生物炭和腐殖酸施用對稻麥輪作系統CH4 和N2O 綜合溫室效應的影響
2023-08-15 16:19:04劉志偉吳家森李永夫姜培坤
農業工程學報 2023年11期
劉志偉 ,劉 娟 ,吳家森 ,李永夫 ,姜培坤
(1. 浙江農林大學環境與資源學院、碳中和學院,亞熱帶森林培育國家重點實驗室,臨安 311300;2. 浙江農林大學環境與資源學院、碳中和學院,浙江省森林生態系統碳循環與固碳減排重點實驗室,臨安 311300)
0 引 言
大氣中CO2、CH4和N2O 等溫室氣體濃度增加而導致的全球變暖成為世界性的環境問題,其中N2O 和CH4的增溫潛勢分別是CO2的298 倍和25 倍左右[1]。中國在聯合國大會上提出中國CO2排放量力爭在2030 年達到峰值,在2060 年達成“碳中和”的目標[2]。農業生產是生態系統中CH4和N2O 的重要來源之一,分別占全球人為排放CH4和N2O 的50%和43%以上[3]。稻麥輪作制度是中國東南部典型的種植制度之一。據估計,稻麥輪作占中國糧食生產總面積的8.2%,其中糧食產量和溫室氣體排放量分別占9.8%和15.0%[4]。如何通過減排增匯實現“碳中和、碳達峰”已經成為當前氣候變化研究的熱點。
自然土壤的酸化伴隨在土壤發生和發育的整個過程,近些年來大量的人為活動進一步加速了土壤酸化的進程[5]。由土壤酸化導致農業土壤退化的問題日益嚴重,中國主要的農作物產區的土壤pH 值在1980—2000 年間平均下降了0.5 個單位[6]。生物炭是秸稈等生物質在厭氧或無氧條件下經過高溫熱解產生的一種富含碳的固態產物[7]。研究表明,生物炭的施用能提高土壤碳、氮含量,改善土壤微生物群落結構,提高農作物產量[8]。研究發現生物炭添加能夠抑制土壤N2O 和CH4排放[9],但作為一種難降解富含碳的物質,生物炭在土壤中可穩定地存在數百甚至千年[10],且對農田溫室氣體排放的影響會隨施用時間而發生改變[11-12]。朱爽閣等[11]通過田間定位試驗,發現陳化生物炭能降低土壤N2O 排放,其抑制作用隨陳化時間延長而減弱。WU 等[12]通過6a 田間試驗,發現生物炭施入初期對土壤CH4排放沒有顯著影響,但是試驗后期,陳化生物炭顯著降低了土壤 CH4的排放。稻麥輪作系統包括灌溉水稻和旱地小麥,土壤環境不同會導致施用生物炭的影響也不一致[13-14]。ZHOU 等[13]在稻麥輪作的研究中發現,施加生物炭能夠顯著降低麥季N2O 排放量,但對稻季的N2O 排放無顯著影響。ZHANG等[14]在太湖地區的稻麥輪作中研究發現,生物炭在20和40 t/hm2施用條件下對麥季GWP 和GHGI 均有顯著降低作用,而稻季則無顯著影響。因此在稻麥輪作這種復雜的環境中陳化生物炭對溫室氣體排放的影響有待進一步研究。
腐殖酸是動植物遺骸(主要是植物的遺骸)經過微生物的分解和轉化以及地球化學的一系列過程形成和積累起來的一類大分子有機物質[15]。腐殖酸具有較大的表面積和內部電荷,可以作為土壤改良劑改善土壤理化性質,提高土壤pH 值和有機碳含量,緩解土壤酸化,提升土壤肥力,提高作物產量[16-18]。腐殖酸作為土壤改良劑對農業溫室氣體排放的研究較少,SUN 等[19]研究發現,土壤中腐殖酸的添加促進了N2O 排放,對CH4排放產生顯著影響。
本文以浙江稻麥輪作系統為研究對象,研究生物炭與腐殖酸單獨施用與配合施用對CH4和N2O 排放影響;同時結合作物產量評估該生態系統綜合溫室效應和溫室氣體強度,旨在綜合評價施用生物炭和腐殖酸對稻麥輪作系統溫室氣體排放的長期效應。
1 材料與方法
1.1 試驗區概況
試驗地點為浙江省杭州市臨安區於潛鎮(30°25’ N,119°45’ E),地處浙江省西北部,屬于亞熱帶季風性氣候。年平均降水量約為1 614 mm,降雨大多發生在4 月份至10 月份期間,年平均降水日158 d,年平均氣溫16.6 ℃,年平均無霜期241 d,年日照時數達1 900 h 以上,土壤類型屬于潛育性水稻土。試驗周期內日降雨量和氣溫的動態變化見圖1。
