干旱對不同水稻萌發(fā)種子中逆境生理及相應(yīng)基因表達的影響
2023-09-02 07:25:02擺小蓉閔煒芳石亞飛舍楊夢斐田浩天羅成科
華北農(nóng)學報 2023年4期
關(guān)鍵詞:水稻
擺小蓉,閔煒芳,石亞飛,舍楊夢斐,田浩天,羅成科
(寧夏大學 農(nóng)學院,寧夏 銀川 750021)
我國干旱和半干旱區(qū)約占國土總面積的1/2以上,據(jù)估計干旱造成的危害接近于其他多種自然災(zāi)害影響的總和[1]。在非生物脅迫中,干旱是影響植物生長發(fā)育的主要脅迫因子之一,破壞了植物體內(nèi)的水分平衡,紊亂了植物細胞中的新陳代謝,引起植物一系列表型、生理生化和分子水平的變化,阻礙植物正常生長[2]。水稻(OryzasativaL.)是重要的糧食作物,全世界約115個國家生產(chǎn)水稻,超過半數(shù)以上的人口以稻米為主食。但由于干旱的頻繁發(fā)生,抗旱性差的水稻品種受其影響嚴重減產(chǎn)[3]。種子萌發(fā)是植物體整個生長發(fā)育過程的起始階段,同時也是種子對外界環(huán)境干擾響應(yīng)最敏感的時期[4]。種子萌發(fā)是指種子從吸脹作用開始的一系列有序的生理過程和形態(tài)發(fā)生過程,在萌發(fā)的過程中除了形態(tài)特征變化,還伴隨著相關(guān)酶的變化[2]。水分是種子萌發(fā)期必備的條件,參與種子萌發(fā)的多個生理生化過程。植物通過足夠的水分活化相關(guān)酶,分解種子內(nèi)貯藏的有機物質(zhì),加快新陳代謝,促進種子萌發(fā)[5]。然而,干旱脅迫會延長種子萌發(fā)時間,減緩發(fā)芽速度,降低發(fā)芽率,影響作物萌發(fā)期進程[6]。因此,萌發(fā)種子的抗旱強弱性可從一定程度上反映作物的抗旱性[7]。
逆境脅迫導(dǎo)致植物體內(nèi)活性氧(ROS)大量積累,進而對胞內(nèi)關(guān)鍵酶、蛋白質(zhì)和核酸造成氧化損傷[8-9]。因而,植物需要通過積累滲透調(diào)節(jié)物質(zhì)、激活抗氧化系統(tǒng)來清除大量積累的ROS,維持ROS產(chǎn)生和清除之間的平衡,以此來提高植物的抗逆性。許健等[10]研究發(fā)現(xiàn),干旱脅迫降低了萌發(fā)期玉米種子的發(fā)芽率、發(fā)芽勢,抑制了胚根和胚芽的生長。裴帥帥等[11]研究表明,干旱脅迫下,萌發(fā)期不同谷子品種的相對發(fā)芽勢、相對發(fā)芽率、發(fā)芽指數(shù)和抗旱指數(shù)都呈下降趨勢;SP、MDA、Pro含量,SOD和POD活性都有所升高,各材料間差異較顯著。其中,晉谷45的Pro、SP含量、SOD和POD活性比對照分別提高462.34%,34.17%,113.84%,61.22%。鞠樂等[12]研究發(fā)現(xiàn),隨著干旱脅迫程度的增加,萌發(fā)期大麥種子的SOD活性、MDA、Pro、SS、SP含量變化均呈顯著的上升趨勢,CAT和POD活性的變化基本呈先上升后下降的趨勢。其中,強抗旱性材料Z037P017Q-1的SOD、POD、CAT活性和SS含量顯著高于其余材料。田又升等[13]研究發(fā)現(xiàn),干旱脅迫下,相較于干旱敏感型品種新陸早27號,抗旱型品種新陸早50號的脯氨酸合成基因GhP5CS、過氧化物酶基因GhPOD、過氧化氫酶基因GhCAT的表達水平均明顯增加,但超氧化物歧化酶基因GhSOD表達水平無明顯差異。
