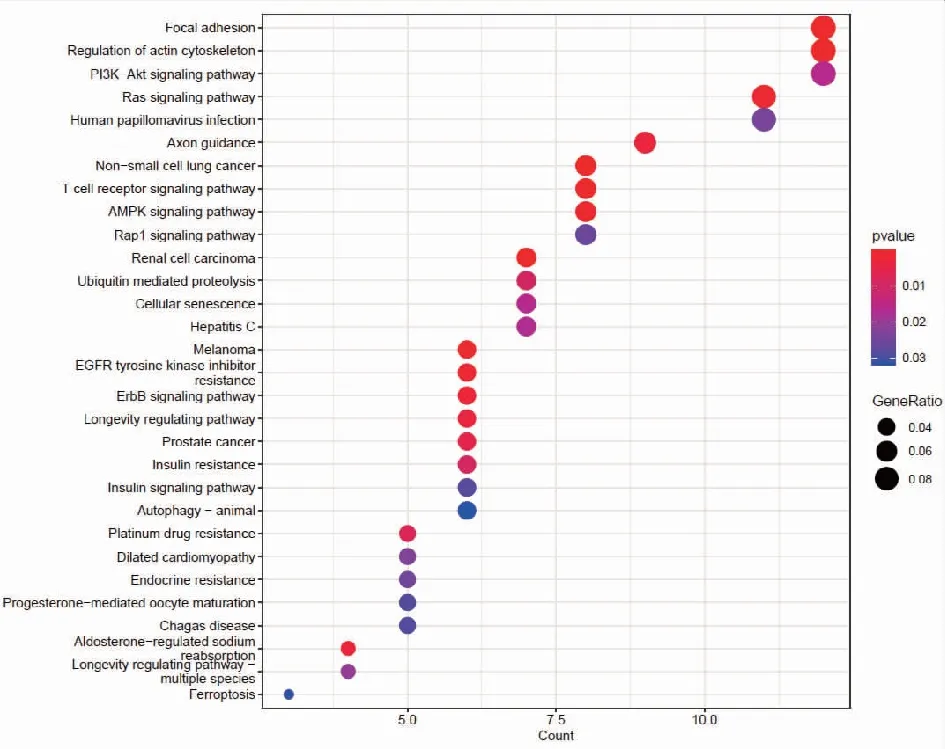
注:富集得到最顯著的前30個KEGG條目,橫軸表示 KEGG通路的基因數目,縱軸表示KEGG通路的名稱,顏色表示 Pvalue。
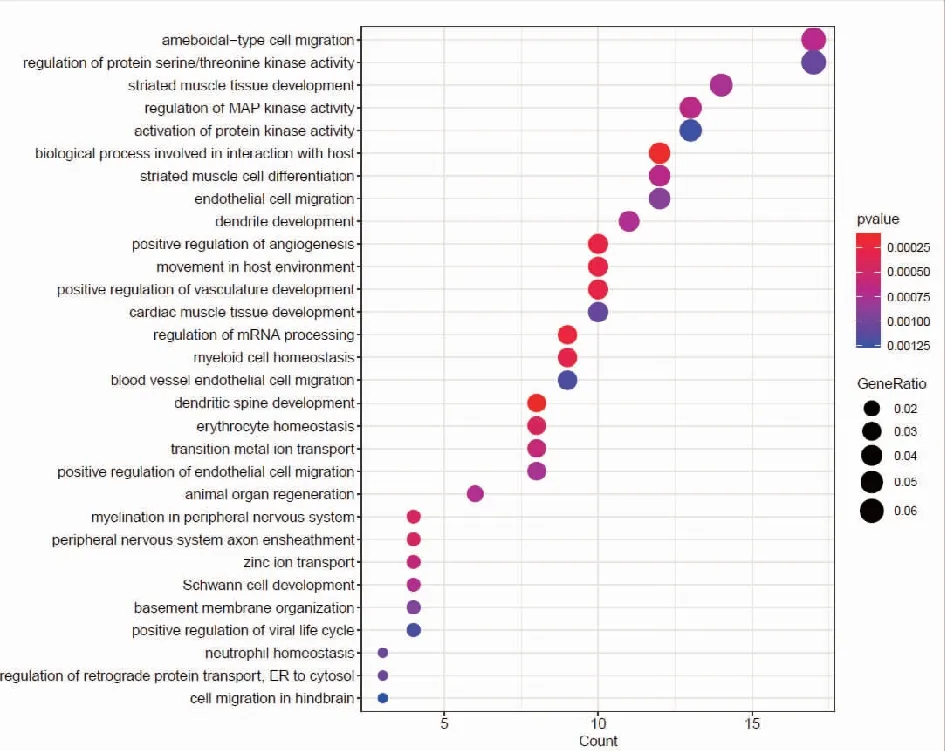
注:富集得到最顯著的前30個GO條目,橫軸表示 GO條目的基因數目,縱軸表示GO條目的名稱,顏色表示 Pvalue。
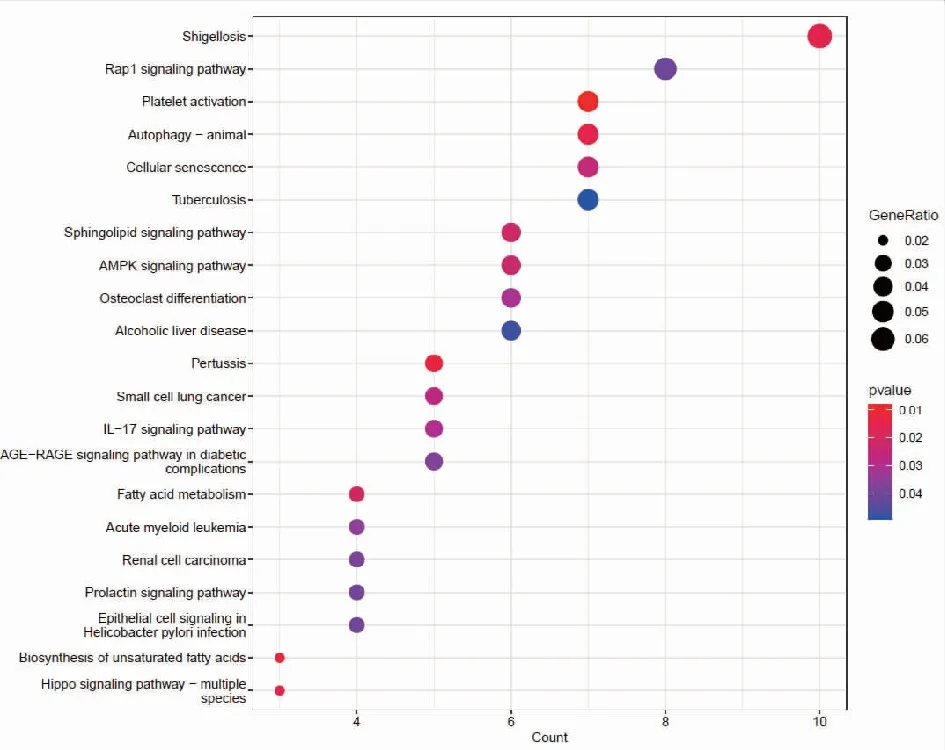
注:富集得到最顯著的前21個KEGG條目,橫軸表示 KEGG通路的基因數目,縱軸表示KEGG通路的名稱,顏色表示 Pvalue。
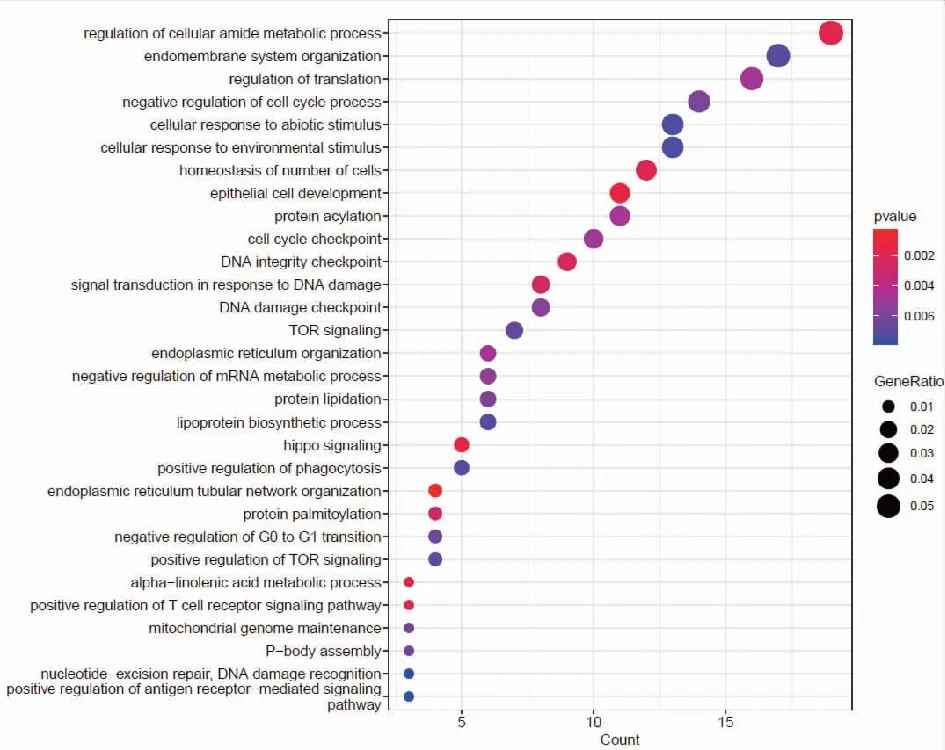
注:富集得到最顯著的前30個GO條目,橫軸表示 GO條目的基因數目,縱軸表示GO條目的名稱,顏色表示 Pvalue。
3 討 論
肺發育是一個復雜的過程,受信號分子網絡的調控。特別是一些miRNAs,miRNA作為一類基因轉錄后調控其表達水平的非編碼RNA,可以通過直接促進mRNA的降解或干擾mRNA翻譯來沉默靶基因,發揮其生物學作用[12],在肺發育過程中miRNA可能通過控制分支形態發生和上皮和間質分化[13],糾正信號分子失調的網絡來促進肺發育。目前研究發現hAFS能夠釋放出包含microRNA的囊泡,能夠抑制炎癥,提供心肌保護,減輕心肌毒性,刺激血管生成等作用[14-16]。使用miRDB,miRWalk,Starbas數據庫對hsa-miR-199b-5p、hsa-miR-199b-3p、hsa-miR-199a-3p進行靶基因預測,因hsa-miR-199a-3p與hsa-miR-199b-3p預測的靶基因一致,故合并討論。
