菜用大豆有機酸的全基因組關聯分析
2023-09-02 07:25:20許文靜張紅梅劉曉慶朱月林陳華濤
華北農學報 2023年4期
關鍵詞:大豆
劉 慧,許文靜,楊 碩,張 威,張紅梅,劉曉慶,朱月林,陳華濤
(1.南京農業大學 園藝學院,江蘇 南京 210095;2.江蘇省農業科學院 經濟作物研究所,江蘇 南京 210014)
菜用大豆(Glycinemax(L.)Merr.)又稱毛豆、鮮食大豆,是指在R6(鼓粒盛期)—R7(初熟期)生育期間采青食用的大豆專用型品種[1]。我國作為世界上最大的菜用大豆出口國,雖然菜用大豆產業已得到快速的發展,但有關菜用大豆食味品質的研究還有待進一步加強,這制約了優質菜用大豆產業健康發展,因此,食味品質的相關研究已成為菜用大豆改良育種方向之一。鮮食風味是菜用大豆食味品質的關鍵因素,其形成與有機酸有著密切的關聯。有機酸作為多種氨基酸生物合成的底物,有利于籽粒中氨基酸的積累從而提高菜用大豆的鮮味[2]。Sowalsky等[3]的研究結果表明,酒石酸、蘋果酸和檸檬酸與澀味之間存在一定的聯系,并且產生的酸味強度具有差異性。因此,不同種類的有機酸有其獨特的風味特征。檸檬酸產生酸感快且酸度高于蘋果酸,但后味時間短;酒石酸略有澀感,酸味爽口;蘋果酸酸味爽口,微有澀苦,呈味時間長,食用適量可給人清爽之感[4],其他有機酸如奎寧酸則主要產生苦味。有機酸不僅能影響菜用大豆的苦味、澀味、酸味,還能通過掩蓋糖的味道來影響甜味,Green等[5]的研究發現,檸檬酸可以掩蓋味覺細胞對于果糖和蔗糖的感知,而毛岳忠[6]研究發現,蘋果酸能增強味覺細胞對于蔗糖的感知。菜用大豆籽粒中有機酸的種類、含量及其構成比例是菜用大豆鮮食風味形成的重要基礎,因此,檢測菜用大豆的有機酸成分分布,研究有機酸合成機制對于菜用大豆的品質改良具有重要的實際意義。
數量性狀的研究對于農作物的遺傳改良具有重要的實際意義,在不同的植物中已報道了許多與有機酸含量相關的QTLs。Liebhard等[7]研究表明,蘋果果實中可滴定酸含量的變化幾乎完全可以由8,16號連鎖群上的2個主效QTL位點決定,分別能夠解釋后代中酸度表型變異的46%,42%,其中位于第16連鎖群上的主效基因位置與Maliepaard等[8]報道的結果相一致。菜用大豆中的酸含量屬于多個基因協同控制的數量性狀,但截至目前,有關菜用大豆酸含量調控機理的研究相對滯后,與菜用大豆有機酸含量相關的SNPs及候選基因有待發掘。與傳統的連鎖定位方法相比,全基因組關聯分析法(Genome wide association study,GWAS)節省了構建群體的時間,能夠對不同種質進行遺傳結構分析,提高QTL定位的精準性,已經成為快速有效的發掘植物數量性狀基因的重要手段[9-10]。