前人對萌發(fā)期不同作物抗逆(抗旱、耐鹽、耐冷等)種質(zhì)資源篩選評價進行了大量的研究,而對逆境脅迫下萌發(fā)種子內(nèi)部生理特性的變化研究較少,尤其對干旱脅迫下萌發(fā)的水稻種子內(nèi)部生理特性及其相應(yīng)基因表達水平的變化更是鮮有報道。本研究在前期“種子萌發(fā)期粳稻種質(zhì)資源耐旱性綜合評價”[14]研究基礎(chǔ)上,挑選了3個抗旱性材料(Farry、松粳3號、寧粳36)和3個旱敏感材料(Calrose、京寧10號、山形86),在種子萌發(fā)期進行模擬干旱脅迫(15% PEG-6000)處理,通過比較分析不同萌發(fā)種子生長指標、生理及相應(yīng)基因表達水平的變化,為揭示萌發(fā)期水稻抗旱性機理提供理論依據(jù)。
1 材料和方法
1.1 試驗材料
供試材料為3個旱敏感材料(Calrose、京寧10號、山形86)和3個抗旱性材料(Farry、松粳3號、寧粳36)。
1.2 試驗設(shè)計
針對不同水稻材料,分別選取籽粒飽滿、大小一致的種子,經(jīng)15%次氯酸鈉溶液消毒30 min,蒸餾水沖洗數(shù)次后,均勻地置于底部鋪有2層無菌濾紙的培養(yǎng)皿中,置于人工氣候箱中萌發(fā),培養(yǎng)條件為12 h/12 h(光照/黑暗),溫度28 ℃/26 ℃(白天/夜間)。設(shè)置處理如下:蒸餾水做對照(CK),15% PEG-6000模擬干旱脅迫處理,試驗過程中及時補充蒸餾水或PEG-6000 溶液,連續(xù)處理7 d(DS)。在種子萌發(fā)期第1,3,5,7天取樣,萌發(fā)指標測定樣品為萌發(fā)種子,生理指標和基因表達量測定樣品為去除胚根與胚芽的種子。每個處理3個重復(fù),每個重復(fù)50粒。
1.3 測定項目與方法
1.3.1 萌發(fā)指標的測定 發(fā)芽標準為胚根突破種皮1 mm、胚芽為種子長度1/2[15]。每日定時測定發(fā)芽種子數(shù),計算相對發(fā)芽勢、相對芽長、萌發(fā)抗旱指數(shù)和活力指數(shù),具體公式詳見參考文獻[14]。
1.3.3 總RNA提取、cDNA合成和RT-qPCR分析 使用TaKaRa MiniBEST Plant RNA Extraction Kit(TaKaRa) 對2種處理(CK、DS)下的不同抗旱性水稻種子進行 RNA 提取。RNA提取液使用NanoDrop 2000/2000c(Gene Company Limited)分光光度計檢測純度和濃度。選取A260/280比值為1.8~2.2且A260/230比值約為2.0的樣品進行后續(xù)分析。使用PrimeScriptTMRT reagent Kit with gDNA Eraser(TaKaRa)去除基因組DNA污染并反轉(zhuǎn)錄獲得cDNA。根據(jù)Rice Genome Annotation Project(http://rice.uga.edu/)中的序列,采用Primer Premier 5進行 RT-qPCR 引物設(shè)計,引物信息參見表1。根據(jù) TaKaRa 公司的 TB Green?PremixExTaqTMⅡ試劑使用說明書,用qTOWER3G(Analytikjena,Germany)進行 RT-qPCR 試驗。每個樣品進行3次重復(fù)。RT-qPCR 結(jié)果采用 2-ΔΔCt方法計算各基因的相對表達量[17]。

