內質網應激預適應對LPS誘導的山羊子宮內膜上皮細胞炎性反應的保護作用
2023-08-25 09:03:18郜康康扆妍妍趙一騰林鵬飛陳華濤靳亞平
畜牧獸醫學報 2023年8期
郜康康,扆妍妍,趙一騰,林鵬飛,陳華濤*,靳亞平*
(1.西北農林科技大學動物醫學院,楊凌 712100;2.西北農林科技大學 農業農村部動物生物技術重點實驗室,楊凌 712100)
內質網(endoplasmic reticulum,ER)是真核細胞蛋白質合成、折疊和運輸的細胞器。當細胞受到各種不良刺激時,ER的蛋白質組裝功能被擾亂,ER腔中出現大量未折疊或錯誤折疊的蛋白質,內質網應激(endoplasmic reticulum stress,ERS)激活,從而引發未折疊蛋白反應(unfolded protein response,UPR)[1]。UPR主要通過肌醇需求激酶1(inositol-requiring kinase 1,IRE1)、活化轉錄因子6(activating transcription factor,ATF6)和蛋白激酶樣內質網激酶(protein kinase-like ER kinase,PERK)3條通路使內質網恢復穩態[2]。UPR是一種保守性的細胞自我保護措施,早期的UPR能及時有效的逆轉內質網應激,增強細胞的存活能力,是細胞應對適度的內質網應激時啟動的防御性反應[3]。當出現過度的ERS之后,則會引發內質網超負荷反應(endoplasmic reticulum-overload response,EOR),EOR主要通過激活NF-κB信號通路,誘導白細胞介素、腫瘤壞死因子、單核細胞趨化因子等炎性因子的表達,進而激活細胞凋亡、細胞炎癥反應和細胞分化等相關信號途徑[4]。有研究表明,在非酒精性脂肪肝形成過程中,UPR可導致炎性小體激活,當出現過度的ERS之后,甚至出現細胞凋亡的現象[5]。此外,在神經炎癥和腸道炎癥過程中也伴隨著ERS的發生,越來越多的研究者將ERS作為炎癥的一個新的研究靶點[6]。
反芻動物分娩后子宮出現細菌感染和組織損傷,是導致子宮內膜炎發生的主要原因,給養殖業帶來巨大的經濟損失;而子宮內膜上皮細胞是抵御細菌入侵的第一道防線,它們是對抗病原體的物理屏障,通過觸發先天免疫反應在防御大多數炎癥性疾病過程中扮演著關鍵角色[7]。反芻動物子宮被革蘭陰性菌(如大腸桿菌)感染后,導致的反芻動物子宮內膜炎通常是由脂多糖(lipopolysaccharide,LPS)介導的[8]。大腸桿菌LPS可通過啟動子宮內膜細胞的免疫反應引起子宮內膜炎癥[9]。LPS可被細胞表面表達的Toll樣受體4(toll-like receptor 4,TLR4)識別,觸發細胞內的信號轉導級聯,導致核因子κB(nuclear factor kappa-B,NF-κB)、絲裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)和含NOD樣受體熱蛋白結構域相關蛋白3(NOD-like receptor thermal protein domain associated protein 3,NLRP3)炎性小體通路的激活,誘導IL-6和TNF-α等炎癥介質的分泌[10-12]。本課題組的前期研究表明,ERS參與LPS刺激誘導的山羊子宮內膜基質細胞炎性反應并促進細胞凋亡[13]。有趣的是,在內皮細胞炎性反應中,抑制ERS和NF-κB信號通路也能夠抑制內皮細胞炎性反應誘導的內皮細胞凋亡[14]。此外,在LPS刺激誘導的小鼠子宮內膜炎模型中,單磷酸腺苷活化蛋白激酶的激活能夠抑制ERS相關的TXNIP/NLRP3炎癥小體活化[15]。