本研究以含有264份菜用大豆種質資源的自然群體為試驗材料,分別在2020,2021年通過高效液相色譜法(High performance liquid chromatography,HPLC)測定酒石酸、蘋果酸和檸檬酸含量,結合GWAS鑒定與有機酸顯著關聯的SNP位點并進一步挖掘相應的候選基因,旨在初步解析菜用大豆有機酸含量的遺傳學基礎,為調控菜用大豆有機酸含量的分子機制研究提供理論基礎。
1 材料和方法
1.1 試驗材料
264份菜用大豆種質資源,包括212份栽培種及52份地方種,均由江蘇省農業科學院經濟作物研究所大豆課題組提供。分別于2020年6月和2021年6月種植于江蘇省農業科學院南京市六合基地試驗田(32°28′12″N,118°37′48″E)。田間試驗采用隨機區組試驗設計,3次重復。每份材料重復種植3壟,每壟播種10穴,每穴2株苗,壟長為1.5 m,壟間距40 cm,穴距15 cm。田間管理按照當地常規方法進行。
1.2 儀器與試劑
植物組織研磨儀;高效液相色譜儀Agilent1100;Aglient ZORBAX SB-Aq色譜柱;超聲波振蕩儀;可見分光光度(UV-2600,日本島津);超純水機;ML204型萬分之一天平;酸度計、離心機、水浴鍋、烘箱及其他實驗室常用設備。
甲醇(色譜純);磷酸(質量分數≥85.0%);蘋果酸、酒石酸、檸檬酸(色譜級純分析標準品),以上試劑均購自上海阿拉丁生物科技有限公司。
1.3 菜用大豆有機酸檢測
1.3.1 色譜條件 色譜柱:Aglient ZORBAX SB-Aq C18柱(粒徑5 μm,柱長250 mm×4.5 mm);流動相:甲醇-0.1%磷酸水溶液(pH=2.30),梯度洗脫;流速0.8 mL/min;檢測波長212 nm;色譜柱溫度30 ℃;進樣量體積10 μL。流動相洗脫條件見表1。
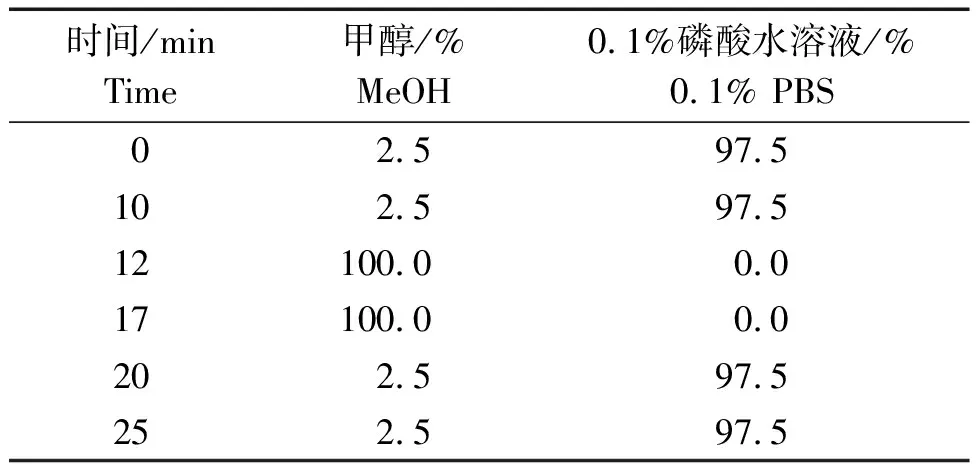
表1 流動相洗脫條件
1.3.2 標準液的配制 精密稱取酒石酸、蘋果酸各50 mg,檸檬酸10 mg(精確至0.1 mg),用流動相溶解并定容至10 mL棕色容量瓶中。配制成酒石酸、蘋果酸、檸檬酸標準品儲備液,質量濃度分別為5 000,5 000,1 000 mg/L。
1.3.3 繪制標準曲線 以1.3.2中的標準品儲備液為母液,共配制5種不同濃度梯度混標溶液。其中蘋果酸和酒石酸濃度分別為5,50,200,1 000,5 000 mg/L,檸檬酸濃度分別為1,10,40,200,1 000 mg/L。以上溶液均經0.45 μm微孔濾膜過濾后,按照1.3.1的色譜檢測條件,利用高效液相色譜儀分析后,以濃度x(mg/L)對色譜峰峰面積y繪制標準曲線,并計算回歸方程。