表1 檢測的基因和RT-qPCR引物序列
1.4 數(shù)據(jù)統(tǒng)計與分析
采用Microsoft Excel 2016軟件整理數(shù)據(jù),SPSS 21.0軟件進行統(tǒng)計分析,差異顯著性定義為P<0.05,用平均值±標準誤表示3次重復(fù)測定結(jié)果,采用Origin 2021 b與GraphPad Prism 8作圖。
2 結(jié)果與分析
2.1 干旱脅迫下萌發(fā)期不同水稻生長指標的影響
由表2可知,正常條件下,3種抗旱性材料之間,發(fā)芽勢、芽長、活力指數(shù)和萌發(fā)指數(shù)均無顯著差異;而旱敏感材料之間則表現(xiàn)為,除芽長外,3種旱敏感材料的發(fā)芽勢、活力指數(shù)和萌發(fā)指數(shù)均無顯著差異。在不同類型材料之間相比,除活力指數(shù)外,抗旱性材料Farry、松粳3號和寧粳36的發(fā)芽勢、芽長和萌發(fā)指數(shù)顯著高于旱敏感材料Calrose、京寧10號和山形86。
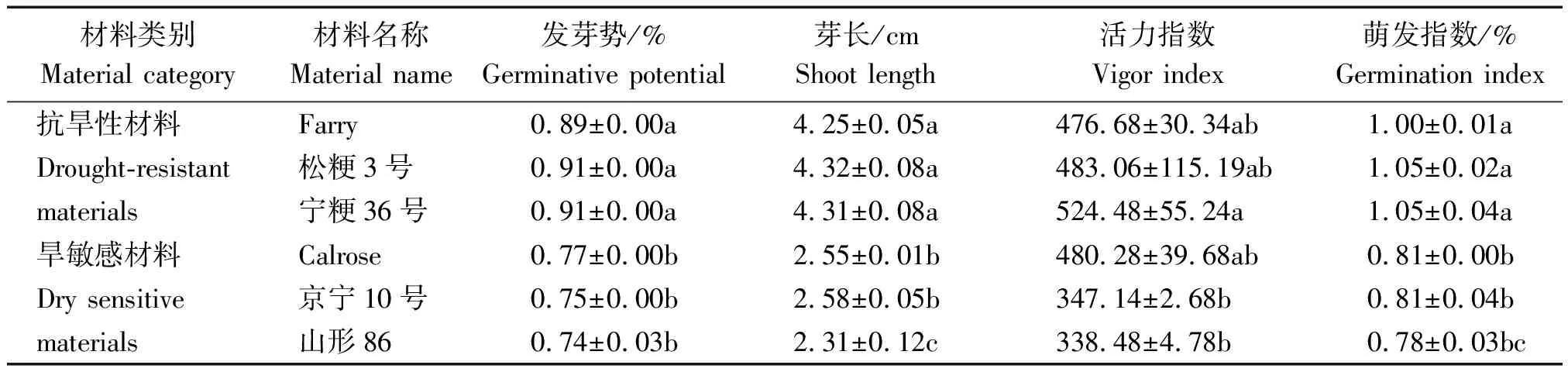
表2 正常條件下萌發(fā)期水稻的生長指標
進一步利用相對發(fā)芽勢、相對芽長、萌發(fā)抗旱指數(shù)和活力指數(shù)分析干旱脅迫對水稻種子萌發(fā)的影響,結(jié)果表明,對抗旱性材料而言,除萌發(fā)抗旱指數(shù)表現(xiàn)出寧粳36顯著高于其他2個抗旱性材料外,其余指標在3個材料間無顯著差異;對于旱敏感材料,相對芽長和萌發(fā)抗旱指數(shù)在3個旱敏感材料間無顯著差異,但活力指數(shù)表現(xiàn)出Calrose和京寧10號顯著高于山形86。不同基因型水稻間相比,抗旱性材料寧粳36的相對發(fā)芽勢顯著高于旱敏感材料(Calrose、京寧10號和山形86),抗旱性材料(Farry、松粳3號)的相對發(fā)芽勢顯著高于旱敏感材料山形86;其余3個指標(相對芽長、萌發(fā)抗旱指數(shù)、活力指數(shù))的變化趨勢與相對發(fā)芽勢的變化趨勢基本一致(圖1)。其中,相比于旱敏感性材料,抗旱性材料萌發(fā)種子的相對發(fā)芽勢(RGP)、相對芽長(RSL)、萌發(fā)期抗旱指數(shù)(GDRI)和活力指數(shù)(VI)分別增加了0.03~0.07百分點,0.32~0.39百分點,0.12~0.18百分點和92.41%~108.39%。表明相對于旱敏感材料,抗旱性材料的種子發(fā)芽速率更快,種子活力更高,抗旱性更強。

不同小寫字母表示不同材料間0.05水平差異顯著(P<0.05)。
2.2 干旱脅迫對萌發(fā)期不同水稻丙二醛和活性氧含量的影響


不同小寫字母表示各處理下不同品種間0.05水平差異顯著。圖3—4同。
2.3 干旱脅迫對萌發(fā)期不同水稻抗氧化酶活性及滲透調(diào)節(jié)物質(zhì)含量的影響