大量的研究證實,ERS作為多種應激源的共同通路,通過UPR與細胞內炎性反應相偶聯,參與機體多種炎癥性疾病的病理過程,但目前關于ERS在反芻動物子宮內膜炎癥反應調控中的作用機制仍未被完全闡明。因此,本研究以山羊子宮內膜上皮細胞(goat endometrial epithelial cells,gEECs)為試驗材料,使用LPS處理構建體外細胞炎性反應模型,探究ERS在LPS誘導的gEECs炎性反應中的作用,為深入解析反芻動物子宮內膜炎的發病機理提供理論基礎。
1 材料與方法
1.1 試劑
衣霉素(tunicamycin,TM,ab120296)購自英國Abcam公司;脂多糖(lipopolysaccharide,LPS,L2630)和4-苯基丁酸(4-phenylbutyric acid,4-PBA,1821-12-1)購自美國Sigma公司;DMEM/F-12細胞培養液(SH30 023.01B)購自美國Hyclone公司;胎牛血清(Z7186FBS-500)購自美國ZETA LIFE公司;RNA Trizol試劑(9109)購自日本TaKaRa公司;ChamQ SYBR qPCR Master Mix(Q311-02)購自南京諾唯贊生物科技股份有限公司;反轉錄試劑盒(AG11711)購自湖南艾科瑞生物工程有限公司;全蛋白提取試劑盒(KGP2100)和BCA蛋白含量檢測試劑盒(KGP902)購自江蘇凱基生物技術股份有限公司;Anti-β-actin抗體(23660-1-AP)和Anti-NLRP3抗體(19771-1-AP)購自武漢三鷹生物技術有限公司;Anti-Phospho-NF-κB P65抗體(3033 T)和Anti-NF-κB P65抗體(8242)購自美國CST公司;Anti-TLR4抗體(sc-293072)購自美國Santa Cruz生物技術公司;HRP標記山羊抗兔IgG(PB001)和HRP標記山羊抗小鼠IgG(PB002)購自陜西中暉赫彩生物醫藥科技有限公司;蛋白Marker(M221-01)和ECL發光液(DE2002)購自北京康潤誠業生物科技有限公司。
1.2 細胞培養
永生化gEECs細胞系為本實驗室前期構建并鑒定保存[16]。將本實驗室保存的30代左右的gEECs置于培養條件為37 ℃,含5% CO2的細胞培養箱中,用含有10%胎牛血清的DMEM/F-12培養基進行培養,當細胞長至匯合度達80%~90%時進行傳代。
1.3 體外細胞模型構建
按照本實驗室前期已經驗證對細胞活性無影響的試驗方法,構建不同的體外細胞模型[17-19]。即將gEECs按照2×105個·孔-1接種至6孔細胞培養板中進行培養,當細胞長至匯合度80%左右時,1)使用LPS(5 μg·mL-1)、TM(0.5 μmol·L-1)和4-PBA(1 mmol·L-1)分別單獨處理6 h后收集細胞樣品。2)使用TM(0.5 μmol·L-1)和4-PBA(1 mmol·L-1)分別預處理2 h,再用LPS(5 μg·mL-1)處理6 h,收集細胞樣品,進行后續試驗。
1.4 實時熒光定量PCR
