
精母細(xì)胞來(lái)源外泌體中的piR-1207/2107靶向調(diào)控精囊上皮細(xì)胞精囊凝膠蛋白1基因表達(dá)的體外實(shí)驗(yàn)研究
2023-08-18 01:12:20鄔燕倩沈鷺蔣瑩洪葉挺
生殖醫(yī)學(xué)雜志 2023年8期
關(guān)鍵詞:檢測(cè)
鄔燕倩,沈鷺,蔣瑩,洪葉挺*
(1.杭州醫(yī)學(xué)院檢驗(yàn)醫(yī)學(xué)院生物工程學(xué)院,杭州 311300;2.浙江省生物標(biāo)志物與體外診斷轉(zhuǎn)化重點(diǎn)實(shí)驗(yàn)室,杭州 310053;3.浙江中醫(yī)藥大學(xué)附屬第一醫(yī)院,杭州 310006)
PIWI蛋白相互作用RNA(piRNA)是從哺乳動(dòng)物生殖細(xì)胞中分離得到的一類(lèi)與生殖細(xì)胞發(fā)育和精子發(fā)生密切相關(guān)的的非編碼小RNA[1-3]。PIWIL1是PIWI家族蛋白成員之一,其可與piRNA相結(jié)合形成piRNA-PIWI復(fù)合體,該復(fù)合體具有抑制轉(zhuǎn)座子、維持基因組穩(wěn)定性、參與調(diào)節(jié)基因轉(zhuǎn)錄后的調(diào)控以及介導(dǎo)泛素化修飾等功能,并在生殖細(xì)胞的發(fā)育和精子的發(fā)生中發(fā)揮著關(guān)鍵的作用和功能[4-9]。本課題組前期研究發(fā)現(xiàn),與正常對(duì)照組相比,弱精子癥患者精漿中piRNA的種類(lèi)和含量分別有所減少和降低,其中piR-1207和piR-2107顯著降低,進(jìn)一步研究發(fā)現(xiàn)精漿中的piRNAs主要位于外泌體中[10-11]。外泌體中含有核酸(piRNA、miRNA和DNA等)、蛋白質(zhì)和脂質(zhì)等信息分子,其可作為從供體細(xì)胞到受體細(xì)胞的信息傳遞載體,從而發(fā)揮細(xì)胞間物質(zhì)交換和信息交流的作用,精漿外泌體中差異表達(dá)的piRNA是否在受體細(xì)胞中具有調(diào)節(jié)功能,從而介導(dǎo)精子活力至今報(bào)道較少。精囊凝膠蛋白1(SEMG1)主要由精囊分泌產(chǎn)生,參與精液凝膠基質(zhì)的構(gòu)成[12-13]。研究表明,SEMG1是一種生理性的精子活力抑制劑,其可通過(guò)參與精子液化、精子獲能以及精子細(xì)胞膜通透性等方面調(diào)節(jié)精子的活力[12-16]。另有研究表明,與正常對(duì)照組相比,弱精子癥患者精子和精漿中SEMG1蛋白水平顯著增高,SEMG1蛋白水平與精子活力呈負(fù)相關(guān)[17-18]。本研究通過(guò)生物信息學(xué)軟件預(yù)測(cè)發(fā)現(xiàn)piR-1207和piR-2107共同靶向SEMG1;擬進(jìn)一步開(kāi)展體外實(shí)驗(yàn)探討精母胞來(lái)源的外泌體中piR-1207和piR-2107是否對(duì)人精囊上皮細(xì)胞的SEMG1蛋白表達(dá)具有調(diào)節(jié)作用,進(jìn)一步促進(jìn)精子活力調(diào)節(jié)機(jī)制的闡明,以期為臨床工作中弱精子癥的診治提供一定的理論依據(jù)。
材料與方法
一、細(xì)胞及實(shí)驗(yàn)試劑
1.細(xì)胞:小鼠精母細(xì)胞[GC-2spd(ts)]購(gòu)自中國(guó)科學(xué)院細(xì)胞庫(kù),人精囊上皮細(xì)胞(HSVEpiC)購(gòu)自上海佰曄生物科技。本課題組前期研究發(fā)現(xiàn),精母細(xì)胞高表達(dá)piR-1207/2107,是研究piR-1207/2107潛在分子機(jī)制的良好細(xì)胞模型[19]。
2.主要實(shí)驗(yàn)試劑:胎牛血清(FND500,蘇州依科賽)、DMEM培養(yǎng)基(C11995500BT,Gibco,美國(guó))、Opti-MEMTMI減血清培養(yǎng)基(31985070,Gibco,美國(guó))、piR-1207/2107模擬物(piR-NC、piR-1207/2107 mimics)及抑制劑(Anti-piR-NC、Anti-piR-1207/2107)均購(gòu)自上海吉瑪,Trizol分離試劑(15596018,Takara,日本)、逆轉(zhuǎn)錄試劑盒(2620,Takara,日本)、SYBR Green熒光染料(RR430,Takara,日本)、SEMG1兔單克隆抗體(Ab139405,Abcam,英國(guó))、PIWIL1兔多克隆抗體(Ab12337,Abcam,英國(guó))、TSG101兔單克隆抗體(Ab133586,Abcam,英國(guó))、CD63兔單克隆抗體(Ab217345,Abcam,英國(guó))、Alix兔單克隆抗體(Sc-53540,Santa Cruz,美國(guó))、GAPDH鼠單克隆抗體(60004-1-Ig,Proteintech,美國(guó))、辣根過(guò)氧化物酶標(biāo)記的多克隆抗小鼠二抗(Sc-2004,Santa Cruz,美國(guó))、辣根過(guò)氧化物酶標(biāo)記的多克隆抗兔二抗(Sc-2005,Santa Cruz,美國(guó))、LipofectamineTM3000(L3000015,Invitrogen,美國(guó))。
二、研究方法
1.生物信息學(xué)分析:通過(guò)生物信息學(xué)軟件RNAhybrid預(yù)測(cè)piR-1207/2107潛在調(diào)控的靶基因。
2.細(xì)胞培養(yǎng)與轉(zhuǎn)染:用含10%胎牛血清的高糖培養(yǎng)基DMEM培養(yǎng)精母細(xì)胞和HSVEpiC,當(dāng)細(xì)胞密度達(dá)到80%~90%時(shí),以1∶3的比例傳代,將細(xì)胞置于37℃、5%CO2培養(yǎng)箱中培養(yǎng)。
HSVEpiC轉(zhuǎn)染前,將細(xì)胞接種至6孔板中培養(yǎng)至融合度70%~80%。以1個(gè)孔為例,取1個(gè)1.5 ml EP管,加入100 μl Opti-MEMTMI減血清培養(yǎng)基和5 μl脂質(zhì)體Lipo 3000,室溫孵育5 min;另取1個(gè)1.5 ml EP管,加入100 μl Opti-MEMTMI減血清培養(yǎng)基和7.5 μl piR-1207/2107模擬物(piR-NC、piR-1207/2107-mimics)或5 μl piR-1207/2107抑制物(Anti-piR-NC、Anti-piR-1207/2107),室溫孵育5 min;把配制好的上述兩管溶液混合,輕輕混勻后,室溫下孵育15 min,將混合后的試劑均勻地滴加進(jìn)6孔板的目的細(xì)胞中,培養(yǎng)24 h,檢測(cè)HSVEpiC中SEMG1蛋白含量和piR-1207/2107表達(dá)情況。piR-1207/2107模擬物和抑制劑序列見(jiàn)表1。