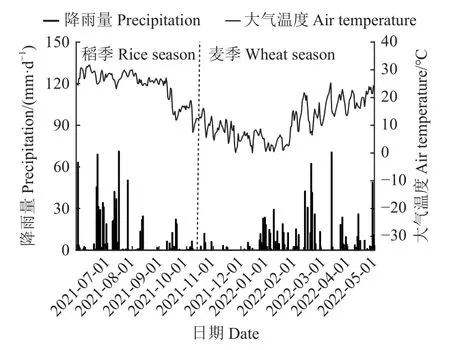
圖1 2021—2022 年水稻-小麥輪作周期內日降雨量和氣溫的動態變化Fig.1 Dynamic changes in daily precipitation and temperature during the rice and wheat annual rotation cycle in 2021-2022
1.2 試驗設計與田間管理
本試驗為雙因素(生物炭和腐殖酸)的田間試驗,各處理的生物炭和腐殖酸添加量見表1。本試驗所用生物炭為江蘇省華豐農業生物工程有限公司提供,以水稻秸稈為原料,采用600 ℃熱解炭化制成。腐殖酸為安徽省無為縣花卉肥料廠提供。生物炭和腐殖酸在施加前均過2 mm 篩處理,其基本理化性質見表2。生物炭和腐殖酸于2020 年7 月11 日一次性施加到土壤0~20 cm 耕作層,手工充分混勻。每個處理3 次重復,試驗共18 個小區,每個試驗小區大小為5 m × 5 m,隨機排列,每個小區設獨立的灌排水口。
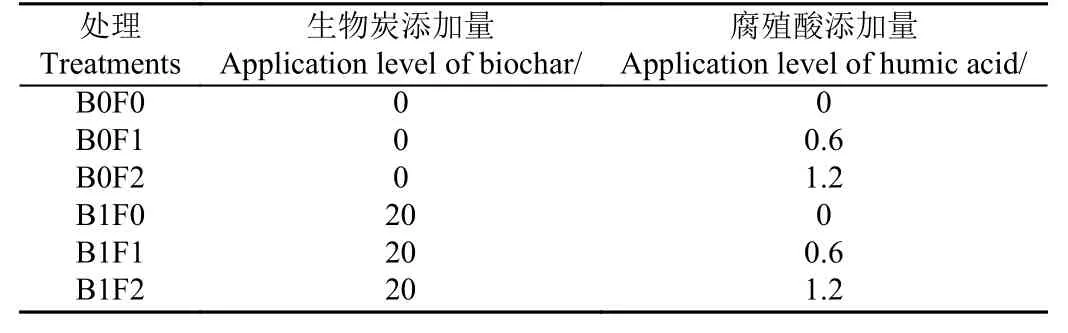
表1 試驗設計Table 1 Field experimental treatments(t·hm-2)
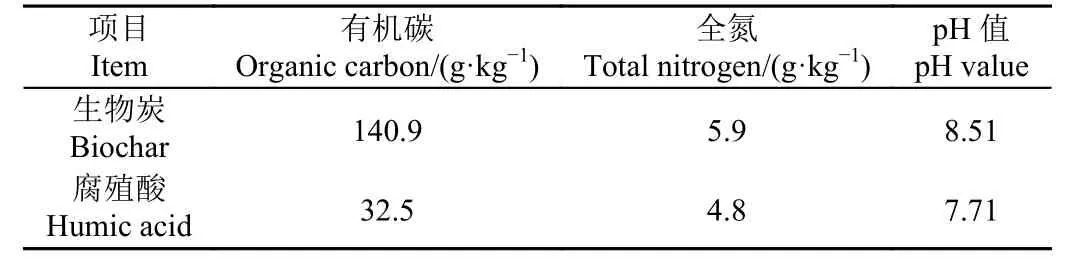
表2 生物炭和腐殖酸基本理化性質Table 2 Basic physical and chemical properties of biochar and humic acid used
自2020 年7 月開始,所有小區在移栽水稻或種植小麥前一天施入化肥作為基肥,稻季氮肥(以N 計)、磷肥(以P2O5計)、鉀肥(以K2O 計)分別為270、75、150 kg/hm2,基肥和分蘗肥的氮肥用量均為135 kg/hm2,基肥采用當地農民常用復合肥(N : P2O5∶K2O 為17∶10∶17),追肥采用尿素和氯化鉀。麥季氮肥(以N計)、磷肥(以P2O5計)、鉀肥(以K2O 計)分別為115、72、38 kg/hm2,基肥和分蘗肥的氮肥用量分別為81 和34 kg/hm2,基肥采用當地農民常用復合肥(N∶P2O5∶K2O 為18∶16∶6),追肥采用尿素和硫酸鉀。所有處理的施肥量、施肥方式、田間病蟲害管理措施同當地農戶習慣常規管理一致。基肥撒施于土壤表層并均勻翻耕,追肥均勻撒施于土壤表層。
本研究觀測期間,水稻于2021 年7 月8 日移栽,11 月11 日收獲,全生育期共 128 d,稻麥輪作田間管理措施依據當地常規進行,水稻生長前期保持淹水,分蘗期后排水曬田,之后恢復灌溉,保持土壤濕潤,后期逐漸排干水分,于2021 年7 月17 日追肥;小麥于2021 年11 月27 日播種,2022 年5 月25 日收獲,全生育期共180 d,追肥時間為2022 年3 月14 日。
1.3 樣品采集與測定
氣體樣品采樣時間為2021 年7 月16 日至2022 年5 月25 日,利用靜態暗箱觀測法采集。水稻和小麥生長期內平均采樣間隔時間為7 和14 d,施肥后適當增加采樣頻率,采樣時間集中在08:00-11:00。采樣箱規格為50 cm×50 cm×50 cm(作物生長前期)或50 cm×50 cm×100 cm(作物生長后期)。采集氣體時,在底座凹槽內加水,以保證頂箱嵌入底座時密封。采樣箱封箱0、10、20 和30 min 后用60 mL 注射器抽取箱內氣體樣品,返回實驗室后立即用氣相色譜儀(Shimadzu, GC-2014,日本)測定。CH4檢測器為FID,溫度為200 ℃,燃氣為氫氣,助燃氣為空氣;N2O 檢測器為ECD,檢測器溫度250 ℃,分離柱溫度55 ℃,載氣為氮氣。
