長期施用餐廚堆肥對水稻生育期雜草群落的影響
2023-08-27 07:58:02宋鵬曹盼劉欣宇方昭林永鋒田光明邵小明
江蘇農業科學 2023年14期
關鍵詞:水稻產量
宋鵬 曹盼 劉欣宇 方昭 林永鋒 田光明 邵小明

摘要:為探究長期施用餐廚堆肥后稻田雜草群落的變化規律,評估雜草群落結構對水稻產量的影響。連續4年設置餐廚堆肥(KF)、雞糞有機肥(CM)、化肥(CF)及對照(CK)4個處理,在2021年對水稻在不同生育期的雜草群落進行監測和取樣,對雜草的種類、密度、生物量、多樣性以及雜草群落與水稻產量間的關系進行分析。結果表明,長期施用餐廚堆肥、雞糞有機肥、化肥均減少了水稻莎草科和闊葉類雜草的密度和生物量,施肥處理的灌漿期雜草總生物量從高到低依次為CM(24.45 g/m2)、KF(21.98 g/m2)和CF(21.79 g/m2),水稻產量從高到低為KF(8 626.50 kg/hm2)、CF(8 071.50 kg/hm2)、CM(7 083.00 kg/hm2);改變了雜草群落多樣性,CF處理雜草群落的 Shannon-Wiener 指數和Pielou均勻度指數較KF和CM處理顯著降低,Simpson優勢度顯著提高;雜草群落結構的NMDS分析結果表明,KF和CM處理的雜草群落與CK有更好的相似性,與化肥相比,施用有機肥對雜草群落結構的改變具有一定的緩沖作用。綜合來看,長期施用餐廚堆肥有利于減少稻田雜草總量并維持較高的雜草群落多樣性和均勻度,減少雜草造成的水稻產量損失,有利于雜草群落的長期穩定。
關鍵詞:餐廚堆肥;稻田;雜草群落;生物多樣性;水稻產量
中圖分類號:S511.06文獻標志碼:A
文章編號:1002-1302(2023)14-0146-07
雜草通過與作物競爭光照、水分和土壤養分等影響作物產量[1],而雜草多樣性在維持生態系統功能方面發揮著重要作用。雜草多樣性不僅可以為農業害蟲的天敵提供棲息環境,降低害蟲的種群數量,還能保持水土流失和促進土壤養分循環[2]。多樣化的雜草群落還有助于維持雜草間的競爭關系,降低雜草與作物的競爭和限制雜草形成優勢物種,以減少雜草對作物產量的負面影響[3]。影響雜草多樣性的因子很多,包括外源肥料投入、種植制度、田間管理等[4-5],一直是雜草管理中關注的熱點。其中,不同施肥條件下土壤中的N、P以及有機質含量都會影響雜草對養分的吸收和雜草生物量,進而影響雜草群落多樣性構成[6],如土壤高氮條件能明顯增加雜草豐富度[7],增加土壤磷含量會提高雜草生物量、莎草科以及莧屬雜草的優勢度[8-9]。因此合理平衡施用N、P、K肥可以減少優勢種數量,維持較高的雜草多樣性[10]。
餐廚堆肥作為一種新型肥料,是居民、餐飲業等食品生產、加工、消費各環節中產生的廢料和食物殘余,經過堆肥化處理得到的有機肥料[11]。已有研究表明,餐廚堆肥可以在改善土壤養分的同時,保障水稻產量[12]。也有研究發現,施用餐廚垃圾作為原料制備的新型有機肥對稻田禾本科和闊葉雜草有抑制作用,平均除草率可以達到80%,顯著增加水稻產量[13]。鑒于餐廚堆肥在廢棄物資源化利用方面的重要作用,卻少有施用餐廚堆肥后對稻田雜草群落變化的研究。因此,本研究通過對連續施用不同類型肥料的稻田雜草群落開展調查,對水稻主要生育期的雜草種類、密度、生物量以及雜草多樣性進行比較,探究施用餐廚堆肥后稻田雜草的演替規律,并通過與水稻產量的聯系,綜合評價餐廚堆肥對稻田雜草群落的影響。
1 材料與方法
