黃鰭棘鯛MSTN1 基因多態性與生長性狀的關聯分析
2023-08-29 02:19:48珠海市現代農業發展中心于方兆盤潤洪李望東李勇楊會兵郭建誼
海洋與漁業 2023年4期
■ 文|珠海市現代農業發展中心 于方兆 盤潤洪 李望東 李勇 楊會兵 郭建誼
中國水產科學研究院南海水產研究所 朱克誠 郭華陽 劉寶鎖 張殿昌
一、前言
黃鰭棘鯛(Acanthopagrus latus),在分類學上隸屬于魚綱(Pisces),鱸形目(Perciformes),鯛科(Sparidae ),棘鯛屬(Acanthopagrus),廣泛分布于紅海、阿拉伯海、印度洋、西太平洋沿岸和中國臺灣、福建、廣東、廣西沿海。黃鰭棘鯛通常在靠近岸邊的海域和河口灣生活,為雜食性魚類,是國內一種重要經濟魚類。但黃鰭棘鯛從魚苗到成年所需的養殖周期較長,一般要經過一年至一年半培養,才能成長到上市規格。近些年來,因為過度捕撈、環境污染等情況使得黃鰭棘鯛自然種群物種質量出現嚴重衰退問題;同時由于黃鰭棘鯛養殖群體的近親繁殖、小規格親本人工育苗等原因,令養殖的黃鰭棘鯛種群物種質量退化程度更嚴重,免疫力下降,養殖黃鰭棘鯛利潤減少等,因此急需進行黃鰭棘鯛的良種選育工作。分子標記輔助選擇育種是目前常用的物種選育手段之一,與目標性狀基因緊密連鎖的分子標記可以用來對目標性狀進行關聯分析,可以明確候選的分子標記與個體表型間的關系,進一步縮短育種年限。
水產養殖收益的檢驗指標之一是種群的生長速率,其受到物種本身的基因型和生存環境的影響,不同的基因型可能會改變蛋白質的表達速度和數量,而處于溫度過高或過低、食物匱乏的環境中生存的生物無法以最大生長速度生長。環境可能會抑制魚類的生長速率,但魚類的最大生長潛力最終由基因型決定。隨著生物技術的不斷創新發展和遺傳學的深入研究,可以選擇分子標記輔助選擇育種方法選擇魚類與其它物種的優良性狀,提高后代生長速率。近年來分子標記輔助選擇育種方法在全世界興盛,它通過利用已知功能的候選基因間接選擇目標性狀,挑選得到目的性狀的基因型,結合傳統育種手段選育出帶有目的性狀品種。與傳統育種相比,它能更方便且準確地選擇隱性的優良性狀,具有選擇準確度高、育種年限短、受環境影響小等一系列的優點。
單核苷酸多態性(Single Nucleotide Polymorphism,SNP)是一種遺傳標記,SNP具有許多特點,一是數量多且分布廣泛。就人類基因組而言,SNP遍布整個基因組中,幾乎每1900bp的片段中就會包含一個SNP位點,正是由于數量巨大的SNP,從而彌補了基因組多態性的不足。二是富有代表性,一些SNP位于基因內部編碼區,這些位點的不同分型可能對基因的表達產生影響,改變蛋白質結構和功能,這類SNP可作為目標性狀的分子選育位點。三是檢測方法簡單,規模化實驗及檢測自動化方面要求低,可以大大縮短工作時間。SNP的堿基類型只有兩種,即雙等位基因。由于SNP具有雙等位基因的特性,在對SNP位點進行檢測時只需對其兩種堿基進行實驗分析,不需要把其所在DNA片段整段測序分析。四是遺傳穩定性高,SNP是單核苷酸突變,突變率較低,與微衛星等重復序列標記相比,具有更高的遺傳穩定性,因此被廣泛應用于和水產動物生長性狀相關聯的研究中。本研究篩選得到黃鰭棘鯛MSTN1基因中分型穩定、多態性好的SNP位點,并與黃鰭棘鯛生長性狀作關聯性分析,為黃鰭棘鯛分子標記輔助育種進程及生長性狀相關基因研究提供理論基礎。
二、材料與方法
1.實驗材料
黃鰭棘鯛(平均體重26.88±7.21g,平均體長10.08±0.96cm)來源于中國水產科學研究院南海水產研究所深圳試驗基地。
(1)實驗所需儀器