由圖3-A—C可知,正常條件下在各萌發(fā)時間點,除萌發(fā)7 d種子中的POD活性外,萌發(fā)種子中的SOD、POD活性(以鮮質(zhì)量計)在不同抗旱性材料之間無顯著差異。相較于正常條件,干旱脅迫下各萌發(fā)時間點不同抗旱性水稻萌發(fā)種子中的SOD和POD活性均明顯增加。其中,抗旱性材料寧粳36的SOD活性顯著高于旱敏感材料(Calrose、京寧10號)。由圖3-C可知,對CAT活性(以鮮質(zhì)量計)來說,無論正常條件還是干旱脅迫,不同抗旱性水稻萌發(fā)種子中其活性隨著萌發(fā)時間的延長逐漸增強。相較于正常條件,干旱脅迫下不同抗旱性水稻萌發(fā)種子中的CAT活性均有增加,其中,抗旱性材料Farry、松粳3號和寧粳36中的CAT活性明顯高于旱敏感材料Calrose和京寧10號。其中,相比于旱敏感性材料,抗旱性材料的抗氧化酶(SOD、POD、CAT)活性分別提高17.29%~33.12%,15.24%~76.06%和14.68%~18.61%。

圖3 干旱脅迫對萌發(fā)期不同水稻抗氧化酶活性及滲透調(diào)節(jié)物質(zhì)含量的影響
由圖3-D—F可知,正常條件下萌發(fā)1~3 d內(nèi),抗旱性材料Farry、松粳3號和寧粳36萌發(fā)種子中的Pro、SP含量(以鮮質(zhì)量計)明顯高于旱敏感材料Calrose、京寧10號和山形86。萌發(fā)5~7 d內(nèi),抗旱性材料寧粳36萌發(fā)種子中的這2種滲透調(diào)節(jié)物質(zhì)含量明顯高于其他3種材料。除萌發(fā)3 d,旱敏感性材料山形86的SS含量顯著低于抗旱性材料(Farry、松粳3號和寧粳36)外,其余時期SS含量在不同抗旱性材料之間均沒有顯著差異。相比于正常條件,在整個萌發(fā)期內(nèi),干旱脅迫明顯增加了不同抗旱性材料萌發(fā)種子中的Pro含量,但并沒有明顯增加SP和SS含量。相比于旱敏感材料Calrose、京寧10號和山形86,萌發(fā)第1~3天內(nèi)抗旱性材料Farry、松粳3號和寧粳36萌發(fā)種子中的Pro含量顯著增加,萌發(fā)第5天只有寧粳36的Pro含量顯著增加。SP含量明顯增加則表現(xiàn)在萌發(fā)第3,7天的寧粳36種子中。對于SS含量來說,萌發(fā)第3天的Farry和松粳3號種子顯著高于旱敏感材料山形86,萌發(fā)第5~7天3種抗旱性材料均顯著高于旱敏感材料。其中,相比于旱敏感性材料,抗旱性材料的滲透調(diào)節(jié)物質(zhì)(Pro、SS、SP)含量分別增加了5.93%~18.29%,1.08%~7.97%和3.47%~6.03%。
2.4 干旱脅迫對萌發(fā)期不同水稻抗氧化物酶合成基因及脯氨酸合成基因相對表達量的影響