細胞自噬是一種進化上的保守過程,細胞內物質被雙膜自噬體隔離,然后在各種生理病理條件下被溶酶體降解和回收。自噬作用本身不能促進肺組織的發育,但當肺損傷發生時,自噬可以減輕肺纖維化的形成,W.Zheng等[17]實驗發現自噬與損傷后的纖維變性呈負相關。實驗結果提示hsa-miR-199a-3p及hsa-miR-199b-5p均富集到AMPK信號通路,且相關性較高,AMPK激活細胞自噬作用上調可誘導細胞死亡或啟動調亡程序促進肺損傷病理進展。除此以外hsa-miR-199a-3p還富集到兩條與自噬相關的信號通路,PI3K信號通路及RAS信號通路。PI3K通路可通過激活成熟的PI3K-AKT-mTOR (雷帕霉素的機械靶點)復合物1(MTORC1)通路抑制自噬[18],且Ⅰ類PI3K的PIK3CB催化亞基可作為自噬的正向調節因子。RAS信號通路可直接激活自噬[19]。hsa-miR-199a-3p及hsa-miR-199b-5p很可能通過細胞自噬在肺組織損傷發生時產生積極作用。
除AMPK信號通路外,在hsa-miR-199a-3p及hsa-miR-199b-5p上同時富集到的還有T細胞受體調節信號通路。而T細胞受體調節信號主要與細胞內的炎癥反應調節相關,與自噬相同,炎癥反應的調節本身不具有促進肺組織發育的功能,但對肺損傷尤其是支氣管肺發育不良等疾病具有重要影響[19]。hsa-miR-199a-3p的富集結果中ErbB信號通路[20]及hsa-miR-199b-5p富集到的IL-7信號通路同樣具有炎癥調節的作用。炎癥調節作用可在肺組織出現感染等損傷時,下調其炎性物質的表達從而減輕肺組織的損傷。