本研究使用的儀器包括超低溫保存箱(DW-86L338J,青島海爾股份有限公司)、冰箱(BCD-205TBDZ,青島海爾股份有限公司)、不同最大量程的移液槍(Eppendorf)、渦旋振蕩儀(Vortex-Genie2,美國SI儀器公司)、微量離心機(Legend Micro21,賽默飛世爾科技公司)、分析天平(ClassicL,梅特勒-托利多公司)、電熱恒溫水槽(DK-8D,上海一恒科技有限公司)、高速冷凍離心機(Multifuge X1R,賽默飛世爾科技公司)、電子秤(YH-C,上海英衡電子秤有限公司)、電泳儀(DYY-6C,北京六一生物科技有限公司)、梯度PCR儀(MastercyclerRep gradient S,Eppendorf)、拍48核酸提取儀(上海玉博生物科技有限公司)、全自動數碼凝膠圖像分析系統(Tanon3500,上海天能公司)、燒烤型微波爐(LG)、高壓滅菌鍋(GR85DR,Zealway)、-25℃醫用低溫箱(DW-YL270,中科美菱)。
(2)實驗所需耗材和試劑
本研究使用的試劑和耗材包括磁珠法DNA提取試劑盒(D6310-0 3 B,上海玉博生物科技有限公司)、6×Loading Buffer(9156,北京寶日醫生物技術有限公司)、DL2,000DNAMarker(3427A,北京寶日醫生物技術有限公司)、無水乙醇(廣州普博公司)、Gelred核酸染料(廣州普博公司)、TB GreenRPremix Ex TaqTMII(Tli RNaseH Plus)(RR820A,北京寶日醫生物技術有限公司)、不同量程移液槍槍頭等耗材(北京普博欣生物科技有限責任公司)、瓊脂糖(111860,上海微科生物技術有限公司)等。
2.實驗方法
(1)用于進行生長性狀關聯性分析研究的樣品采集
隨機挑選健康的,有活力的,同池塘養殖的黃鰭棘鯛171尾,測量其體長、體重等數據,做好記錄后取其尾鰭放入裝有無水乙醇的樣品瓶中,并做好標記。
(2)黃鰭棘鯛DNA的提取
取浸泡在無水乙醇中的黃鰭棘鯛尾鰭樣品,用吸水紙擦干后剪下20mg樣品放入1.5mL離心管中用剪刀剪碎,后續提DNA操作用拍48核酸提取儀按上海玉博生物科技有限公司的磁珠法DNA提取試劑盒說明書的步驟提取DNA。將提取后的DNA溶液用1%的瓊脂糖凝膠進行電泳,分析凝膠條帶,檢查DNA的提取質量。
(3)MSTN1基因擴增和測序
基于黃鰭棘鯛基因組序列,應用 Primer5軟件設計多對引物擴增 MSTN基因(表1),研究采用100μL反應體系進行PCR擴增,含 2×PCRMIX50μL,正反向引物各2μL,DNA模板6μL,ddH2O40μL。實驗開始前先進行溫度梯度PCR確定引物的最適退火溫度,PCR反應設置信息:先在94℃下預變性4min;按此設置循環30次:94℃變性40s,最適溫度退火40s,72℃延伸90s;循環結束后在72℃下延伸10min(若擴增片段長度<600kb,設置改為:94℃預變性4min;按94℃變性30s,最適溫度退火30s,72℃延伸40s的時長循環30次;最后72℃延伸10min),延伸結束后在4℃保存。將PCR擴增結束后的DNA溶液用1%的瓊脂糖凝膠進行電泳,分析凝膠條帶,隨后將pcr產物送至北京擎科新業生物技術有限公司進行測序。
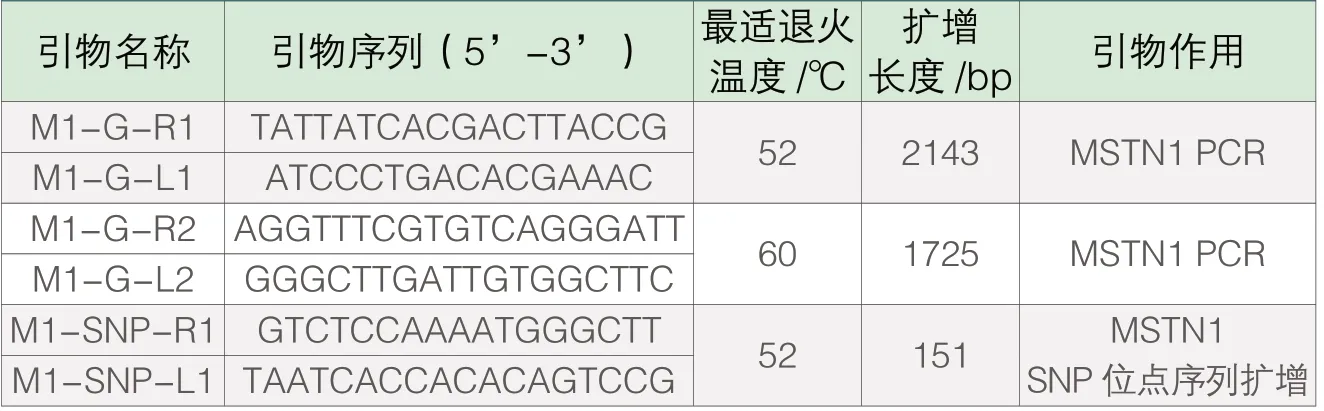
表1 本研究所設計引物的相關信息
(4)SNP位點的篩選和分型