1.1 試驗地概況
試驗地位于江蘇省蘇州市臨湖鎮中國農業大學水稻生態農場臨湖基地(31°12′N, 120°45′E)。該地處于北亞熱帶濕潤性季風氣候區,年均氣溫為 16 ℃,降水量為1 160 mm,是水稻適宜生長區。試驗地土壤為長三角地區典型滲育型水稻土,土壤基本理化性質:pH值為6.23,電導率為0.118 mS/cm,土壤速效氮含量為153.42 mg/kg,速效磷含量為 28.19 mg/kg,速效鉀含量為171.67 mg/kg。
1.2 試驗設計
本試驗自2018年開始,試驗所用肥料為餐廚堆肥、雞糞有機肥和化肥。其中,餐廚堆肥為餐廚垃圾生化一體機處理,經過條垛式好氧發酵充分腐熟的堆肥產品(氮磷鉀養分質量比為129 ∶24 ∶20);雞糞有機肥為購買的商品有機肥(氮磷鉀養分質量比為160 ∶117 ∶214);化肥為當地農業部門測土配施的化肥(氮磷鉀養分質量比為16 ∶12 ∶17)。試驗為隨機區組設計,設置3個施肥處理:餐廚堆肥(KF)、雞糞有機肥(CM)和化肥(CF),以不施肥為對照(CK)。各肥料按照等氮量在種植前作基肥施用,每個處理3個重復,隨機區組排列,小區面積為10 m×10 m。各小區用田埂隔開,埂寬為1 m,埂兩邊采用薄膜隔離小區,并設置單獨進出水道防止竄水竄肥。試驗小區在每年的6月到11月種植水稻,水稻品種為南粳46(Oryza sativa Nanjing 46),行間距為15 cm。該試驗地只種植一季水稻,其他時間不種植其他作物。為探究長期施肥后雜草群落原始結構的差異,在維持基本田間管理的基礎上不采取額外除草措施。
1.3 調查方法
2021年,在水稻的拔節期(8月25日)、揚花期(9月16日)、灌漿期(9月26日)和成熟期(10月26日)在每個小區采用5點取樣法,收集1 m×1 m樣方中的全部雜草,帶回實驗室記錄樣方內雜草的種類及數量,并將雜草按種類分裝,剪除根系后在 105 ℃ 條件下殺青1 h,隨后80 ℃條件下烘干24 h后稱其質量,雜草的鑒定主要參考《中國植物志》[14]和“iplant植物智”網站。在水稻成熟期每小區選擇5個1 m×1 m樣方,記錄樣方中水稻穴數,收割后用于脫粒考種計算水稻產量。在上述4個采樣時期用土鉆采集每個小區的土壤表層(0~15 cm)樣品5個,混勻、風干后過篩用于常規養分檢測。土壤養分含量測定參考鮑士旦的方法[15],測定pH值、全碳、全氮、堿解氮、有效磷、速效鉀含量。
1.4 數據處理與分析
雜草多樣性指數使用Past3軟件計算[16],包括物種豐富度(S)、Shannon Wiener多樣性指數(H)、Simpson優勢度指數(D)和Pielou均勻度(J)指數。土壤碳氮比用全碳含量除以全氮含量得到。利用R 4.0.5軟件對雜草密度、雜草生物量以及雜草多樣性指數進行單因素方差分析,用 “Vegan”包基于Bray-Curtis距離對不同施肥處理下的雜草群落進行非度量多維尺度分析(NMDS),用“adonis2”函數進行置換多元方差分析(PREMNOVA),檢驗不同施肥條件下的雜草群落結構差異顯著性,并利用“ggplot2”包進行繪圖。
2 結果與分析
2.1 雜草密度和生物量
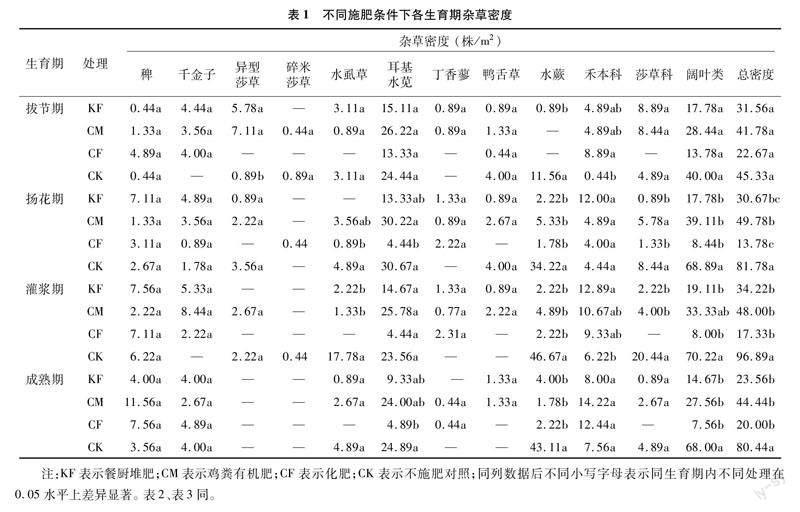