1.3.4 有機酸提取方法 根據蓋鈞鎰等[11]的報道,菜用大豆最適宜的采收期為R6期,因此,本試驗選擇R6期(鼓粒盛期)即鮮莢翠綠,籽粒飽滿時進行取樣,設置3次生物學重復。將收取的鮮莢放入烘箱先105 ℃殺青30 min,然后以80 ℃烘干至恒質量,取出干籽粒放入植物組織研磨儀中(40 Hz,2 min)研磨至粉末備用。菜用大豆干籽粒粉末過0.45 mm篩,稱取粉末1 g(精確至0.000 1 g),置于50 mL量瓶中,加入40 ℃蒸餾水約40 mL,40 ℃下超聲提30 min,冷卻至室溫,定容,混勻。在4 000 r/min下離心20 min,取上清液1 mL,經0.45 μm濾膜過濾,超聲脫氣后,供分析用。
1.3.5 HPLC分析菜用大豆有機酸 按照1.3.1的色譜檢測條件,從同一菜用大豆樣品中平行取3份進行重復性試驗,根據標品繪制的標準曲線分析計算菜用大豆樣品中各有機酸組分的質量濃度,取3次進樣測得的平均值作為菜用大豆有機酸組分最終的檢測結果。
1.4 數據處理
各有機酸含量數據采用Excel 2019計算平均值、標準差及變異系數,應用SPSS(IBM SPSS Statistics 26.0,SPPS,USA)軟件分別對酒石酸、蘋果酸和檸檬酸含量進行相關性分析。
1.5 全基因組關聯分析
本研究中,用于全基因組關聯分析的自然群體SNP標記來自前期的重測序工作,共2 597 425個,該群體的連鎖不平衡衰減區間為120 kb,具體群體結構分析見江蘇省農業科學院豆類作物研究室的研究報道[12],采用基于群體結構(Q)+親緣關系(K)的混合線性模型(Mixed linear model,MLM)進行全基因組關聯分析以控制假陽性關聯位點。全基因組關聯分析采用基于R軟件的GAPIT算法包進行計算[13],設定-log10(P)=5.0作為顯著閾值,當關聯到的SNP閾值≥5.0時,則被認為是顯著關聯位點。
1.6 候選基因分析
全基因組關聯分析得到2個不同的環境下(2020,2021年)檢測到的顯著SNP位點后,挑選出其上下游120 kb區間范圍內所有的基因。參照在線數據庫Phytozome 13網站(https://phytozome.jgi.doe.gov/pz/portal.html)中的大豆基因組注釋信息確定目標性狀的候選基因。
2 結果與分析
2.1 HPLC色譜分析
在1.3.1的色譜檢測條件下,采用HPLC法分離并檢測有機酸標準品和菜用大豆籽粒樣品中的酒石酸、蘋果酸和檸檬酸,可以清晰地將3種有機酸分離出來,且峰型良好,洗脫順序分別為酒石酸、蘋果酸、檸檬酸,平均保留時間分別為4.316,5.410,9.868 min(圖1)。