表1 piRNA模擬物及抑制物核苷酸序列
3.外泌體分離和鑒定:收集精母細(xì)胞上清液于50 ml離心管中,4℃ 500g離心5 min,收集上清液;4℃ 3 000g離心25 min,收集上清液用0.22 μm濾膜過(guò)濾,以去除大顆粒和細(xì)胞碎片;4℃ 110 000g離心2 h;外泌體在離心管底部聚集沉淀,去除上清液后PBS重懸外泌體,4℃ 110 000g離心2 h;去除上清液,在外泌體沉淀中加入200 μl PBS重懸后取出。對(duì)提取的外泌體進(jìn)行粒徑和透射電鏡的測(cè)定。同時(shí)在外泌體中加入RIPA裂解液提取總蛋白,鑒定表征蛋白TSG101、CD63和Alix的表達(dá)情況。
4.精母細(xì)胞外泌體與HSVEpiC共培養(yǎng):在精母細(xì)胞中轉(zhuǎn)染Cy3標(biāo)記的Anti-piR-NC、Anti-piR-1207/2107,6 h后更換新鮮的培養(yǎng)基,培養(yǎng)24 h后,收集細(xì)胞上清液,分離外泌體。提取外泌體分別加入到HSVEpiC共培養(yǎng)24 h。激光共聚焦顯微鏡下觀察含Cy3標(biāo)記的Exo-Anti-piR-NC、Exo-Anti-piR-1207/2107是否進(jìn)入細(xì)胞,并對(duì)外泌體共培養(yǎng)的HSVEpiC進(jìn)行SEMG1蛋白的鑒定和piR-1207/2107表達(dá)含量的測(cè)定。
5.RNA提取和實(shí)時(shí)熒光定量PCR(qRT-PCR)的檢測(cè):采用Trizol法提取細(xì)胞、外泌體中的總RNA,使用AMV逆轉(zhuǎn)錄酶逆轉(zhuǎn)錄成cDNA,轉(zhuǎn)錄方法嚴(yán)格按照試劑盒說(shuō)明書(shū)進(jìn)行。以U6為內(nèi)參,數(shù)據(jù)采用2-ΔΔCT方法分析,引物序列見(jiàn)表2。