本試驗在不同施肥處理的水稻4個生育期共發現9種雜草,分為禾本科、莎草科和闊葉類雜草三大類。與CK相比,施肥顯著降低了除拔節期外的雜草總密度(表1),在水稻拔節期,各施肥處理的雜草總密度差異不顯著,僅禾本科雜草密度在CK處理中顯著少于CF處理,雜草總密度從大到小分別為CK(45.33株/m2)>CM(41.78株/m2)>KF(31.56株/m2)>CF(22.67株/m2);在水稻揚花期,與CK相比施肥顯著降低了雜草總密度、闊葉類雜草的密度,其中CF處理的雜草總密度最低,為13.78株/m2,CK處理的雜草密度最高,為 81.78株/m2,與拔節期相比,CK處理的雜草總密度增加了80.4%;在水稻灌漿期,施肥增加了禾本科雜草密度,降低了雜草的總密度以及莎草科和闊葉類雜草密度,與CK相比KF處理顯著增加了禾本科雜草尤其是千金子的密度,CF處理中無莎草科雜草被發現,與揚花期相比,CK處理的雜草總密度增加了18.48%;在水稻成熟期,與CK相比,施肥僅對闊葉類雜草密度影響顯著,進而顯著降低了雜草總密度。
從物種來看,與CK相比,施肥主要減少了水虱草(Fimbristylis littoralis)、耳基水莧(Ammannia arenaria)和水蕨(Ceratopteris thalictroides)的密度,從而使莎草科、闊葉類雜草減少,導致總雜草密度顯著減少。隨著水稻生育期的推進,密度上的差距逐漸擴大。各處理間,施肥處理的雜草總密度在各生育期較穩定,而CK處理的雜草總密度隨生育期的推進而呈現整體增加趨勢,在水稻成熟期有所下降。在施肥的3個處理中,與施用化肥(CF)相比,施用有機肥(KF、CM)增加了田間出現的雜草種類數,同時也增加了闊葉類雜草密度和雜草總密度,主要增加了耳基水莧的密度。在施用有機肥的處理中,與CM相比,KF降低了耳基水莧的密度,使雜草總密度減少。
施肥整體明顯增加了水稻各生育期禾本科雜草生物量、減少了莎草科和闊葉類雜草生物量,在雜草總生物量方面也有所減少(表2)。在水稻拔節期,禾本科雜草生物量從大到小為CF(4.55 g/m2)>KF(2.73 g/m2)>CM(1.44 g/m2)>CK(0.10 g/m2)。在水稻揚花期,施肥處理的禾本科雜草生物量,與CK相比,CF處理的雜草總生物量減少了57.40%,KF和CM分別減少了16.71%和14.24%;在水稻灌漿期,與CK相比,KF、CM和CF處理的莎草科雜草生物量減少了91.58%、82.80%和100%,雜草總生物量CF和KF分別減少了29.16%和28.54%;在水稻成熟期,與對照相比,施肥顯著減少了闊葉類雜草生物量,同時增加了禾本科雜草生物量,CF和KF處理的雜草總生物量較CK顯著減少,分別減少了54.05%和51.86%,CM處理減少了19.37%。
具體從物種水平看,與對照相比,施肥減少了水虱草和水蕨的生物量,增加了稗草生物量。與施用有機肥(KF、CM)相比,施用化肥(CF)使莎草科和闊葉類雜草生物量減少,在物種水平上主要是減少了耳基水莧的生物量。在施用有機肥的2個處理中,施用餐廚堆肥(KF)通過減少耳基水莧的生物量減少了雜草總生物量。
2.2 雜草多樣性
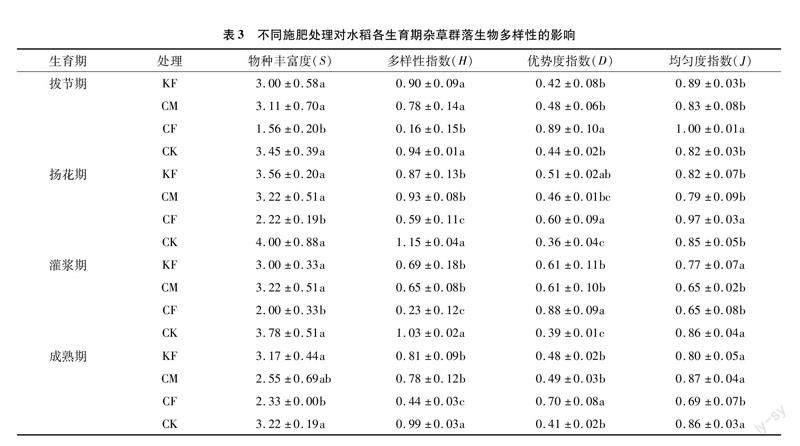