采集氣體的同時,用溫度計記錄箱內溫度變化情況,用地溫計測定地下5 cm 的土壤溫度。采氣后采集0~20 cm 土層土壤樣品對土壤含水量、pH 值,NH4+-N 和NO3--N 含量進行測定。土壤含水量采用烘干法測定,pH 值采用pH 計法測定(水土比 2.5∶1);用靛酚藍比色法(625 nm)和紫外雙波長法(275 和220 nm)測定土壤NH4+-N 和NO3--N 含量。水稻和小麥收獲前,采集測定0~20 cm 土層的土壤基本理化性質:pH 值采用pH 計法測定(水土比 2.5∶1);環刀法測定土壤容重;重鉻酸鉀-硫酸消化法測定有機質;凱氏定氮法測定全氮。水稻和小麥在生理成熟期收獲,并測定作物產量。
1.4 數據處理
CH4和N2O 排放通量根據公式(1)計算。
式中F為CH4或N2O 的排放通量,mg/(m2·h);ρ為標準狀態下氣體流動時的密度,kg/m3;h為采樣箱的高度,m;dc/dt為采樣過程中箱內氣體摩爾分數變化速率,μL/(L·h);T為采樣過程中采樣箱內的平均溫度, ℃。
CH4和 N2O 累積排放量根據公式(2)計算。
式中f為CH4或N2O 的累積排放通量,mg/m2;n為采樣總次數;i為采樣次數,(ti+1-ti)為兩次采樣時間的間隔天數,d。
在100 a 尺度上綜合溫室效應[1](GWP),計算見式(3)。
式中XGWP為綜合溫室效應,以CO2排放量計,kg/hm2。ECH4是CH4累積排放量,kg/hm2;EN2O表示N2O 累積排放量,kg/hm2;25 和298 分別為CH4、N2O 轉換為CO2排放量的系數。
溫室氣體排放強度是單位產量的溫室氣體排放強度(GHGI)計算見式(4)。
式中XGHGI為溫室氣體排放強度,kg/kg;XGWP為綜合溫室效應,kg/hm2;Y為作物產量,kg/hm2。
運用Excel 2013 進行原始數據的計算,利用SPSS 22 進行統計分析,用Origin 2018 軟件作圖。采用雙因素方差法分析生物炭和腐殖酸對土壤基本理化性質、CH4和N2O 累積排放量、水稻產量、綜合溫室效應GWP 和GHGI 的影響,用 LSD 法檢驗不同處理間差異的顯著性,采用Pearson 相關性分析處理CH4和N2O 排放通量與土壤理化性質的相關性。本文中所有圖表中的數據均是3次重復的平均值。除特殊說明外,顯著性水平a=0.05。
2 結果與分析
2.1 土壤理化性質和產量
生物炭一般為堿性,其灰分含有的鈣、鎂、鉀、鈉等鹽基離子可以交換土壤中的H+及交換性Al3+,降低其在土壤中的含量,提高土壤pH 值,可作為改良劑緩解土壤酸化[20]。由表3 可知,生物炭顯著提高了麥季土壤pH 值(P<0.05)。生物炭比表面積大,具有極強的吸附性,能夠吸附土壤中的有機分子,并通過表面催化活性促進有機小分子聚合形成土壤有機質[21]。由表3 可知,生物炭顯著提高了稻麥兩季土壤有機碳含量(P<0.05)。研究表明,腐殖酸中含有大量的有機膠體和有機質,可以作為土壤改良劑或者肥料增加土壤有機質含量,提高土壤養分含量[22]。由表3 可知,腐殖酸顯著提高了稻季土壤有機碳含量(P<0.05)。本試驗中,腐殖酸處理對麥季土壤pH 值、容重、有機碳和全氮含量均無顯著影響,可能是腐殖酸的改良效果持續時間有限,隨著施用時間的增加,腐殖酸對土壤的影響逐漸減弱所致,一般需要持續施用才能達到較好的改良效果,也可能是大田的緩沖作用以及大田環境的多變性所致,具體原因和機制還需后續試驗進行驗證。
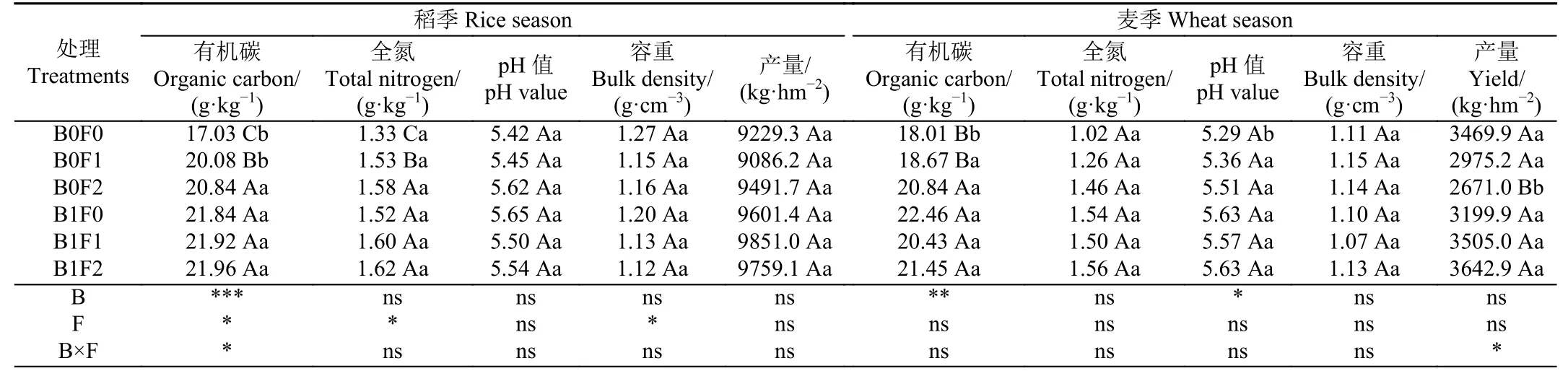
表3 不同處理2021—2022 年稻麥輪作土壤理化性質和產量的變化Table 3 Variation in soil physicochemical properties and yield under different treatments during rice-wheat rotation cycle in 2021-2022