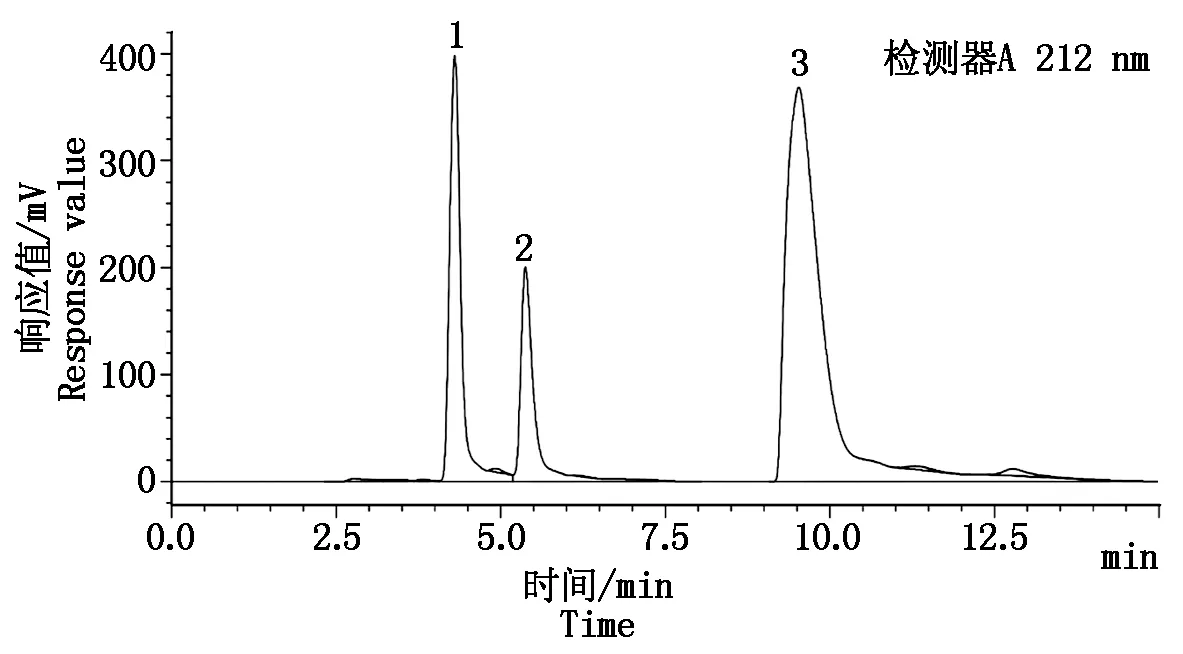
1.酒石酸;2.蘋果酸;3.檸檬酸。
2.2 菜用大豆3種有機酸組分線性回歸方程
分別將不同濃度梯度的3種有機酸的混合標準溶液經高效液相色譜儀分析后,以質量濃度為橫坐標(x),峰面積為縱坐標(y)進行線性回歸分析。計算得到酒石酸、蘋果酸和檸檬酸標準品的線性回歸方程(表2)。3種有機酸標準品的峰面積和質量濃度的相關系數為0.997 9~0.999 1,在設定的濃度范圍內呈現良好的線性關系,根據信噪比確定的定量限(RSN=10)為2.5~10 μg/mL。
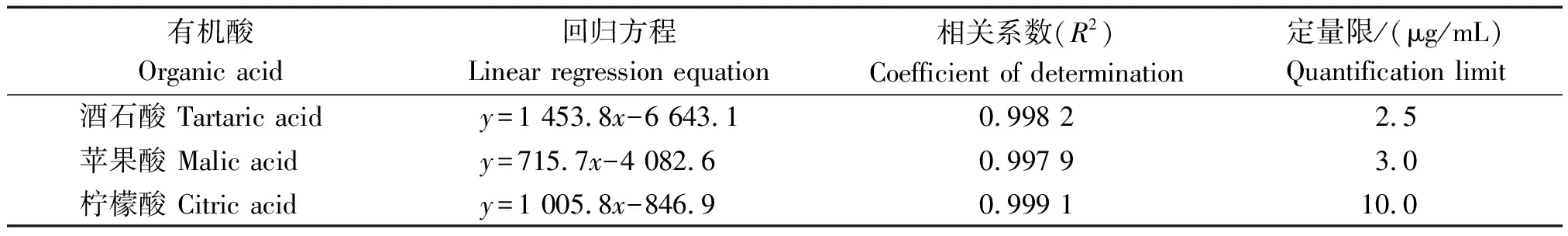
表2 菜用大豆3種有機酸組分線性回歸方程
2.3 表型分析
菜用大豆3種有機酸含量的描述性統計分析(表3)顯示,2020,2021年供試群體的酒石酸含量分別為1.66~9.19 mg/g,0.48~13.52 mg/g,平均含量分別為4.13,4.16 mg/g;蘋果酸含量分別為1.47~36.46 mg/g,1.89~25.19 mg/g,平均含量分別為7.26,8.99 mg/g;檸檬酸含量分別為2.56~12.05 mg/g,1.54~18.31 mg/g,平均含量分別為7.12,10.88 mg/g。上述結果均表明,供試群體有機酸含量差異較大,種質資源遺傳變異豐富,為特異種質的篩選提供了有利條件。
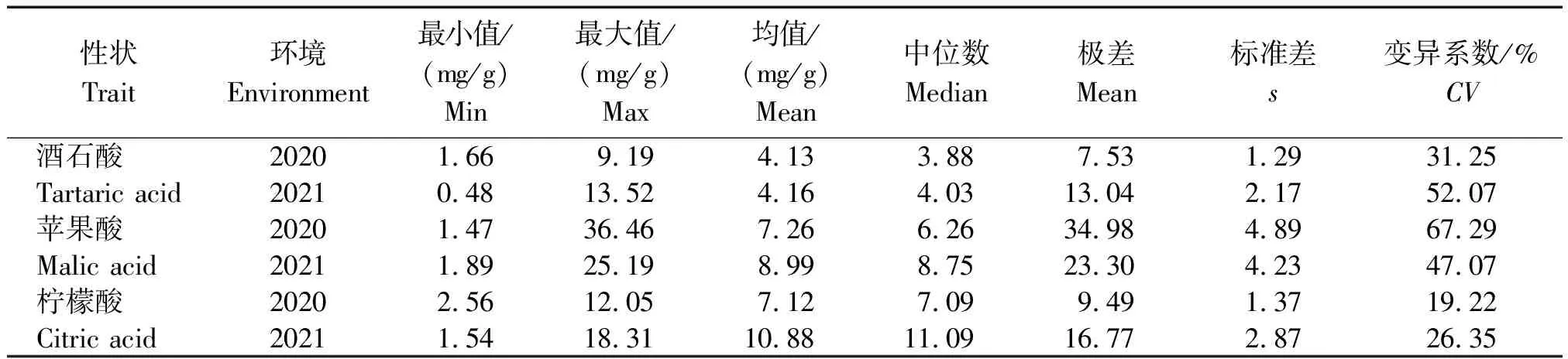
表3 菜用大豆3種有機酸含量的描述性統計分析
2020年菜用大豆酒石酸、蘋果酸和檸檬酸變異系數分別為31.25%,67.29%,19.22%,2021年3種有機酸變異系數分別為52.07%,47.07%,26.35%,且3種有機酸組分中檸檬酸變異系數最低。遺傳變異系數能夠體現各性狀的變異豐富度,一般來說,當變異系數大于10%時,可以認為種質間差異較大,且變異系數越大,表明該性狀的變異潛力越豐富。2 a中3種有機酸的變異系數均大于10%,反映出供試群體不同品種間有機酸各組分存在豐富的變異,這些遺傳變異的存在可為菜用大豆食味品質的改良提供廣泛的遺傳學基礎。
2.4 相關性分析
本研究對264份材料酒石酸、蘋果酸、檸檬酸含量在2020,2021年2 a間進行相關性分析,結果表明,264份材料檸檬酸含量在2020,2021年,2 a間呈顯著正相關,相關系數為0.156*(表4)。同時,基于供試群體3種有機酸2 a表型數據的平均值,對不同有機酸組分含量進行相關性分析。結果發現,酒石酸含量與蘋果酸、檸檬酸含量間均存在顯著正相關,相關系數分別為0.695*,0.739*,蘋果酸含量與檸檬酸含量也有著顯著的正相關,相關系數為0.790*(表5)。