雜草密度和生物量在不同施肥條件下的變化促使各處理間雜草多樣性產生差異。與CK相比,KF、CM和CF處理均減少了雜草群落的物種豐富度和Shannon-Wiener指數,其中CF處理降低效果顯著,說明相較對照,施肥會減少雜草群落中出現的物種數目(表3)。同時,CF處理由于物種數目和多樣性的降低使雜草群落的Simpson優勢度提高,單一物種優勢變大,表明施用化肥會影響雜草間的競爭能力,抑制部分雜草的生長。雜草群落Pielou均勻度指數方面,在水稻生長前期的拔節期和揚花期CF處理的均勻度指數顯著高于其他處理,而隨著生育期的推進而逐漸降低。CK處理的Pielou均勻度指數隨著生育期推進而提高,說明CK處理中每種雜草的生物量較均勻,即每種雜草在與水稻競爭中都具有相同的競爭能力。
在物種水平上,造成CF處理與其他處理間群落多樣性差異的原因是CF處理通過抑制碎米莎草、異型莎草和水虱草的生長使雜草群落的Shannon-Wiener指數下降,并且通過削弱耳基水莧和水蕨的競爭能力提高了群落Simpson優勢度,促使雜草群落均勻度降低。而施用有機肥(KF、CM)時,雜草群落各物種組成均勻,多樣性指數與CK相比差異不大。
2.3 雜草群落結構相似性及對環境因子的響應
NMDS圖中處理間的距離表示各處理雜草群落結構的相似程度,通過對雜草群落的NMDS分析(圖1),表明施肥改變了水稻各生育期的雜草群落結構。在水稻拔節期,施肥顯著影響了雜草群落結構(PREMNOVA test P=0.015),CF處理與CK處理的距離最遠,表明二者在雜草群落的組成和數量方面相差較大,KF和CM處理的原始距離較為接近,說明雜草群落的物種組成和數量較為相似。并且KF和CM與CF相比,前者與CK雜草群落的距離接近,表明相較于CF,KF和CM處理的雜草群落在物種組成和數量上與CK更為相似。在水稻揚花期,不同處理的雜草群落結構呈現明顯分離(PREMNOVA test P=0.012),并且從距離上看,KF與CF的雜草群落相似度較高,CM和CK的相似度較高。水稻灌漿期展示了不同種類的雜草生物量和土壤養分對雜草群落結構的影響(圖1-C),CF、KF和CM處理的雜草群落較為相似,而CK處理的雜草群落與其他3個處理沿莎草科和闊葉類雜草生物量增加方向明顯分離,表明莎草科和闊葉類雜草生物量的差異是促使CK處理雜草群落與施肥處理不同的直接因素。由圖1-C可知,表征土壤養分的堿解氮、有效磷和速效鉀含量與莎草科和闊葉類雜草生物量之間呈負相關關系,表明施肥增加了土壤養分含量,導致雜草群落中禾本科、莎草科和闊葉類雜草組成和生物量的變化,間接引起雜草群落組成的差異。在水稻成熟期,各處理的雜草群落沒有明顯分離,表明在該時期,各處理的雜草群落相似度比較高,不同處理間雜草群落差異不顯著(PREMNOVA test P=0.077)。
因此,與施用有機肥相比,施用化肥對雜草群落組成和數量的改變較大,與CF相比,KF和CM的雜草群落與CK有著更好的相似性,說明相比CF,KF和CM處理對雜草群落結構的改變具有緩沖作用,縱觀整個生育期,長期施肥會顯著改變拔節期、揚花期和灌漿期的雜草群落結構,而到了水稻成熟期,各處理雜草群落各組分的生物量又趨于相似。
2.4 雜草生物量和水稻產量的關系
不同施肥條件下水稻產量如圖2所示,施用餐廚堆肥和化肥后水稻產量顯著高于對照,施用餐廚堆肥、雞糞有機肥、化肥后水稻增產分別達53.23%、25.81%、43.37%,其中施用KF產量最高,為8 626.50 kg/hm2,較CM和CF分別增產21.79%、6.88%,但未達到顯著水平。說明長期施用餐廚堆肥可以顯著提升水稻產量,且效果優于雞糞有機肥和化肥。