血管的發育在胎肺發展的過程中有著重要作用,而血管重塑在發生肺損傷時也是重要的病理過程。在hsa-miR-199a-3p的GO富集分析中,與hsa-miR-199a-3p最顯著差異的前30個GO通路中共有兩個與血管調節相關的通路,血管發育的正向調節、血管生成的正性調節),而此機制可能是hsa-miR-199a-3p影響肺發育的機制之一。
在現有的前臨床實驗中,已驗證P53可通過調節miR-199a-3p來影響體細胞編程。而在我們對hsa-miR-199a-3p的富集結果中,我們富集到miR-199a-3p在動物器官再生、mRNA加工的調控、體內平衡等與組織發育、細胞編程相關的通路,而mRNA加工調控與樹突棘發育通路間有共同基因參與。Hsa-miR-199a-3p與細胞的發育有著密切關系。而在hsa-miR-199b-5p的GO富集分析中,細胞酰胺代謝過程的調節、轉化的調節、mRNA代謝過程的負調控、細胞對環境刺激的反應、細胞數量的穩態、蛋白質酰化、細胞周期檢查點等生物活性通路;其富集到的可參與細胞編程的通路數量更多,且在這些通路中有多個基因重復出現,目前已有研究[21]表明miR-199b可抑制VEGF、JAG1、SET蛋白的表達,為其靶基因,可促進細胞的增殖,延長細胞周期,并抑制細胞凋亡,而靶基因的富集結果與之相符,由此推測hsa-miR-199b-5p可能直接參與到了細胞對環境的應激甚至mRNA的表達調節中,對于細胞活性及細胞反應有直接作用,上述通路可能對肺發育的進程產生作用。