OsALM1、OsPOX1、OsCATC和OsP5CS是編碼抗氧化酶SOD、POD、CAT和脯氨酸Pro合成基因。如圖4所示,正常條件下,水稻萌發(fā)種子中的OsALM1、OsCATC、OsPOX1和OsP5CS基因表達量在不同抗旱性材料之間無顯著差異。相較于正常條件,干旱脅迫不同程度地誘導(dǎo)了不同抗旱性水稻萌發(fā)種子中這4個基因的相對表達水平。其中,抗旱性材料(Farry、松粳3號和寧粳36)萌發(fā)種子中ALM1和CATC基因相對表達量明顯高于旱敏感材料(Calrose、京寧10號和山形86),基因表達量分別增加了2.81,2.90,2.08倍。對于OsPOX1和OsP5CS基因相對表達量變化來說,它們僅在寧粳36的表現(xiàn)顯著高于旱敏感材料Calrose、京寧10號和山形86,基因表達量分別增加了1.97,1.96,2.03倍。其中,相比于旱敏感性材料,抗旱性材料的脯氨酸合成關(guān)鍵基因OsP5CS、抗氧化酶合成基因(OsALM1、OsPOX1、OsCATC)的相對表達量分別上調(diào)了2.66%~182.31%,57.14%~513.27%,0.38%~109.06%和63.39%~184.25%。

圖4 干旱脅迫對萌發(fā)期不同水稻抗氧化物酶合成基因及脯氨酸合成基因相對表達量的影響
2.5 干旱脅迫下萌發(fā)期不同水稻中各指標間的相關(guān)性分析

CAT.過氧化氫酶;SOD.超氧化物歧化酶;POD.過氧化物酶;SS.可溶性糖;MDA.丙二醛;SP.可溶性蛋白;Pro.脯氨酸;H2O2.過氧化氫;超氧陰離子;RGP.相對發(fā)芽勢;RSL.相對芽長;GDRI.萌發(fā)抗旱指數(shù);VI.活力指數(shù);OsP5CS.脯氨酸合成基因;OsPOX1.過氧化物酶基因;OsCATC.過氧化氫酶基因;OsALM1.超氧化物歧化酶基因。*.在P<0.05水平差異顯著,圖6同。
2.6 干旱脅迫下萌發(fā)期不同水稻中各指標間主成分分析及各指標參數(shù)的選擇
在相關(guān)性分析的基礎(chǔ)上,進一步對干旱脅迫下萌發(fā)期不同水稻中的以上17個指標進行了主成分分析,根據(jù)指標選擇的原則,提取3個主成分進行分析。對抗旱性材料(Farry、松粳3號、寧粳36)來說,第1主成分包括OsPOX1、OsP5CS、OsCATC,第2主成分包括VI、POD,第3主成分為SP(圖6-A—C),本研究認為OsPOX1、OsP5CS、OsCATC、VI、POD、SP是影響抗旱性材料的關(guān)鍵指標;對旱敏感材料(Calrose、京寧10號、山形86)而言,第1主成分包括VI、RSL、OsPOX1,第2主成分包括OsALM1、POD,第3主成分為H2O2(圖6-D—F),本研究認為VI、RSL、OsPOX1、OsALM1、POD、H2O2是影響旱敏感材料的關(guān)鍵指標。

圖6 干旱脅迫下萌發(fā)期不同水稻中各指標間的主成分分析
3 結(jié)論與討論
3.1 干旱脅迫對不同水稻種子萌發(fā)的影響
水分是限制種子萌發(fā)的主要環(huán)境因子。種子萌發(fā)期極易受水分的影響,一旦水分短缺,將會導(dǎo)致種子萌發(fā)困難,發(fā)芽率下降。在進行作物種子萌發(fā)期抗旱性鑒定時,常采用相對發(fā)芽勢、相對芽長、活力指數(shù)和萌發(fā)抗旱指數(shù)等指標來評價作物不同種質(zhì)資源的抗旱性強弱[18]。例如,在評價萌發(fā)期不同黑小麥[19]、不同燕麥[20]抗旱性時,這些作物的抗旱性材料通常長勢較好,生長指標受脅迫抑制小,而旱敏感性材料長勢較差,生長指標受脅迫抑制大。本研究結(jié)果也表明,干旱脅迫抑制了2種基因型水稻種子發(fā)芽勢、芽長和種子活力。相較于旱敏感性材料(Calrose、京寧10號和山形86),抗旱性材料(Farry、松粳3號和寧粳36)的相對發(fā)芽勢、相對芽長、活力指數(shù)以及萌發(fā)抗旱指數(shù)均明顯增加,說明抗旱性材料的生長指標受干旱脅迫影響較小,其對干旱脅迫的適應(yīng)性更強。