為探究不同雜草類群對水稻產量的響應,對各生育期的雜草類群生物量與水稻產量進行回歸分析(表4)。結果表明,在水稻拔節期,禾本科雜草生物量與水稻產量具有顯著的正相關關系,而莎草科和闊葉類雜草與水稻產量表現為不顯著的負相關效應;水稻產量與揚花期莎草科和闊葉類雜草生物量表現為極顯著的負相關關系,與禾本科雜草生物量為不顯著的正相關關系;在水稻灌漿期,禾本科雜草生物量與水稻產量表現為極顯著正相關,與莎草科和闊葉類雜草生物量表現為顯著負相關關系;水稻產量與成熟期的禾本科和莎草科雜草生物量分別呈正相關和負相關,相關性不顯著,與闊葉類雜草生物量呈顯著負相關關系。這說明水稻的產量潛力越高,對于莎草科和闊葉類雜草的生長抑制作用就越強。
3 結論與討論
農田生態系統中,施肥通過促進作物生長和使雜草適應土壤養分的規律性變化,改變雜草和作物的競爭關系,使雜草群落發生演替[1,9]。不同施肥方式對田間雜草總密度和生物量以及不同種類雜草的影響存在差異。有研究表明,長期施用豬糞有機肥和有機無機配施會增加稻田中禾本科雜草的密度,使雜草總密度和生物量增加[17-19];另有研究表明,長期施肥配合秸稈還田顯著降低了莎草科和闊葉類雜草的密度和生物量,而對禾本科雜草密度僅有降低趨勢[20]。在本研究中,施用的3種肥料均抑制了水稻各生育期莎草科和闊葉類雜草的生長,顯著降低了莎草科以及闊葉類雜草的密度,同時減少二者以及總體的生物量,但不同肥料間仍存在差異。相比有機肥,施用化肥對莎草科和闊葉類雜草的抑制效果更好。與施用雞糞有機肥相比,餐廚堆肥處理的雜草群落通過降低耳基水莧的發生數量,使闊葉類雜草的密度在水稻4個生育期分別降低了37.48%、54.54%、42.66%、46.77%,總雜草密度也均降低;在禾本科雜草方面,施肥明顯提升了禾本科雜草生物量;而在密度方面,除水稻揚花期外,其他各生育期施肥均增加了禾本科雜草的密度。造成莎草科、闊葉類和禾本科雜草不同變化的原因可能是由于養分釋放速率和不同種類雜草對光照的敏感度不同[10]。施肥促進了水稻快速生長,同時化肥的養分釋放快,易使水稻迅速形成遮陰環境[21],抑制生長高度較矮的莎草科和闊葉類雜草的光合作用。而在對照中,由于水稻缺乏養分生長緩慢,不利于形成遮陰環境,使莎草科和闊葉類雜草大量生長,反過來抑制了同為禾本科的雜草和水稻的生長。
農田雜草多樣性對于養分循環、保護天敵和控制害蟲方面有重要作用,在農業生態系統中發揮著重要的生態功能[2]。雜草物種豐富度、多樣性、優勢度和均勻度指數可以從不同方面衡量雜草多樣性,多樣性指數反映了雜草群落物種數量、主要雜草的危害情況以及群落中每種雜草是否均勻,雜草種類越豐富,每種雜草越均勻,雜草群落的穩定性也就越高。施用不同肥料可以通過影響雜草群落中各種類型雜草的數量使雜草多樣性指數發生改變。本研究結果表明,化肥處理的物種豐富度和多樣性指數均較低,均勻度也隨著生育期的推進逐漸降低,群落優勢度較高,易使單一優勢雜草演化為惡性雜草。施用有機肥處理維持了較高的物種豐富度和群落多樣性指數,較低的群落優勢度并在水稻生育后期維持較高的群落均勻度。說明施用有機肥有利于調節雜草的種間競爭,降低優勢雜草演化為惡性雜草的可能,利于維持穩定的農田生態系統[18,22-23]。