表2 qRT-PCR引物序列
6.蛋白提取和蛋白免疫印跡(Western Blot):在收集的細(xì)胞中加入適量的RIPA裂解液,在冰上裂解30 min,提取總蛋白并測(cè)定濃度;在聚丙烯酰胺凝膠中進(jìn)行電泳分離后將蛋白轉(zhuǎn)印至PVDF膜上,5%脫脂奶粉室溫封閉2 h后,加入檢測(cè)抗體:SEMG1兔單克隆抗體(1∶2 000)、PIWIL1兔多克隆抗體(1∶2 000)、GAPDH鼠單克隆抗體(1∶4 000)、TSG101兔單克隆抗體(1∶2 000)、CD63兔單克隆抗體(1∶2 000)、Alix兔單克隆抗體(1∶1 000),4℃孵育過(guò)夜;用TBST洗3次,每次10 min,室溫孵育二抗(1∶4 000)1~2 h,洗滌3次后使用Bio-Rad ChemiDoc成像系統(tǒng)進(jìn)行曝光,并用Image J軟件定量蛋白質(zhì)條帶的強(qiáng)度。
7.激光共聚焦:取適量的細(xì)胞接種到共聚焦小室,鏡下觀察細(xì)胞接種情況;次日,棄去小室中的培養(yǎng)基,沿小室側(cè)壁緩慢加入250 μl 4%多聚甲醛,室溫下固定10 min,棄去固定液,加入250 μl PBS浸洗3次,每次5 min;向小室孔中加入250 μl 0.2%的Triton X-100室溫下處理10 min后棄去,沿小室側(cè)壁緩慢加入250 μl PBS浸洗3次,每次5 min;洗滌后,細(xì)胞中加入Hoechst染料避光搖床孵育30 min,棄染色反應(yīng)液后加入PBS清洗3次,最后沿著小室側(cè)壁加入500 μl無(wú)血清培養(yǎng)基,在激光共聚焦顯微鏡下進(jìn)行觀察和拍攝。
三、統(tǒng)計(jì)與分析
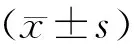
結(jié) 果
一、piR-1207/2107低表達(dá)模型的構(gòu)建
1.信息生物學(xué)分析:通過(guò)核苷酸序列比對(duì)顯示,piR-1207/2107在人類(lèi)、小鼠、大鼠、牛和兔等不同的物種間的核酸序列保守(圖1)。

A:piR-1207核苷酸序列比對(duì)結(jié)果;B:piR-2107核苷酸序列比對(duì)結(jié)果。圖1 不同物種間piR-1207/2107的序列比對(duì)結(jié)果
2.轉(zhuǎn)染精母細(xì)胞:在精母細(xì)胞中轉(zhuǎn)染Cy3標(biāo)記的Anti-piR-1207/2107培養(yǎng)24 h后,激光共聚焦拍攝結(jié)果顯示,Anti-piR-1207/2107主要分布在精母細(xì)胞的細(xì)胞質(zhì)中(圖2)。

紅色為Cy3標(biāo)記的Anti-piR-1207/2107染色;藍(lán)色為Hoechst對(duì)精母細(xì)胞的細(xì)胞核染色。圖2 激光共聚焦拍攝圖
3.qRT-PCR檢測(cè):精母細(xì)胞轉(zhuǎn)染Anti-piR-1207/2107后,qRT-PCR檢測(cè)結(jié)果顯示,與Anti-piR-NC比較,piR-1207和piR-2107的相對(duì)表達(dá)量均顯著降低(P<0.001)(圖3)。