根據體重數據,將10份黃鰭棘鯛大魚DNA樣品和10份小魚DNA樣品進行 PCR擴增和測序后,用ClustalX軟件比對測序結果,篩選MSTN1基因中多態性較高的位點。根據篩選出的SNP位點,用Chromas軟件查看位點所在位置的測序峰圖,并對該位點進行分型。基因型根據峰圖決定,當峰圖為單峰或雙峰中低峰峰值高度不到高峰一半時,此位點基因型認定為是純合;當雙峰中低峰峰值高度達到或超過高峰峰值的一半時,此位點基因型則認定為是雜合(圖1)。

圖1 MSTN1 SNP819位點三種基因型峰圖
(5)SNP位點和生長性狀的關聯性分析
根據黃鰭棘鯛基因組序列設計包含SNP位點的短擴增引物(表1),對用于性狀關聯性分析的171份黃鰭棘鯛DNA樣品進行擴增和測序。重復上述步驟后,利用excel表格記錄每個SNP位點在樣品序列中的位置和對應的基因型。使用SPSS Statistics軟件對SNP位點與黃鰭棘鯛各項生長性狀進行單因素ANOVA(one-way ANOVA)分析,分析與各項生長性狀間是否存在顯著性差異(P<0.05)。
三、結果與分析
1.基因組DNA質量
提取的黃鰭棘鯛鰭條的基因組DNA用1%的瓊脂糖凝膠電泳檢測,部分樣品的電泳檢測結果見圖2。圖中可見提取的基因組DNA條帶明顯且無拖帶現象,完整性較好,沒其它雜帶,提取結果優良。
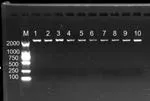
圖2 黃鰭棘鯛部分樣品DNA電泳結果圖
2.黃鰭棘鯛MSTN1基因多態性與生長性狀關聯性分析
采用上述引物擴增來獲得黃鰭棘鯛MSTN1全長序列,混池測序發現MSTN1基因的內含子存在1個SNP位點,將其命名為SNP819。
再以M1-SNP-R1/L1的引物對進行擴增,擴增后使用1%瓊脂糖凝膠電泳檢測,凝膠圖中有均一條帶的樣品進行測序。根據測序峰圖判斷潛在的SNPs位點,接著隨機選取養殖的黃鰭棘鯛個體171尾,并進行基因分型。
經過線性模型分析后,SNP819位點不同基因型與生長性狀的關聯分析結果如表2中所示。

表2 SNP819 位點不同基因型與生長性狀的關聯分析
綜上所述,從表1 中可知,自5’端起第819位,其堿基為A或T的SNP位點的AA和AT基因型個體的軀干長大于TT基因型個體(P<0.05);SNP位點的AA和AT基因型個體的尾長大于TT基因型個體(P<0.01);SNP位點的AA和AT基因型個體的體長大于TT基因型個體(P<0.01);SNP位點的AA和AT基因型個體的尾鰭長大于TT基因型個體(P<0.05);SNP位點的AA和AT基因型個體的全長大于TT基因型個體(P<0.01)。
肌肉生長抑制基因(Myostatin)是控制動物骨骼肌生長的主要基因,屬于轉化生長因子β(TGF-β)超家族,并包含TGF-β前肽域和TGFB域,MSTN基因通過抑制生肌決定因子(MyoD)的轉錄活性抑制肌細胞增殖和分化,限制肌纖維的數量和大小。該基因最先從小鼠骨骼肌的cDNA文庫中篩選獲得。當MSTN基因發生突變會導致肌細胞增生、肌纖維肥大,比如敲除MSTN基因的斑馬魚的肌肉比對照組的明顯發達;向大黃魚幼魚注射MSTN1前肽基因其增重率顯著降低。此外還有真鯛、金頭鯛等均發現MSTN基因中突變位點與生長性狀之間存在著相關性。
采用單因素方差分析不同基因型(頻率低于2.5%的基因排除)與生長性狀的相關性。結果表明,MSTN1 SNP819中TT基因型體長和全長顯著小于 AT和 AA基因型,為劣勢基因型。因此,本結果表明SNP-819位點可應用于以快速生長為目標的黃鰭棘鯛育種材料早期篩選,能夠有效提高育種的效率和縮短育種年限。
猜你喜歡
小讀者(2021年2期)2021-03-29 05:03:48
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