研究表明,生物炭的施用能促進作物根系生長,進而提高作物吸收養分的能力,增加作物產量[23]。研究發現,腐殖酸可以通過改善植物根系,提高作物對土壤養分的吸收利用,促進作物產量的提高[24]。在本試驗中,經過1a 陳化后的生物炭和腐殖酸均未對水稻和小麥產量產生顯著影響(P>0.05),生物炭和腐殖酸配施顯著增加了小麥產量增幅為1.0%~5.0%(P<0.05),對水稻產量沒有顯著影響(表3)。生物炭添加對農作物產量的影響較為復雜,與生物炭類型、試驗地土壤性質、種植作物種類等因素相關,也有研究表明,與小麥等旱地作物相比,生物炭添加對水稻產量的影響有限[25]。
2.2 CH4 排放
CH4和N2O 排放情況如圖2 所示。稻麥輪作期內不同處理稻季CH4排放通量變化趨勢相近,生物炭處理對CH4排放通量無顯著影響(P>0.05),腐殖酸處理顯著降低了CH4排放通量(P<0.05)。B0F2、B1F0、B1F1和B1F2 處理CH4排放通量的峰值均在水稻移栽后第28天出現,分別為42.99、55.82、41.75和43.21 mg/(m2·h),B0F0 和B0F1 的峰值均出現在移栽后第35 天,分別為50.40 和52.13 mg/(m2·h),隨后減少且維持較低排放水平(圖2a)。
水稻生長期間B0F0、B0F1、B0F2、B1F0、B1F1 和B1F2 的CH4平均排放通量分別為:17.96、13.38、14.15、16.50、11.78 和13.18 mg/(m2·h)。生物炭處理對稻季土壤CH4累積排放量沒有顯著影響(P>0.05),腐殖酸處理顯著降低了CH4累積排放量(P<0.05),腐殖酸和生物炭對CH4累積排放量不存在顯著交互作用(P>0.05)(表4)。麥季不同處理的CH4排放通量的變化規律基本一致,生物炭和腐殖酸處理對CH4排放通量均沒有顯著影響(P>0.05)。B0F0、B0F2、B1F0、B1F1 和B1F2處理的峰值均在小麥播種后第112 天出現,峰值分別為2.78、3.43、1.93、2.10 和2.88 mg/(m2·h),B0F1 的峰值均出現在小麥播種后第122 天,為1.72 mg/(m2·h),隨后減少且維持較低排放水平(圖2a)。小麥生長期間B0F0、B0F1、B0F2、B1F0、B1F1 和B1F2 的CH4平均排放通量分別為:1.14、0.66、0.86、0.65、0.70 和0.85 mg/(m2·h)。生物炭和腐殖酸處理均顯著降低了麥季CH4累積排放量(P<0.05),腐殖酸和生物炭之間對CH4累積排放量存在顯著交互作用(P<0.05) (表4)。
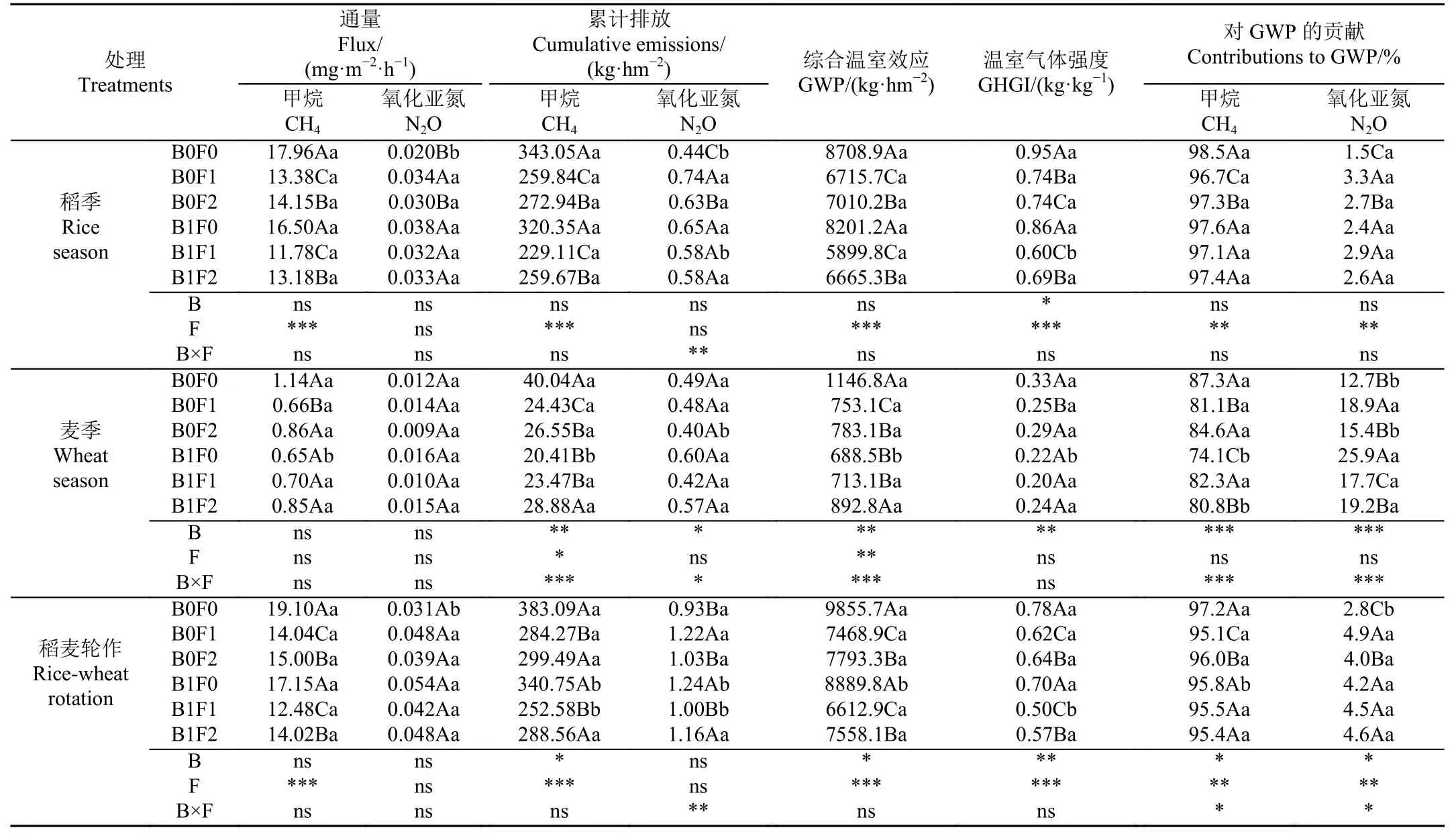
表4 不同處理2021—2022 年稻麥輪作周期內CH4 和N2O 通量、CH4 和N2O 累積排放量、CH4 和N2O 對GWP 的貢獻、GWP 和GHGITable 4 Flux and cumulative emissions of CH4 and N2O, contributions of CH4 and N2O to GWP, GWP, and GHGI under different treatments during rice-wheat rotation cycle in 2021-2022