降低雜草總量同時提高雜草多樣性和均勻度對水稻增產有積極作用。本試驗中,與對照相比,施用3種肥料通過提升土壤養分含量增加水稻養分吸收,顯著提升了水稻產量,并通過增強水稻對雜草的競爭能力,降低了莎草科、闊葉類雜草以及總雜草生物量。本試驗中施肥處理禾本科雜草生物量的增加,是出現禾本科雜草生物量與水稻產量呈正相關的原因,不代表禾本科雜草的增加會促使水稻增產,這可能是由于施肥處理中莎草科和闊葉類雜草減少,使其對禾本科雜草的競爭作用減弱,使禾本科雜草生物量增加。在水稻產量方面,施用餐廚堆肥略高于化肥處理,而雜草總生物量方面化肥處理雜草生物量小于餐廚堆肥,一般來說,雜草生物量越高對水稻產量的影響越大,本試驗中可能是由于餐廚堆肥處理的雜草群落的多樣性、均勻度提高,通過雜草性狀的多樣性限制了雜草與作物間的生態位重疊,減少了雜草與水稻的競爭,降低了雜草總量對水稻產量的負面影響[3,24]。綜合來看,施用餐廚堆肥提升水稻產量的機制是通過減少群落中的雜草總量,改變禾本科、莎草科和闊葉類雜草三者的相對含量,調節群落結構,降低雜草與水稻間的競爭能力,減少雜草對水稻產量的負面效果。相關研究也認為減少雜草生物量同時增加雜草群落均勻度可以減輕作物的產量損失[3],這說明多樣化的雜草群落限制了競爭性和優勢物種對作物生產力的負面影響,同時可能促進雜草多樣化提供的生態系統服務。
長期施用餐廚堆肥可以減少稻田中莎草科和闊葉類雜草的密度和生物量,使雜草總密度和生物量降低。其中,在水稻灌漿期雜草總密度和生物量相比對照分別減少了64.68%、28.54%。施肥顯著改變了水稻各生育期的雜草群落結構,餐廚堆肥可以在降低雜草總密度和生物量的同時增加雜草群落多樣性和均勻度,維持雜草群落的多樣性和穩定性。與施用化肥相比,施用餐廚堆肥的雜草群落多樣性和均勻度顯著提高。長期施用餐廚堆肥可以通過改變水稻和雜草的競爭關系,降低莎草科和闊葉類雜草生物量,平衡不同種類雜草間的競爭關系,減少雜草導致的水稻產量損失。與化肥相比,餐廚堆肥處理中禾本科、莎草科和闊葉類雜草三者占比更均勻,雜草群落組成更穩定,并且在水稻產量方面相比化肥提高了6.88%,有較好的生態效益和經濟效益。
參考文獻:
[1]Tshewang S,Sindel B M,Ghimiray M,et al. Weed management challenges in rice (Oryza sativa L.) for food security in Bhutan:a review[J]. Crop Protection,2016,90:117-124.
[2]陳 欣,王兆騫,唐建軍. 農業生態系統雜草多樣性保持的生態學功能[J]. 生態學雜志,2000,19(4):50-52.
[3]Adeux G,Vieren E,Carlesi S,et al. Mitigating crop yield losses through weed diversity[J]. Nature Sustainability,2019,2(11):1018-1026.
[4]李照全,尹力初,周衛軍,等. 農田管理措施對紅壤稻田系統雜草種群結構的影響[J]. 農業現代化研究,2008,29(2):239-241,245.
[5]趙玉信,楊惠敏. 作物格局、土壤耕作和水肥管理對農田雜草發生的影響及其調控機制[J]. 草業學報,2015,24(8):199-210.
[6]Blackshaw R E ,Molnar L J ,Larney F J . Fertilizer, manure and compost effects on weed growth and competition with winter wheat in western Canada[J]. Crop Protection,2005,24(11):971-980.