與Anti-piR-NC組比較,***P<0.001。圖3 精母細(xì)胞piR-1207/2107的表達(dá)情況
二、精母細(xì)胞來(lái)源的外泌體的特征
透射電子顯微鏡拍攝精母細(xì)胞來(lái)源的外泌體的形態(tài),結(jié)果顯示,外泌體呈圓形或橢圓形膜性小囊泡(圖4A)。納米激光粒度分析儀檢測(cè)結(jié)果顯示,外泌體的直徑主要分布在100~200 nm之間(圖4B)。Western blot檢測(cè)結(jié)果顯示,精母細(xì)胞來(lái)源的外泌體中表征蛋白TSG101、CD63、Alix呈現(xiàn)陽(yáng)性,但未檢測(cè)到PIWIL1蛋白(圖4C)。qRT-PCR檢測(cè)結(jié)果顯示,精母細(xì)胞來(lái)源的外泌體中存在piR-1207和piR-2107的表達(dá)(圖4D)。

A:透射電子顯微鏡下所見(jiàn),箭頭示外泌體;B:外泌體粒徑分析;C:Western blot檢測(cè)各蛋白表達(dá)的結(jié)果;D:外泌體與精母細(xì)胞中piR-1207/2107表達(dá)量比較:與精母細(xì)胞比較,**P<0.01。圖4 外泌體鑒定及piR-1207/2107表達(dá)情況
三、piR-1207/2107調(diào)控SEMG1蛋白的表達(dá)
1.生物信息學(xué)預(yù)測(cè):采用生物信息學(xué)軟件
RNAhybrid預(yù)測(cè)piR-1207/2107潛在的靶基因,結(jié)果顯示,piR-1207和piR-2107可以共同靶向結(jié)合SEMG1 mRNA的CDS區(qū)(圖5)。

圖5 piR-1207/2107與SEMG1 mRNA的CDS區(qū)結(jié)合示意圖
2.精母細(xì)胞和HSVEpiC中piR-1207/2107及PIWIL1的表達(dá)分析:qRT-PCR結(jié)果顯示,精母細(xì)胞中piR-1207/2107相對(duì)表達(dá)量顯著高于HSVEpiC(P<0.001)。Western blot結(jié)果顯示,精母細(xì)胞和HSVEpiC中都有PIWIL1蛋白的表達(dá)(圖6)。

A:精母細(xì)胞和HSVEpiC中piR-1207/2107的相對(duì)表達(dá)情況:與精母細(xì)胞比較,***P<0.001;B:Western blot檢測(cè)PIWIL1蛋白表達(dá)的結(jié)果。圖6 精母細(xì)胞和HSVEpiC中piR-1207/2107及PIWIL1蛋白的表達(dá)情況
3.HSVEpiC轉(zhuǎn)染piR-1207/2107模擬物:HSVEpiC轉(zhuǎn)染piR-1207/2107模擬物后,與piR-NC相比,轉(zhuǎn)染模擬物組的piR-1207/2107表達(dá)量顯著升高(P<0.001),而SEMG1蛋白表達(dá)水平顯著降低(P<0.01)(圖7)。

A:piR-1207的相對(duì)表達(dá)水平;B:piR-2107的相對(duì)表達(dá)水平;C:Western blot檢測(cè)各蛋白表達(dá)的結(jié)果;D:SEMG1蛋白表達(dá)的灰度分析。與piR-NC比較,**P<0.01,***P<0.001。圖7 piR-1207/2107模擬物調(diào)控SEMG1蛋白的表達(dá)
4.HSVEpiC轉(zhuǎn)染piR-1207/2107抑制劑:HSVEpiC轉(zhuǎn)染piR-1207/2107抑制劑(Anti-piR-1207/2107)后,與Anti-piR-NC相比,piR-1207/2107表達(dá)量顯著降低(P<0.001),而SEMG1蛋白表達(dá)水平顯著升高(P<0.05)(圖8)。