表4 264份材料酒石酸、蘋果酸、檸檬酸含量在2020,2021年2 a的相關系數
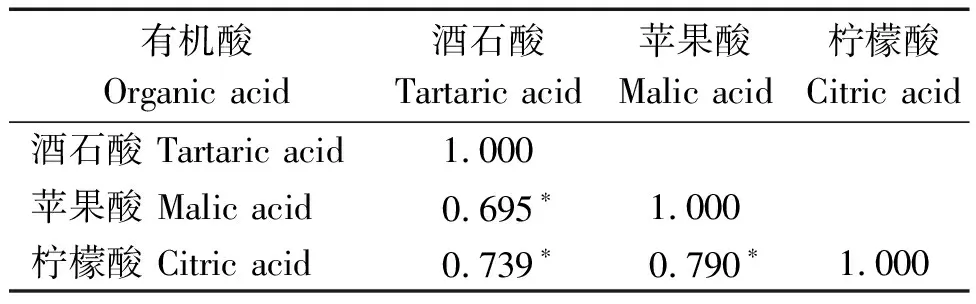
表5 菜用大豆3種有機酸組分相關性分析
2.5 菜用大豆3種有機酸組分含量特異種質篩選
基于供試群體3種有機酸含量,分別對每種有機酸篩選出2份高含量的特異種質。將264份菜用大豆種質資源按照酒石酸含量由高到低排序,并將位于前15%的品種劃分為高酒石酸含量組(Tartaric acid high content group,T1),在2020,2021年的檢測中均屬于T1組的種質為阜04-35和桂春豆112(表6)。同理,將供試群體按照蘋果酸含量由高到低排序,并將前15%的品種劃分為高蘋果酸含量組(Malic acid high content group,M1),2 a中均屬于M1組的種質包括華春9號和邯12-204(表7);將供試群體按照檸檬酸含量由高到低排序,前15%的品種被劃分為高檸檬酸含量組(Citric acid high content group,C1),2 a中均屬于C1組的種質有灌豆3號和新沂大紫花(表8)。以上篩選到的種質,進一步豐富了我國菜用大豆有機酸含量特異種質資源。
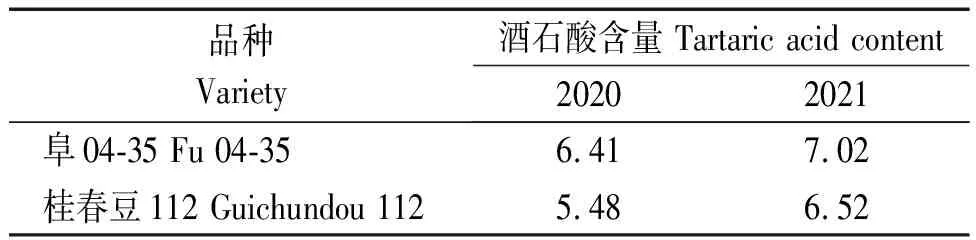
表6 2 a的檢測中均篩選到的高酒石酸含量的特異種質
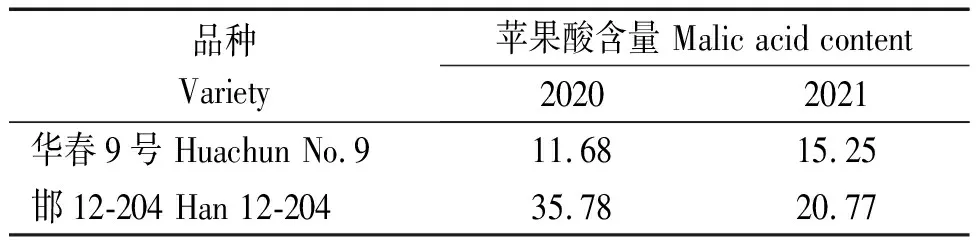
表7 2 a的檢測中均篩選到的高蘋果酸含量的特異種質
2.6 菜用大豆酒石酸、蘋果酸和檸檬酸含量GWAS分析
為了深入研究菜用大豆有機酸含量的遺傳機制,利用MLM模型對2020,2021年的酒石酸、蘋果酸和檸檬酸含量表型值進行GWAS分析(圖2,3),設定顯著閾值為-log10(P)= 5.0,如表9所示,2020年檢測到與酒石酸顯著關聯的SNP位點31個,每個SNP解釋了7.99%~11.23%的表型變異;2021年檢測到與酒石酸顯著關聯的SNP位點23個,每個SNP解釋了7.00%~10.62%的表型變異。2020年檢測到與蘋果酸顯著關聯的SNP位點169個,每個SNP解釋了7.39%~24.66%的表型變異;2021年檢測到與蘋果酸顯著關聯的SNP位點20個,每個SNP解釋了7.30%~9.05%的表型變異。在2020年并沒有檢測到與檸檬酸顯著關聯的位點,2021年檢測到與檸檬酸顯著關聯的SNP位點43個,分別位于2,6,13號染色體上,單個SNP的表型變異解釋率為8.06%~10.96%。上述結果表明,菜用大豆籽粒中有機酸含量是由多基因控制的。