稻季各處理CH4累積排放量的變化范圍為229.11~343.05 kg/hm2,麥季各處理的CH4累積排放量的變化范圍為20.41~40.04 kg/hm2,稻麥兩季CH4累積排放量變化范圍為252.58~383.09 kg/hm2。CH4排放主要集中在稻季,占稻麥輪作系統周年CH4總排放量的89.5%~94.0%,而麥季CH4排放量僅占總排放量的6.0%~10.5%。在整個稻麥輪作周期內,生物炭和腐殖酸均顯著降低了CH4累積排放量(P<0.05),生物炭和腐殖酸對稻麥輪作周期內CH4累積排放量的影響不存在顯著交互作用(P>0.05)(表4)。與B0F0 相比,單獨施用生物炭或腐殖酸以及二者混施均降低了土壤CH4累積排放量,降幅分別為11.1%、21.8%~25.8%和24.7%~34.1%。
2.3 N2O 排放
稻麥輪作期內不同處理稻田N2O 排放通量的變化趨勢相近,生物炭和腐殖酸處理對N2O 排放通量均沒有顯著影響(P>0.05)。稻田B1F0、B1F1 和B1F2 處理的峰值均在水稻移栽后第8 天出現,峰值分別為0.190、0.142和0.135 mg/(m2·h);B0F0、B0F1 和B0F2 處理的峰值分別出現在移栽后第20、13 和28 天,分別為0.036、0.092 和0.143 mg/(m2·h),隨后減少且維持較低排放水平(圖2b)。水稻生長期間B0F0、B0F1、B0F2、B1F0、B1F1 和B1F2的N2O 平均排放通量分別為:0.020、0.034、0.030、0.038、0.032 和0.033 mg/(m2·h) (表4)。稻季N2O 的排放主要集中在水稻生長的前中期,其原因是稻田前期處于淹水狀態,土壤中以反硝化作用為主,前期基肥和追肥為微生物和植物生長提供了所需的營養物質,為反硝化微生物營造更好的厭氧生存環境,因而N2O 排放量高,而分蘗期后排水曬田,硝化作用處于主導地位,N2O排放少。施用生物炭或腐殖酸對稻季N2O 累積排放量均未產生顯著影響(P>0.05),生物炭和腐殖酸對稻季N2O 累積排放量存在顯著交互作用(P<0.05)。在B0 水平下,添加腐殖酸顯著增加了N2O 累積排放量(P<0.05),與B0F0 相比,B0F1和B0F2 處理的N2O 累積排放量分別增加了65.7%(P<0.05)和40.9%(P<0.05)。在F0水平下,添加生物炭顯著增加了N2O 累積排放量,與B0F0 對比,B1F0 的N2O 累積排放量升高了45.3%(P<0.05)(表4)。麥季不同處理的N2O 排放通量的變化規律一致,生物炭和腐殖酸處理對N2O 排放通量均沒有顯著影響(P>0.05),且兩者對N2O 排放的影響不存在顯著交互作用(P>0.05)。B0F1、B1F0、B1F1 和B1F2處理的峰值均在小麥播種后第112 天d 出現,峰值分別為0.043、0.044、0.031 和0.046 mg/(m2·h),B0F0 和B0F2處理的峰值出現在小麥播種后第26 天,分別為0.023 和0.021 mg/(m2·h),隨后減少且維持較低排放水平(圖2b)。小麥生長期間B0F0、B0F1、B0F2、B1F0、B1F1 和B1F2 的N2O 平均排放通量分別為:0.012、0.014、0.009、0.016、0.010 和0.015 mg/(m2·h)(表4)。麥季前期N2O排放量較低,其原因可能是冬季降雨少和氣溫低,土壤中微生物活性相對較弱,土壤N2O 排放被限制,隨著氣溫回升土壤N2O 排放量逐漸增加,在追肥后達到峰值,隨后迅速減少至較低水平。由表4 可知,生物炭處理顯著增加了麥季N2O 的累積排放量(P<0.05),腐殖酸處理對麥季N2O 累積排放量沒有顯著影響(P>0.05),生物炭和腐殖酸對麥季N2O 累積排放量的影響存在顯著交互作用(P<0.05)。在B0 和B1 水平下,添加腐殖酸對N2O累積排放量均無顯著影響(P>0.05)。在F0 和F1 水平下,生物炭處理對N2O 累積排放量均無顯著影響(P>0.05),在F2 水平下,生物炭顯著增加了N2O 累積排放量(P<0.05),與B0F2 對比,B1F2 的N2O 累積排放量升高了42.9%(P<0.05)(表4)。