取“1.3”中收集的各組細胞樣品,用Trizol進行細胞總RNA的提取,并利用反轉錄試劑盒合成cDNA。根據ChamQ SYBR qPCR Master Mix試劑盒說明書在無RNA酶且全程避光的條件下利用Bio-Rad CFX96 PCR儀進行RT-qPCR檢測。試驗所需引物見表1。反應條件:95 ℃ 5 min;95 ℃ 10 s,60 ℃ 30 s,循環39次。采用2-ΔΔCt方法計算試驗結果,以GAPDH為內參基因。

表1 目的基因片段引物序列
1.5 Western blot
取“1.3”中收集的各組細胞樣品,使用細胞蛋白裂解Buffer按照細胞全蛋白提取試劑盒提取細胞全蛋白,利用BCA蛋白含量檢測試劑盒測定蛋白濃度,然后加入5×Loading Buffer,充分混勻煮沸15 min。蛋白樣品進行SDS-PAGE電泳,電泳結束后,將目的蛋白轉移至PVDF膜,置于含10%脫脂奶粉的TBST中封閉2 h。經TBST洗滌后,分別置于Phospho-NF-κB P65(1∶1 000),NF-κB P65(1∶1 000),TLR4(1∶200),NLRP3(1∶1 000)和β-actin(1∶5 000)一抗中4 ℃孵育過夜;TBST洗滌后,置于HRP標記山羊抗兔IgG或HRP標記山羊抗小鼠IgG中室溫孵育1 h;TBST洗滌后,置于ECL發光液中孵育30 s后,使用凝膠成像系統(GBox-Chem-XRQ)拍攝并保存。
1.6 數據統計與分析
2 結 果
2.1 TM處理條件篩選
如圖1所示,與對照組相比,TM處理3 h后即可顯著促進內質網應激通路關鍵基因GRP78和XBP1 mRNA的表達(P<0.001),但對ATF6 的mRNA表達無顯著影響(P>0.05)。隨著TM處理時間的增加,當TM處理6 h后可以觀察到GRP78、XBP1和ATF6的mRNA表達量均顯著增加,且XBP1的mRNA表達量達到最高(P<0.001)。因此,后續試驗將TM處理條件選擇為終濃度0.5 μmol·L-1、處理6 h。

RT-qPCR檢測GRP78、XBP1和AFT6 mRNA表達變化(ns.無顯著性差異;*.P<0.05;**.P<0.01;***.P<0.001。下同)
2.2 TM處理對gEECs炎癥相關基因表達的影響
以本實驗室前期構建的體外細胞炎性反應模型(5 μg·mL-1LPS處理gEECs 6 h)為陽性對照,檢測TM對gEECs炎癥相關基因表達的影響。與對照組相比,TM處理組gEECs炎性細胞因子IL-6和TNF-α 的mRNA表達量顯著降低,同時顯著低于LPS處理組炎性細胞因子的mRNA表達量(P<0.01)(圖2)。此外,如圖2所示,TM處理使經典炎癥通路關鍵基因TLR4、NF-κBP65和NLRP3 的mRNA表達量顯著低于對照組及LPS處理組(P<0.01)。同時,Western blot結果進一步顯示,TM處理顯著抑制了NF-κB P65蛋白的磷酸化和NLRP3蛋白的表達(P<0.05)(圖3)。

RT-qPCR檢測IL-6、TNF-α、TLR4、NF-κB P65和NLRP3的mRNA表達水平

圖3 Western blot檢測TM處理后NF-κB P65蛋白磷酸化和NLRP3蛋白表達水平
2.3 4-PBA處理對gEECs炎癥相關基因表達的影響