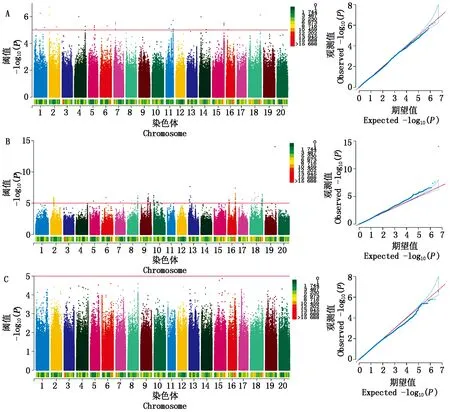
A、B和C分別是酒石酸、蘋果酸和檸檬酸的GWAS結果。圖3同。
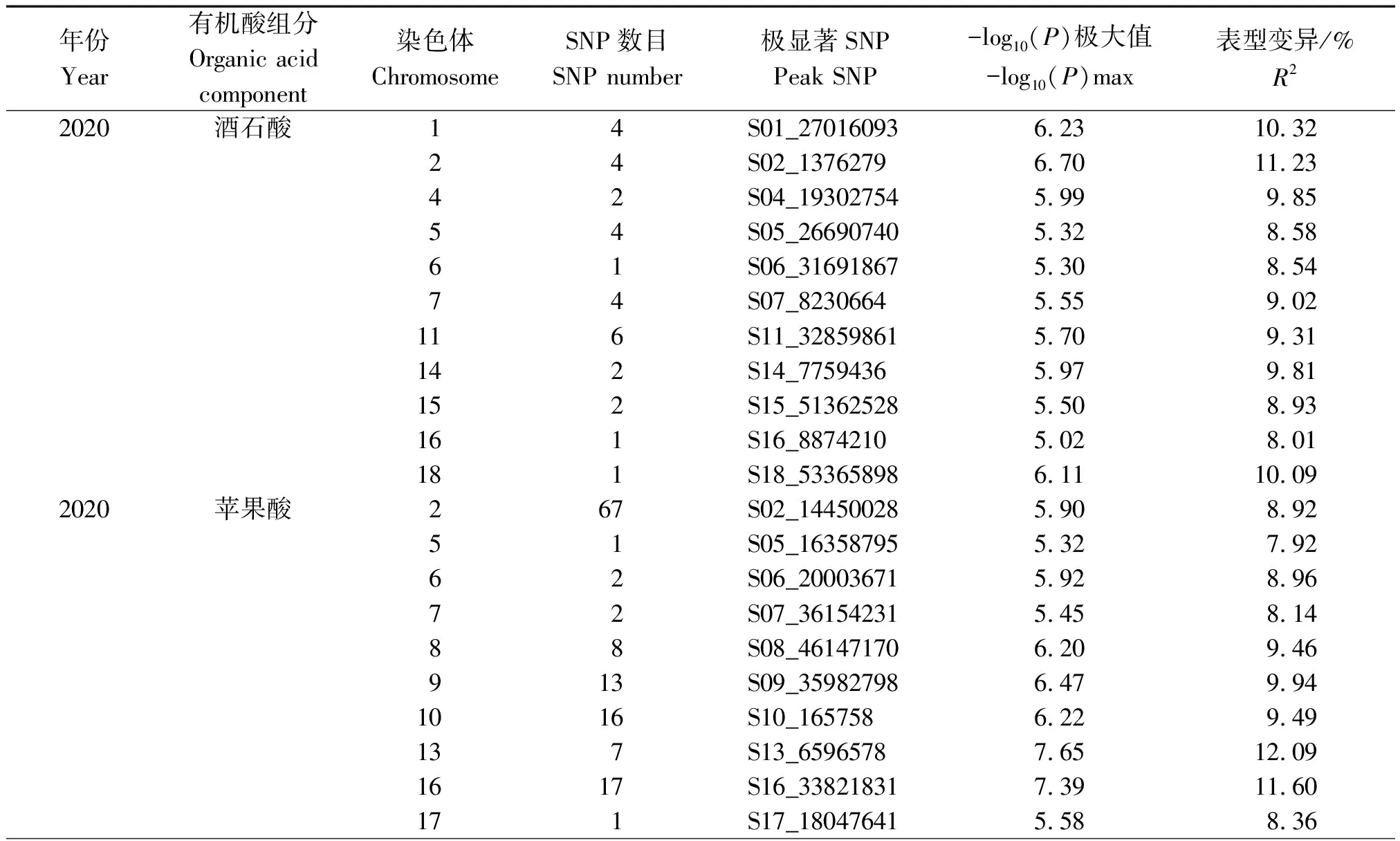
表9 3種有機酸組分顯著相關SNPs分布
2.7 菜用大豆3種有機酸組分相關候選基因挖掘
為了研究顯著關聯的SNP位點等位變異的表型效應,分別對2020,2021年中酒石酸、蘋果酸和檸檬酸檢測到的閾值最高的顯著SNP位點進行單倍型分析。發現SNP S02_1376279堿基變化為A至T,攜有SNP S02_1376279-A的菜用大豆種質的酒石酸平均含量為4.01 mg/g,顯著低于基因型S02_1376279-T的菜用大豆種質的6.40 mg/g(圖4-A)。S19_44610972的等位變異為C/T,攜有SNP S19_44610972-C的菜用大豆種質的蘋果酸平均含量為6.52 mg/g,顯著低于基因型S19_44610972-T的菜用大豆種質的19.52 mg/g(圖4-B)。S17_41027045的等位變異為T/A,攜有S17_41027045-T的菜用大豆種質的酒石酸平均含量要顯著低于攜有S17_41027045-A的菜用大豆種質的酒石酸平均含量(圖4-C)。S13_25565436和S06_6589288的等位變異均為G/A,攜有S13_25565436-G的菜用大豆種質的蘋果酸平均含量要顯著低于攜有S13_25565436-A的菜用大豆種質,而攜有S06_6589288-G的菜用大豆種質的檸檬酸平均含量顯著高于攜有S06_6589288-A的菜用大豆種質(圖4-D、E)。

同一圖中不同小寫字母表示在0.05水平下差異顯著。
在檢測到的與3種有機酸顯著關聯(-log10(P)≥ 5.0)的SNP位點上下游120 kb范圍內進行候選基因的篩選及功能預測。參照大豆參考基因組的基因功能注釋信息,鑒定到8個與有機酸含量顯著相關的候選基因(表10),這些基因可能參與菜用大豆籽粒中有機酸的調控和積累,作為重要候選基因進一步分析。共挖掘到3個與酒石酸含量顯著相關的候選基因,其中,定位于Chr.02上的基因Glyma.02G015700和Glyma.02G015800分別編碼延胡索酸酶1(Fumarase 1,FUM1)和延胡索酸酶2(Fumarase 2,FUM2);定位于Chr.13上的基因Glyma.13G077800編碼結節蛋白MTN21/EAMA樣轉運蛋白家族蛋白。