研究發現長江中下游地區稻麥輪作中N2O 累積排放量為0.23~4.54 kg/hm2[26-27],本試驗中,在稻麥輪作周內N2O 累積排放量為0.93~1.24 kg/hm2,處于這個范圍之內。各處理排放的 N2O-N(0.93~1.24 kg/hm2)占總施氮量(385 kg/hm2)的0.24%~0.32%,處于全球農業生態系統的0.03%~2.00%的范圍內[28]。
在整個稻麥輪作周期內,生物炭和腐殖酸N2O 累積排放量均無顯著影響(P>0.05),生物炭和腐殖酸對稻麥輪作周期內N2O 累積排放量的影響存在顯著交互作用(P<0.05)(表4)。與B0F0 相比,單獨施用生物炭或腐殖酸以及二者混施均增加了土壤N2O 的累積排放量,增幅分別為33.3%、10.0%~30.1%和7.2%~23.7%。
2.4 綜合溫室效應和溫室氣體排放強度
GWP 表示溫室氣體的綜合增溫效應,GHGI 表示溫室氣體強度。通過對N2O 和CH4綜合增溫效應、氣體貢獻率以及溫室氣體排放強度的計算,可以更直觀地評估生物炭和腐殖酸對本研究中溫室氣體排放與產量的綜合效益。在稻季,生物炭處理對GWP 沒有顯著影響(P>0.05),而腐殖酸處理顯著降低了GWP(P<0.01),生物炭和腐殖酸對GWP 沒有顯著交互作用(P>0.05)。與B0F0 相比,單獨施用生物炭或腐殖酸以及二者混施均降低了稻季GWP,降幅分別為5.8%、19.5%~22.9%和23.5%~32.3%。稻季各處理GWP 變化范圍為5 899.8~8 708.9 kg/hm2,不同處理GWP 從大到小依次為:B0F0、B1F0、B0F2、B0F1、B1F2 和B1F1。CH4是稻季綜合溫室效應的主要貢獻者,其對GWP 的貢獻率范圍為96.7%~98.5%,N2O 對GWP 貢獻率的變化范圍為1.5%~3.3%。在麥季中,生物炭和腐殖酸處理均顯著降低了GWP(P<0.01),生物炭和腐殖酸對GWP 存在顯著的交互作用(P<0.01)。麥季各處理的GWP 變化范圍為688.5~1 146.8 kg/hm2,不同處理GWP 從大到小依次為:B0F0、B1F2、B0F2、B0F、B1F1 和B1F0。CH4在麥季對綜合溫室效應的貢獻率相較于稻季有所降低,其變化范圍為74.1%~87.3%,N2O 對GWP 貢獻率的變化范圍為12.7%~25.9%。在整個稻麥輪作周期內,生物炭和腐殖酸處理均顯著降低了GWP(P<0.05),生物炭和腐殖酸對GWP 沒有顯著交互作用(P>0.05)。稻麥輪作周期內GWP 的變化范圍為6 612.9~9 855.7 kg/hm2,不同處理GWP 從大到小依次為:B0F0、B1F0、B0F2、B1F2、B0F1 和B1F1。稻麥輪作周期內各處理CH4對GWP 的貢獻率范圍為95.1%~97.2%,N2O 對GWP 貢獻率的變化范圍為2.8%~4.9%(表4)。
溫室氣體排放強度(GHGI)可以直觀地評估溫室氣體排放與產量的綜合效益。在本試驗中,在整個稻麥輪作周期內生物炭和腐殖酸添加對作物產量沒有顯著影響,GHGI 的降低要歸因于CH4排放量的減少。稻麥輪作周期內GHGI 變化范圍為0.50~0.78,生物炭和腐殖酸處理均顯著降低了GHGI (P<0.01),生物炭和腐殖酸對GHGI 的影響不存在顯著交互作用(P>0.05)。
與B0F0 相比,在稻麥輪作周期內單獨施用生物炭或腐殖酸以及二者混施處理的GHGI 降幅分別為10.2%,17.3%~20.2%和27.2%~36.2%。B0F0、B0F1、B0F2、B1F0、B1F1 和B1F2 的GHGI 分別為0.78、0.62、0.64、0.70、0.50 和0.57,B1F1 處理最低(表4)。
2.5 環境因子