[7]Ryan M R,Smith R G,Mirsky S B,et al. Management filters and species traits:weed community assembly in long-term organic and conventional systems[J]. Weed Science,2010,58(3):265-277.
[8]趙 鋒,董文軍,芮雯奕,等. 不同施肥模式對南方紅壤稻田冬春雜草群落特征的影響[J]. 雜草科學,2009,27(1):7-12.
[9]蔣 敏,黃年生,張小祥,等. 施肥對農田雜草生理生態及群落形成的影響綜述[J]. 雜草學報,2016,34(4):1-6.
[10]潘俊峰,萬開元,李祖章,等. 施肥模式對晚稻田雜草群落的影響[J]. 植物營養與肥料學報,2015,21(1):200-210.
[11]Ren Y Y,Yu M,Wu C F,et al. A comprehensive review on food waste anaerobic digestion:research updates and tendencies[J]. Bioresource Technology,2018,247:1069-1076.
[12]滿吉勇,林永鋒,李叢林,等. 餐廚垃圾堆肥對水稻生長、產量及土壤養分含量的影響[J]. 中國農業大學學報,2021,26(11):165-179.
[13]Li Z R. The weeds control of a novel bioorganic fertilizer and its effects on agronomic traits of rice[J]. International Journal of Agriculture and Biology,2018,20(3):507-512.
[14]中國科學院中國植物志編輯委員會. 中國植物志:第十八卷[M]. 北京:科學出版社,1999.
[15]鮑士旦. 土壤農化分析[M]. 3版.北京:中國農業出版社,2000:40-114.
[16]Hammer-Muntz O,Harper D,Ryan P. PAST:paleontological statistics software package for education and data analysis[J]. Palaeontologia Electronica,2001,4(1):1-9.
[17]孫金秋,任相亮,胡紅巖,等. 農田雜草群落演替的影響因素綜述[J]. 雜草學報,2019,37(2):1-9.
[18]李昌新,趙 鋒,芮雯奕,等. 長期秸稈還田和有機肥施用對雙季稻田冬春季雜草群落的影響[J]. 草業學報,2009,18(3):142-147.
[19]董春華,曾希柏,文石林,等. 長期施肥對紅壤雙季稻冬閑田春季雜草群落的影響[J]. 中國生態農業學報,2015,23(9):1150-1157.
[20]張海艷,朱榮松,孫國俊,等. 不同施肥方式對稻麥輪作制稻田雜草群落的影響[J]. 雜草學報,2016,34(1):50-55.
[21]Yin L C,Cai Z C,Zhong W H. Changes in weed community diversity of maize crops due to long-term fertilization[J]. Crop Protection,2006,25(9):910-914.
[22]李儒海,強 勝,邱多生,等. 長期不同施肥方式對稻油輪作制水稻田雜草群落的影響[J]. 生態學報,2008,28(7):3236-3243.
[23]湯雷雷,萬開元,陳 防. 養分管理與農田雜草生物多樣性和遺傳進化的關系研究進展[J]. 生態環境學報,2010,19(7):1744-1749.
[24]Navas M L. Trait-based approaches to unravelling the assembly of weed communities and their impact on agro-ecosystem functioning[J]. Weed Research,2012,52(6):479-488.
收稿日期:2022-09-30
基金項目:國家重點研發計劃(編號:2016YFD0800605);江蘇省蘇州市農業科技創新項目(編號:SNG2020041、SNG2021012)。
作者簡介:宋 鵬(1998—),男,山東日照人,碩士研究生,從事稻田雜草多樣性和廢棄物資源化研究。E-mail:songp@cau.edu.cn。
通信作者:邵小明,博士,教授,研究方向為植物分類和生態學。E-mail:shaoxm@cau.edu.cn。
猜你喜歡
江蘇農業科學(2016年1期)2017-05-17 09:27:45
江蘇農業科學(2017年4期)2017-05-08 21:11:04
南方農業·下旬(2016年9期)2017-04-27 10:11:28
湖北農業科學(2017年6期)2017-04-26 14:22:46
農業與技術(2017年5期)2017-04-11 07:26:43
農民致富之友(2017年4期)2017-04-10 13:10:27
農業與技術(2017年4期)2017-04-05 14:35:48
江蘇農業科學(2016年8期)2017-02-15 19:25:09
江蘇農業科學(2016年8期)2017-02-15 19:24:57
天津農業科學(2015年10期)2015-10-20 00:32:05