3.2 干旱脅迫對不同水稻萌發(fā)種子內(nèi)生理特性的影響
逆境脅迫下,植物能夠啟動抗氧化防御系統(tǒng)(抗氧化酶、非酶類抗氧化劑)來清除體內(nèi)過多的ROS,其中抗氧化酶被認為是清除ROS最直接、最有效的途徑[27]。段驊等[28]研究發(fā)現(xiàn),相較于正常條件,干旱脅迫下,萌發(fā)期不同抗旱性水稻的SOD、POD、CAT活性均明顯增加。其中,抗旱性品種黃華占的抗氧化酶活性顯著高于旱敏感性品種雙桂1號。另外,萌發(fā)期不同水稻抗旱性的其他研究報道也證明了抗旱性材料中的各種抗氧化酶活性明顯高于旱敏感性材料[29]。本研究結(jié)果表明,干旱脅迫均明顯增加了各萌發(fā)時間點不同水稻萌發(fā)種子中的SOD、POD、CAT活性。其中,3個抗旱性材料(Farry、松粳3號和寧粳36)中SOD和POD活性均顯著高于3個旱敏感材料(Calrose、京寧10號和山形86),3個抗旱性材料(Farry、松粳3號和寧粳36)中CAT活性明顯高于2個旱敏感性材料(Calrose和京寧10號),說明萌發(fā)期抗旱性強的水稻材料可以通過合成更多的抗氧化酶來清除過量的ROS,進而增強其抗旱性。
植物還可以通過積累滲透調(diào)節(jié)物質(zhì)(Pro、SP、SS)來保護自身免受非生物脅迫損傷[16]。在干旱脅迫下,滲透調(diào)節(jié)物質(zhì)含量的增加有利于維持胞質(zhì)膨壓,其中,脯氨酸還對ROS有直接的清除作用[30]。趙振寧等[31]研究發(fā)現(xiàn),干旱脅迫下不同大豆萌發(fā)種子胚根中滲透調(diào)節(jié)物質(zhì)含量相比正常條件均有增加,其中,抗旱性材料中Pro、SS、SP含量顯著高于旱敏感性材料。本研究結(jié)果也表明,相比于正常條件,干旱脅迫明顯增加了不同水稻萌發(fā)種子中3種滲透調(diào)節(jié)物質(zhì)的含量。其中,萌發(fā)期抗旱性材料寧粳36中SP含量明顯高于3個旱敏感材料(Calrose、京寧10號和山形86),萌發(fā)前期1~3 d抗旱性材料Farry和松粳3號中Pro和SS含量、寧粳36中Pro含量均明顯高于3個旱敏感材料,而萌發(fā)后期5~7 d抗旱性材料寧粳36中Pro和SS含量、Farry和松粳3號中SS含量明顯高于3個旱敏感材料,說明抗旱性材料通過積累更多的滲透調(diào)節(jié)物質(zhì)來提高自身抗旱性。
3.3 干旱脅迫對不同水稻萌發(fā)種子內(nèi)抗氧化酶與脯氨酸合成基因表達量的影響