由表5 可知,在稻麥輪作周期內,CH4和N2O 排放通量與土壤溫度和含水量均存在顯著相關關系(P<0.05),CH4排放通量與土壤pH 值均存在顯著負相關關系(P<0.05),N2O 排放通量與土壤pH 值沒有顯著相關性(P>0.05)。在麥季中,CH4和N2O 排放通量與土壤銨態氮和硝態氮含量均存在顯著正相關關系(P<0.05)。
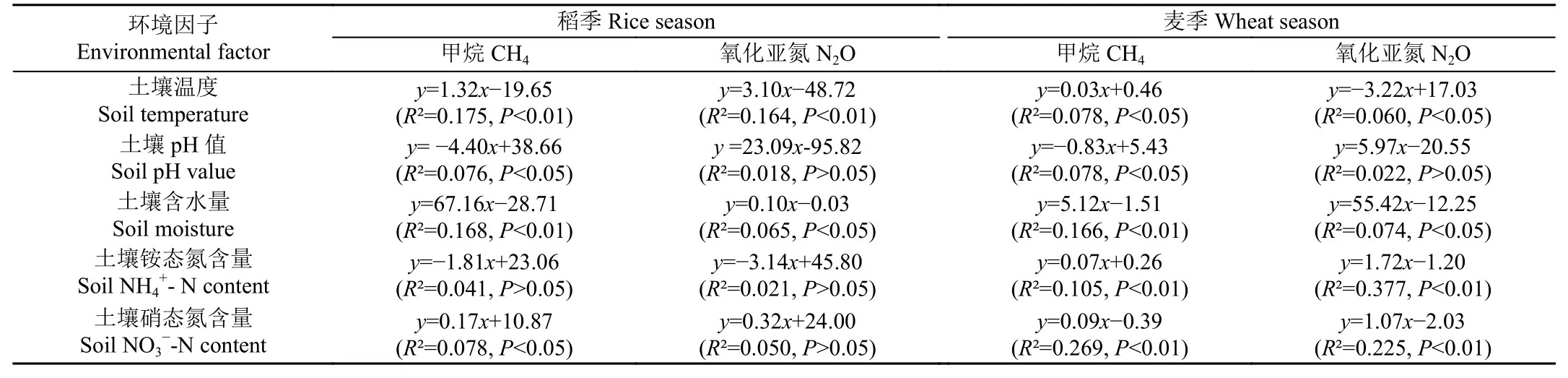
表5 2021—2022 年稻麥輪作周期內CH4 和N2O 排放通量與環境因子的相關方程Table 5 Correlation equations between CH4 and N2O fluxes against environment parameters during rice-wheat rotation cycle in 2021-2022
3 討 論
3.1 不同處理對CH4 排放的影響
生物炭對于CH4的作用效果受到添加量、熱解溫度、原料類型、土壤和植被類型等因素的影響,大量研究表明生物炭能夠抑制土壤CH4排放[9,29]。本試驗中,經過1 a 陳化后的生物炭顯著降低了稻麥輪作周期內CH4累積排放量。生物炭添加導致CH4排放量降低的主要原因有:1)土壤pH 值升高。甲烷氧化菌在酸性環境中活性較強,而產甲烷菌則適合生活在中性或稍堿性環境中,且對pH 值變化很敏感[30]。在本研究中,由表2 可知,生物炭pH 值為8.51,施入后土壤的pH 值在5.29~5.65之間,提高了土壤的pH 值(表3),從土壤pH 值變化影響產甲烷菌活性這個角度來講,生物炭添加對甲烷氧化菌活性增強的程度比產甲烷菌的更大一些,因此使土壤 CH4的最終釋放量減小,從而減少了土壤 CH4的釋放。2)土壤通氣性增加。土壤CH4氧化消耗受到土壤氧氣和養分供應的控制。生物炭疏松多孔,施加后可以改善土壤的通氣環境,提高土壤的透氣性和甲烷氧化活性[31]。3)土壤硝酸鹽含量增加。HU 等[32]認為,硝酸鹽是土壤甲烷氧化菌的重要氮源,可以作為土壤中CH4氧化過程中的底物。而生物炭作為改良劑可以促進土壤硝化作用,提高土壤硝酸鹽含量[33]。4)土壤有機碳含量增加。生物炭添加促進土壤碳循環,表明生物炭增強了土壤對甲烷的氧化能力,增加甲烷氧化菌的活性和豐富度,進而減少甲烷了的產生[34]。
腐殖酸是動植物遺骸經過微生物分解、轉化和地球化學形成的有機物質[15]。本試驗中,經過1 a 陳化后的腐殖酸顯著降低了輪作周期內CH4累積排放量。有研究發現,腐殖酸能作為電子受體并抑制土壤中甲烷的排放[35]。SUN 等[19]研究發現,雖然腐殖酸的施用可以為產甲烷菌提供有機底物,但由于試驗樣地較高的鹽分含量降低了土壤中微生物的活性和豐富度,從而導致腐殖酸的添加對甲烷排放沒有產生顯著影響。
3.2 不同處理對N2O 排放的影響
本試驗中,經過1 a 陳化后的生物炭有增加稻季N2O 累積排放量的趨勢,顯著提高了麥季N2O 的累積排放量。生物炭促進N2O 排放量升高的主要原因有:1)土壤pH 值升高。LI 等[36]研究發現,生物炭改良土壤中N2O 排放的增加歸因于土壤pH 值升高促進了土壤中硝化作用相關微生物的豐富度和活性增加,促進了土壤硝化的進程及N2O 的產生。在本試驗中,生物炭添加顯著提高了麥季生長期內土壤pH 值,這可能是生物炭促進N2O 排放的主要原因。2)土壤含水量增加。生物炭添加下的較高含水量進一步增加了異養硝化和反硝化對N2O排放的貢獻,生物炭的添加與氮肥結合進一步促進土壤反硝化作用,導致高土壤含水量下N2O 的排放增加[37]。在本試驗中,生物炭顯著提高了麥季的土壤含水量,N2O 通量與土壤含水量呈顯著的正相關關系。3)土壤反硝化作用增強。CLOUGH 等[38]研究發現,生物炭增加了土壤反硝化作用的氮基質可用性,促進了N2O 排放。此外,生物炭表面的羧基官能團會抑制土壤中N2O 還原酶電子的傳遞,導致土壤對N2O 還原能力減弱,N2O 的排放量相對增加[39]。