A:piR-1207的相對(duì)表達(dá)水平;B:piR-2107的相對(duì)表達(dá)水平;C:Western blot檢測(cè)各蛋白表達(dá)的結(jié)果;D:SEMG1蛋白表達(dá)的灰度分析。與Anti-piR-NC比較,*P<0.05,***P<0.001。圖8 Anti-piR-1207/2107(抑制劑)調(diào)控SEMG1蛋白的表達(dá)
四、精母細(xì)胞來(lái)源的外泌體中piR-1207/2107跨細(xì)胞調(diào)控SEMG1蛋白的表達(dá)
1.外泌體與HSVEpiC共培養(yǎng):激光共聚焦成像結(jié)果顯示,精母細(xì)胞來(lái)源的Exo-Anti-piR-NC、Exo-Anti-piR-1207和Exo-Anti-piR-2107能夠進(jìn)入到HSVEpiC細(xì)胞中(圖9A)。外泌體與HSVEpiC共培養(yǎng)后,qRT-PCR檢測(cè)結(jié)果顯示,與Exo-Anti-piR-NC相比,Exo-Anti-piR-1207/2107中piR-1207/2107水平均顯著降低(P<0.001)(圖9B、C)。

A:HSVEpiC細(xì)胞外泌體攝入激光共聚焦拍攝圖,紅色為Cy3標(biāo)記的外泌體,藍(lán)色為Hoechst對(duì)HSVEpiC細(xì)胞的細(xì)胞核染色;B:piR-1207的相對(duì)表達(dá)水平;C:piR-2107的相對(duì)表達(dá)水平。與Exo-Anti-piR-NC比較,**P<0.01,***P<0.001。圖9 外泌體piR-1207/2107進(jìn)入到HSVEpiC細(xì)胞
2.Western blot檢測(cè):結(jié)果顯示,與Exo-Anti-piR-NC相比,Exo-Anti-piR-1207/2107組HSVEpiC中SEMG1蛋白表達(dá)水平升高(P<0.05)(圖10)。