添加終濃度為1 mmol·L-1的4-PBA處理gEECs 6 h后,檢測炎癥相關基因的表達變化[16]。如圖4所示,與對照組相比,4-PBA處理組gEECs炎性細胞因子IL-6和TNF-α的mRNA表達量顯著升高,但其表達量仍顯著低于LPS處理組(P<0.01或P<0.001)。接著,對經典炎癥通路關鍵基因進行檢測,結果如圖4所示,4-PBA處理組TLR4和NLRP3 的mRNA表達量顯著高于對照組(P<0.01),而NF-κBP65 的mRNA表達量與對照組相比無顯著性差異(P>0.05);同時,對照組和4-PBA處理組炎癥通路相關基因表達量均顯著低于LPS處理組(P<0.01)。Western blot顯示出相似的結果,如圖5所示,4-PBA處理顯著促進了NF-κB P65蛋白的磷酸化和NLRP3蛋白的表達,但均顯著低于LPS處理組(P<0.05)。

RT-qPCR檢測IL-6、TNF-α、TLR4、NF-κB P65和NLRP3的mRNA表達水平

圖5 Western blot檢測4-PBA處理后NF-κB P65蛋白磷酸化和NLRP3蛋白表達水平
2.4 ERS對LPS誘導的gEECs炎性反應的影響
TM或4-PBA預處理2 h后,再使用LPS刺激,檢測ERS對LPS誘導的gEECs炎性反應的影響。如圖6A所示,與LPS單獨處理組相比,TM預處理顯著抑制了LPS誘導的gEECs炎性細胞因子IL-6 mRNA的表達,而4-PBA預處理則顯著促進了LPS誘導的IL-6 mRNA的表達,且3種不同處理組IL-6 mRNA表達量均顯著高于對照組(P<0.05)。接著,檢測了經典炎癥通路關鍵基因的表達。如圖6B~D所示,TM預處理顯著抑制了LPS誘導的gEECs炎性反應中TLR4、NF-κBP65和NLRP3 mRNA的表達(P<0.05);而4-PBA預處理則顯著促進了TLR4、NF-κBP65和NLRP3 mRNA的表達(P<0.05)。Western blot結果進一步顯示,TM預處理顯著抑制了LPS誘導的NF-κB P65蛋白的磷酸化以及TLR4和NLRP3蛋白的表達(P<0.05);而4-PBA預處理則顯著促進了NF-κB P65蛋白的磷酸化以及TLR4和NLRP3蛋白的表達(P<0.05)(圖6E~I)。
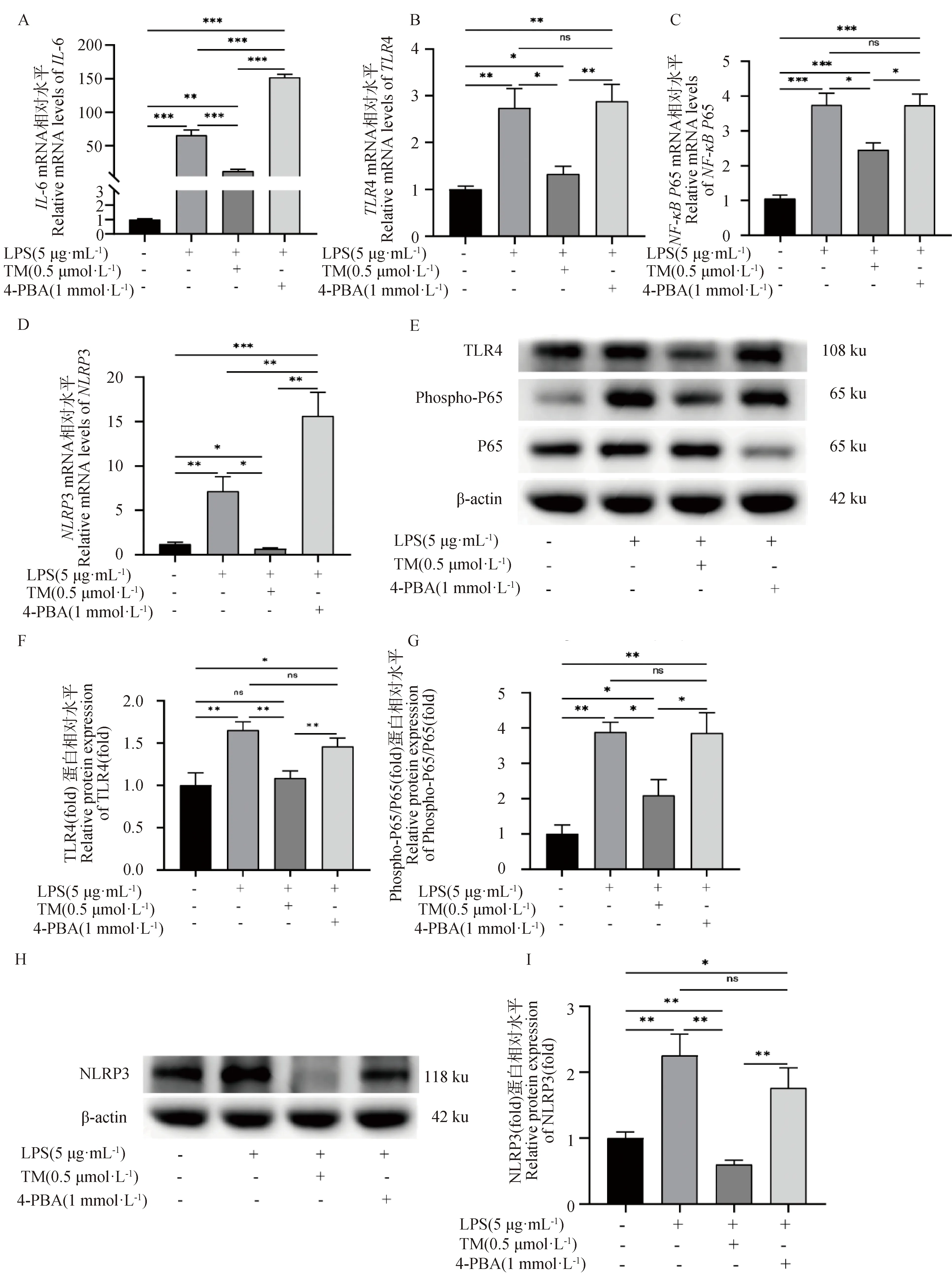
A~D.RT-qPCR檢測IL-6、TLR4、NF-κB P65和NLRP3的mRNA表達水平;E~I.Western blot檢測磷酸化NF-κB P65蛋白以及TLR4和NLRP3蛋白表達水平
3 討 論
動物子宮是一個具有復雜免疫功能的器官,需要在半同種異體胎兒的發育過程中維持免疫耐受環境,同時保持其監測和響應感染因子的能力[20]。然而,分娩后子宮處于一種開放性狀態,細菌感染和組織損傷等可引起子宮內膜炎的發生,給養殖業帶來極大的經濟損失[21]。目前大量研究證實子宮內膜上皮細胞是抵御病原體入侵的第一道防線,子宮內膜上皮細胞的Toll樣受體(toll-like receptors,TLRs)能夠檢測到LPS等病原體相關分子模式(pathogen-associated molecular patterns,PAMP),激活細胞內NF-κB通路等炎癥通路對其產生應答,對機體起保護作用[22]。此外,NOD樣受體(NOD-like Receptors,NLRs)也存在于子宮內膜上皮細胞中,參與子宮抵抗病原微生物入侵的反應[23]。因此,更深入地了解子宮內膜上皮細胞在炎性反應中的功能,對反芻動物子宮內膜炎的防治來說至關重要。