綜上所述,hsa-miR-199b-5p、hsa-miR-199b-3P、hsa-miR-199a-3p是圍產期羊水干細胞中對肺發育起作用的因子,對肺發育的調控可能是多維度的。hsa-miR-199a-3p及hsa-miR-199b-5p的靶基因具有豐富的生物功能,對靶基因富集到的信號通路進行進一步分析得出以下結論:①hsa-miR-199a-3p及hsa-miR-199b-5p可能通過AMPK信號通路、動物自噬、P13k-Akt信號通路等通路上調細胞的自噬作用;②通過血管發育的正向調節、血管生成的正向調節促進肺發育中肺血管的生成或減輕肺損傷中肺血管重塑;③T細胞受體信號通路、ErbB信號通路及hIL-7信號通路通過對炎性介質的調節在肺損傷的過程中起到積極作用;④hsa-miR-199b-5p、hsa-miR-199b-3p、hsa-miR-199a-3p還具有mRNA加工的調控、DNA損傷檢查點、DNA損傷的信號轉導反應等具有調控作用,對肺發育可能是有益的。
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
中國生殖健康(2019年3期)2019-02-01 06:12:26
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
海軍航空大學學報(2015年3期)2015-11-11 17:20:00
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
終身教育研究(2014年5期)2014-02-28 01:23:06