此外,挖掘到5個與蘋果酸含量顯著相關的候選基因,其中,定位于Chr.05上的基因Glyma.05G112700編碼鋁激活蘋果酸轉運蛋白12(Aluminum-activated malate transporters 12,ALMT12);定位于Chr.16上的基因Glyma.16G176000和Glyma.16G176100分別編碼UDP-糖基轉移酶88A1(UDP-glucosyl transferase 88A1,UGT88A1)和SPX(SYG1/PHO81/XPR1)結構域蛋白;定位于Chr.19上的基因Glyma.19G188600和Glyma.19G189100分別編碼真核天冬氨酸蛋白酶家族蛋白和泛素蛋白連接酶7(Ubiquitin-protein ligase 7,UPL7)。
3 結論與討論
有機酸作為菜用大豆中重要的風味物質,對菜用大豆風味的形成具有顯著的影響。目前,有機酸含量的檢測通常采用酸堿滴定法、離子色譜法、氣相色譜法和高效液相色譜法等。其中,高效液相色譜法可同時定量測定多種有機酸,且操作簡便、準確度高、穩定性好,因此,被越來越多的利用于有機酸快速檢測領域。本研究對264份菜用大豆種質資源進行有機酸含量測定,2020,2021年供試群體酒石酸含量分別為1.66~9.19 mg/g,0.48~13.52 mg/g,平均含量分別為4.13,4.16 mg/g;2020,2021年菜用大豆自然群體蘋果酸含量分別為1.47~36.46 mg/g,1.89~25.19 mg/g,平均含量分別為7.26,8.99 mg/g;2020,2021年菜用大豆自然群體檸檬酸含量分別為2.56~12.05 mg/g,1.54~18.31 mg/g,平均含量分別為7.12,10.88 mg/g。2021年菜用大豆籽粒中3種有機酸平均含量最高的是檸檬酸,其次是蘋果酸,這與Song等[14],Hu等[2]及張玉梅等[15]的研究結果一致。2 a中3種有機酸變異系數均大于10%,變異分析結果表明,供試菜用大豆種質資源遺傳多樣性豐富,有機酸含量存在非常廣泛的表型變異,具有很大的遺傳改良潛力。在此基礎上篩選出6份高有機酸含量的特異種質資源,其中在2020,2021的檢測中均屬于高酒石酸含量組的種質為阜04-35和桂春豆112;2 a中均屬于高蘋果酸含量組的種質為華春9號和邯12-204;2 a中均屬于高檸檬酸含量組的種質為灌豆3號和新沂大紫花,以上鑒定到的種質為菜用大豆有機酸品種改良育種提供優秀的材料。