A:Western blot檢測(cè)SEMG1蛋白表達(dá)結(jié)果;B:SEMG1蛋白表達(dá)水平的灰度分析:與Exo-Anti-piR-NC比較,*P<0.05。圖10 外泌體中piR1207/2107跨細(xì)胞調(diào)控SEMG1蛋白
討 論
全球約有超過(guò)15%的育齡夫婦受到不孕不育問(wèn)題的影響,其中男性因素約占50%。在中國(guó),不孕不育發(fā)病率也已從2.5%~3%上升到12.5%~15%[20]。精子活力低下導(dǎo)致的弱精子癥是男性不育的重要原因之一[21]。然而,弱精子癥是多種因素作用的結(jié)果,其具體的分子機(jī)制尚不完全清晰。已知piRNA是生殖特異性的非編碼小RNA,其在生殖細(xì)胞的發(fā)育和精子的發(fā)生中發(fā)揮著關(guān)鍵的作用和功能。相關(guān)研究表明,成熟精子中表達(dá)的miRNAs可調(diào)節(jié)精子活力,外泌體中的miRNAs可通過(guò)生殖微環(huán)境介導(dǎo)細(xì)胞間通信,進(jìn)而在精子成熟(如精子活力的獲得和氧化應(yīng)激的減少)和受精過(guò)程中發(fā)揮關(guān)鍵作用[22-26]。與miRNAs相比,piRNAs在生殖細(xì)胞中的表達(dá)更豐富且更具有生殖細(xì)胞特異性,然而研究探索piRNAs介導(dǎo)精子活力的分子機(jī)制的研究較少。本課題組前期研究發(fā)現(xiàn),與正常對(duì)照組比較,弱精子癥患者的精漿中piRNA的種類(lèi)減少、含量降低,其中piR-1207和piR-2107表達(dá)顯著降低,且精漿中的piRNAs主要位于外泌體中[10-11]。本研究使用富含piRNAs和PIWI蛋白的精母細(xì)胞作為探索piRNA潛在分子機(jī)制的體外細(xì)胞模型。我們使用抑制劑(Anti-piR-1207/2107)轉(zhuǎn)染精母細(xì)胞,構(gòu)建piRNA低表達(dá)模型。并通過(guò)提取和分析精母細(xì)胞上清液中的外泌體發(fā)現(xiàn),精母細(xì)胞來(lái)源的外泌體中含有piRNA,可使用精母細(xì)胞作為工具細(xì)胞模擬分泌外泌體的過(guò)程。
人類(lèi)成熟的精子中含有大量非編碼小RNA,精母細(xì)胞中的piRNA可與PIWIL1蛋白及脫腺苷酶CAF1組裝成RNA沉默復(fù)合物調(diào)控大量mRNA的降解[27-29]。本文利用生物信息學(xué)軟件RNAhyrid預(yù)測(cè)發(fā)現(xiàn),piR-1207和piR-2107可以同時(shí)調(diào)控精子活力相關(guān)的SEMG1。既往研究表明,SEMG1是弱精子癥的潛在生物標(biāo)志物,被證明是精子活力抑制蛋白。射精后,精子先被包裹在SEMG1蛋白等組成的精液凝膠中,隨后前列腺特異性抗原(PSA)將SEMG1酶解成小分子的多肽,使得精液凝膠液化并釋放運(yùn)動(dòng)精子[15-18,30]。Mitra等[31]研究發(fā)現(xiàn),在體外加入重組SEMG1,重組SEMG1劑量越多,SEMG1與精子結(jié)合的時(shí)間越長(zhǎng),精子運(yùn)動(dòng)抑制越明顯。另有研究發(fā)現(xiàn),SEMG1蛋白及其水解后的小肽段通過(guò)與精子表面結(jié)合從而抑制精子活力[32-33]。因此,當(dāng)SEMG1蛋白在精液凝膠及精子表面含量異常增高時(shí)將抑制精子活力,導(dǎo)致弱精子癥甚至男性不育。還有研究發(fā)現(xiàn),精子和精漿中高表達(dá)的SEMG1蛋白不僅可造成精子活力差,同時(shí)也會(huì)降低體外受精的成功率[34-35]。
弱精子癥患者的精漿外泌體中piR-1207/2107表達(dá)顯著降低(P<0.05),而SEMG1蛋白表達(dá)顯著增高(P<0.05),并都與精子活力呈負(fù)相關(guān),提示piRNA和SEMG1蛋白在弱精子癥的發(fā)生發(fā)展中具有重要作用。外泌體可作為piRNA載體,發(fā)揮細(xì)胞之間物質(zhì)交換和信息交流的作用,基于piRNA可與PIWIL1組裝成RNA沉默復(fù)合物靶向降解mRNA,本文繼而探究精母細(xì)胞來(lái)源外泌體中的piR-1207/2107能否調(diào)控精囊細(xì)胞中SEMG1蛋白表達(dá)。本研究在HSVEpiC中分別轉(zhuǎn)染piR-1207/2107模擬物和抑制劑,結(jié)果表明在HSVEpiC細(xì)胞中piR-1207/2107可以負(fù)向調(diào)控SEMG1的表達(dá);在精母細(xì)胞中分別轉(zhuǎn)染Cy3標(biāo)記的Anti-piR-NC、Anti-piR-1207/2107,提取外泌體與HSVEpiC共培養(yǎng),激光共聚焦成像結(jié)果顯示,精母細(xì)胞來(lái)源的Exo-Anti-piR-NC、Exo-Anti-piR-1207/2107能夠進(jìn)入到HSVEpiC細(xì)胞中;qRT-PCR結(jié)果顯示,與Exo-Anti-piR-NC相比,Exo-Anti-piR-1207/2107組HSVEpiC中piR-1207/2107水平均顯著降低(P<0.01)。以上結(jié)果表明,精母細(xì)胞來(lái)源的外泌體可有效抑制HSVEpiC中piR-1207/2107的表達(dá)水平。進(jìn)一步研究發(fā)現(xiàn)下調(diào)精母細(xì)胞來(lái)源的外泌體中piR-1207和piR-2107可導(dǎo)致HSVEpiC中SEMG1蛋白表達(dá)量顯著增加(P<0.05)。提示精母細(xì)胞來(lái)源的外泌體中piR-1207/2107可跨細(xì)胞調(diào)控HSVEpiC中SEMG1蛋白的表達(dá)。
目前關(guān)于弱精子癥發(fā)生的分子機(jī)制尚不完全清晰,阻礙了弱精子癥的診斷和治療。本研究以弱精子癥患者精漿和精子中含量高且差異表達(dá)顯著的兩個(gè)piRNA(piR-1207和piR-2107)為切入點(diǎn),通過(guò)體外細(xì)胞實(shí)驗(yàn)探討piRNA在精子活力調(diào)節(jié)中的潛在生物學(xué)功能和下游基因相互作用的分子機(jī)制。但是本研究主要集中在體外細(xì)胞水平,未來(lái)將收集弱精子癥的臨床樣本對(duì)這一潛在的機(jī)制進(jìn)行深入探究,以期為弱精子癥的發(fā)生提出更完善的潛在分子信號(hào)通路。
猜你喜歡
中國(guó)設(shè)備工程(2022年12期)2022-07-11 04:33:00
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:36
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:34
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:50
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:48