越來越多的研究發現ERS通過多種機制與炎癥信號通路相偶聯,包括NF-κB通路和JNK通路等,但ERS在反芻動物子宮內膜炎中的作用仍未被完全闡明[24]。本研究發現,TM誘導的ERS能夠抑制gEECs中炎性細胞因子IL-6和TNF-αmRNA的表達;而用4-PBA處理后則促進了IL-6和TNF-αmRNA的表達。同時,在TM誘導的ERS中,NF-κB P65蛋白的磷酸化和NLRP3蛋白的表達同樣受到抑制;而4-PBA則促進了gEECs中經典炎癥通路關鍵基因的表達。這些結果表明,TM誘導的ERS能夠抑制gEECs中炎癥相關基因的表達,而使用4-PBA處理抑制生理條件下內質網應激的程度可能發揮促炎作用,提示ERS可能參與調節山羊子宮內膜炎的發生和發展。與本研究結果一致,ERS也可減輕游離脂肪酸誘導的3T3-L1脂肪細胞炎癥,抑制NF-κB P65的磷酸化和炎性細胞因子的表達[25]。
ERS發生后激活UPR對外界的細胞不良應激進行應答,UPR信號通路能夠通過對細胞轉錄以及翻譯過程的改變來緩解ERS,以維持細胞內穩態,促進細胞存活[26]。最近的研究發現,UPR通過ERS誘導的信號通路與免疫反應之間的直接串擾發揮免疫作用[27]。此外,也有研究發現,TLR4-Myd88信號通路與PERK/eIF2α/CHOP通路的相互作用在壞死性小腸結腸炎的炎癥發生中發揮著重要作用[28]。值得一提的是,作為適應性UPR的效應器,XBP1s可直接與細胞因子TNF-α和IL-6的啟動子結合調節其表達[29]。在本研究中,作者首先確定了TM處理條件為終濃度0.5 μmol·L-1、處理6 h,此時,發現XBP1的表達量最高,同時內質網應激通路關鍵基因GRP78和ATF6 mRNA的表達量也顯著升高但并非處于最大值,因此,推測TM可能誘導了適應性UPR的發生。有研究表明,一定程度的內質網應激能激活適應性UPR的發生,從而激活機體保護機制發揮保護作用[30]。此外,Ishikawa等[31]在Sec63和XBP1s失活的基礎上開發出了腎小管間質性腎病的遺傳小鼠模型,該模型呈現間質炎癥和進行性腎功能不全的癥狀,同時,XBP1s的異位表達可以減少由Sec63缺失引起的腎間質炎癥,顯示出ERS對機體局部炎癥的抑制作用。本研究發現,TM預處理顯著抑制了LPS誘導的gEECs中IL-6 mRNA的表達,同時顯著抑制了LPS誘導的NF-κB P65蛋白的磷酸化和NLRP3蛋白的表達;而4-PBA預處理則促進了炎癥相關基因的表達。提示TM誘導的ERS預適應激活了適應性UPR,從而減輕LPS誘導的gEECs炎性反應,其可能是通過抑制NF-κB通路和NLRP3炎性小體通路的激活來實現的。這與先前的研究一致,ERS激活UPR對其產生應答,能夠抑制炎癥和氧化應激,保護神經元細胞免受損傷[32]。此外,Ferrè等[33]也發現ERS能夠促進腎炎和膿毒癥等腎病的發生發展,導致腎損傷。總之,上述研究證據提示,ERS在炎癥性疾病的發生發展中具有雙重作用,它既可以抑制炎癥,也可以促進炎癥,造成這一現象的背后的原因仍待進一步研究。
4 結 論
TM誘導的ERS通過抑制NF-κB通路和NLRP3炎性小體通路的激活抑制LPS誘導的gEECs炎性反應;4-PBA處理抑制ERS能夠促進LPS誘導的gEECs炎性反應。本試驗結果為揭示ERS在山羊子宮內膜炎的發生發展中的作用提供了前期理論基礎。