基于MLM模型的全基因組關聯分析2 a中共檢測到了54個與酒石酸顯著相關的SNP,189個與蘋果酸顯著相關的SNP以及43個與檸檬酸顯著相關的SNP。但在2 a中3種有機酸均未鑒定到共同顯著關聯位點,這可能是因為有機酸是典型的數量性狀,由多基因控制,含量極易受環境條件等因素的影響,因此,即便使用相同的自然群體,在不同環境條件下檢測到一系列不同的關聯位點的情況也是存在的[16]。此外,Hu等[2]在菜用大豆的籽粒中共鑒定出36種有機酸,其中含量最高的5種有機酸分別為檸檬酸、丁二酸、富馬酸、蘋果酸和草酰乙酸,本研究僅測定了酒石酸、蘋果酸和檸檬酸這3種有機酸,而含量較高的丁二酸、富馬酸還未進行研究,如果丁二酸和富馬酸在年份之間有很大的變動,勢必會引起其他有機酸的變化,從而會導致共定位的難度增大。因此,后續研究的重點一方面可以增加研究有機酸的種類,另一方面通過多年多點的不同環境試驗,對菜用大豆自然群體有機酸含量進行檢測,以期獲得更加可靠的顯著關聯SNP位點,使有機酸遺傳標記的開發和候選基因的發掘更加精確。以顯著關聯的SNP位點上下游120 kb范圍內的基因為候選基因,根據基因功能注釋信息,共鑒定到3個與酒石酸含量顯著相關的候選基因及5個與蘋果酸含量顯著相關的候選基因。其中,Glyma.05G112700在擬南芥中的同源基因為AtALMT12,編碼鋁激活蘋果酸轉運蛋白12。在不同物種中均證實了ALMT家族基因能介導植物中蘋果酸的轉運,且參與果實中的蘋果酸含量的調控,在果實酸化過程中發揮著重要作用[17-19],然而,對于該家族基因在菜用大豆中的功能及其調控機制尚不清楚。前人研究表明擬南芥中的AtALMT12調控蘋果酸的運輸,并且參與調節氣孔運動[20],由此推測菜用大豆中的Glyma.05G112700基因可能與其具有相似的功能,與菜用大豆中蘋果酸的含量具有相關性,但其調控機理還需進一步研究。綜上所述,本試驗結果可以為今后研究菜用大豆有機酸的基因位點提供參考依據,同時為候選基因的功能研究奠定基礎,進而為菜用大豆育種提供參考借鑒。
猜你喜歡
農業科技通訊(2023年1期)2023-02-12 07:09:18
今日農業(2022年16期)2022-11-09 23:18:44
中國化肥信息(2022年7期)2022-08-31 01:29:28
中國化肥信息(2022年5期)2022-08-30 01:58:26
今日農業(2021年20期)2021-11-26 01:23:56
今日農業(2021年14期)2021-10-14 08:35:34
下一代英才(酷炫少年)(2018年6期)2018-07-09 03:17:44
農產品市場周刊(2017年4期)2017-03-03 19:40:05
兒童故事畫報·智力大王(2015年10期)2016-01-27 01:01:35
讀寫算(中)(2015年10期)2015-11-07 07:24:12