在本試驗中,經過1 a 陳化后的腐殖酸有增加輪作周期內N2O 累積排放量的趨勢。SUN 等[19]研究發現,在有無氮肥施用條件下,添加腐殖酸分別使N2O 累積排放量增加了4%和35%,原因是土壤氧化還原電位與N2O 排放呈顯著正相關,腐殖酸施入后增加了土壤氧化還原電位,進而促進了N2O 排放[40]。此外,CARPENTERBOGGS 等[41]研究發現,腐殖酸可以增強微生物的多樣性和活性,這可能進一步促進了土壤N2O 排放。
3.3 不同處理對GWP 和GHGI 的影響
在本試驗中,稻季GWP 和GHGI 均顯著高于麥季,主要體現在稻季CH4排放量遠高于麥季,這與稻田土壤處于淹水狀態相關。在整個稻麥輪作周期內,經過1 a陳化后的生物炭和腐殖酸均顯著降低了GWP 和GHGI。在稻田或者稻麥輪作系統中CH4排放量決定著綜合溫室效應的大小[42]。在生物炭和腐殖酸經過1 a 陳化后,稻季GWP 的降幅(5.8%~32.3%)相較于第一年(8.2%~43.6%)[43]呈現出降低的趨勢。說明生物炭和腐殖酸對綜合溫室效應的影響隨著施用年限增加呈現出減弱的趨勢。在整個稻麥輪作周期內,各處理的溫室氣體對GWP的貢獻率CH4始終占主導地位,均高于90%。生物炭和腐殖酸添加顯著降低了CH4對GWP 的貢獻率,二者之間存在顯著交互作用,因此,在稻麥輪作系統溫室氣體減排的研究中應該重點關注CH4的排放。
根據已有研究結果,農田土壤生物炭平均施用量為22.63 t/hm2,當施用量范圍為10~20 t/hm2時綜合溫室效應降低幅度最大[44]。從 Meta 分析結果的總趨勢來看,當生物炭施用量不超過30 t/hm2時,GHGI 的減少量一般隨生物炭施用量的增加而增加,但當生物炭施用量大于30 t/hm2時,產量尺度GHGI 的減少量呈下降趨勢[45]。這表明,生物炭帶來的環境效益和農藝效益并不總是隨著生物炭施用量的增加而增加。在稻麥輪作周期內,與B0F0 相比,單獨施用生物炭,腐殖酸以及二者混施處理的GHGI 降幅分別為10.2%、17.3%~20.2%和27.2%~36.2%,B1F1 處理(生物炭添加量為20 t/hm2,腐殖酸添加量為0.6 t/hm2)的GHGI 降幅最大(36.2%),具有最佳的減排效果和農藝效益。
4 結 論
1)在土壤中經1 a 陳化后,生物炭顯著增加了稻麥兩季土壤有機碳含量;腐殖酸增加了稻季土壤有機碳含量,對麥季土壤有機碳含量無顯著影響。單獨施用生物炭或腐殖酸對水稻和小麥產量均沒有顯著影響,生物炭和腐殖酸混施處理顯著提高了小麥產量(增幅為1.0%~5.0%),對水稻產量沒有顯著影響。
2)生物炭和腐殖酸均降低了土壤CH4排放,促進了N2O 排放,生物炭和腐殖酸主要通過抑制土壤CH4排放顯著降低了稻麥輪作系統的綜合溫室效應和溫室氣體排放強度,且兩者混施的效果優于單獨施用生物炭或腐殖酸,B1F1 處理(生物炭添加量為20 t/hm2,腐殖酸添加量為0.6 t/hm2)的綜合溫室效應和溫室氣體強度均為最低。
3)施用1 a 后的陳化生物炭和腐殖酸依然能夠降低稻麥輪作系統的綜合溫室效應和溫室氣體排放強度,但其減排效應隨著施用年限增加呈現了減弱趨勢。因此,未來應基于原位田間試驗補施生物炭和腐殖酸,在較長時間尺度上評價生物炭和腐殖酸的固碳減排潛力,為生物炭和腐殖酸的作用機理研究以及在農田生態系統中的推廣應用提供理論依據。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
豬業科學(2018年4期)2018-05-19 02:04:38
浙江農業科學(2016年11期)2016-05-04 04:16:49
化工進展(2015年6期)2015-11-13 00:27:33
無機化學學報(2014年9期)2014-02-28 17:32:57