植物抗旱機制十分復(fù)雜,受到各種生理生化過程和基因表達水平的綜合調(diào)控。其中,抗氧化酶活性和滲透調(diào)節(jié)物質(zhì)積累受相應(yīng)基因的表達控制,如OsALM1、OsPOX1、OsCATC分別是SOD、POD、CAT抗氧化酶合成基因,OsP5CS是滲透調(diào)節(jié)物質(zhì)脯氨酸合成基因。Luo等[16]研究表明,相較于正常條件,干旱脅迫顯著上調(diào)了抗氧化酶基因OsALM1、OsPOX1、OsCATC和脯氨酸合成基因OsP5CS的表達水平。與單獨干旱脅迫相比,褪黑素預(yù)處理更加明顯地促進了干旱脅迫水稻中這些基因的表達水平。Challabathula等[32]研究發(fā)現(xiàn),在干旱脅迫條件下,抗旱品種Vaisakh中編碼Cu/Zn SOD、CAT和APX酶基因的表達量明顯高于旱敏感品種Aiswarya,說明這些抗氧化酶為抗旱品種抵御ROS毒害提供了先天保護。本研究表明,干旱脅迫明顯誘導(dǎo)了萌發(fā)期不同水稻中抗氧化酶基因(OsALM1、OsPOX1、OsCATC)和脯氨酸合成基因OsP5CS的表達水平。其中,抗旱性材料(Farry、松粳3號和寧粳36)中OsALM1和OsCATC基因表達量的上調(diào)程度均明顯大于旱敏感材料(Calrose、京寧10號和山形86),而對于OsPOX1和OsP5CS基因表達量的上調(diào)程度而言,抗旱性材料寧粳36明顯高于3個旱敏感材料。說明抗旱性材料通過上調(diào)抗氧化酶合成基因和脯氨酸合成基因的表達水平,激活植物抗氧化防御系統(tǒng)和增強滲透調(diào)節(jié)能力,從而增強萌發(fā)期水稻的抗旱性。
3.4 萌發(fā)期不同水稻抗旱性關(guān)鍵指標選擇
萌發(fā)期作物抗旱性是一個復(fù)雜的數(shù)量遺傳性狀。萌發(fā)期作物在干旱脅迫下會調(diào)節(jié)體內(nèi)生理生化反應(yīng),并能激活抗旱相關(guān)基因表達,從而做出對干旱脅迫的響應(yīng)和適應(yīng)。在評價萌發(fā)期作物抗旱性的過程中,選擇單一或少數(shù)幾個指標存在一定的片面性,為了做出全面、科學的評價,需要從形態(tài)、生理及相應(yīng)基因表達量等多個層次選擇抗旱性相關(guān)指標[33]。例如,范蓉[34]在綜合分析苗期不同抗旱性棉花材料時,選擇5個抗逆生理指標(MDA、Pro含量,SOD、POD活性和電導(dǎo)率)和4個基因表達量(GhHSP70-26、GhHsfA7、GhbZIP15和GhNAC2)共9個指標進行相關(guān)性分析,分析結(jié)果表明,這些指標與棉花苗期干旱脅迫應(yīng)答反應(yīng)密切相關(guān),其中,SOD、POD、Pro、GhHSP70-26、GhHsfA7、GhbZIP15、GhNAC2這7個指標是影響棉花幼苗抗旱性的關(guān)鍵指標。本試驗對干旱脅迫下萌發(fā)期不同水稻中的生長指標、生理指標及其相應(yīng)基因表達量共17個指標進行相關(guān)性和主成分分析,篩選獲得了萌發(fā)期抗旱性水稻的關(guān)鍵指標是OsPOX1、OsP5CS、OsCATC、VI、POD、SP,萌發(fā)期旱敏感水稻的關(guān)鍵指標是VI、RSL、OsPOX1、OsALM1、POD、H2O2,說明不同基因型水稻在響應(yīng)干旱脅迫過程中存在一定的生理和分子特性差異。
干旱脅迫抑制了水稻種子的萌發(fā),影響了萌發(fā)期種子中的生理特性及其相應(yīng)基因的表達。干旱脅迫下,無論是抗旱性材料還是旱敏感性材料,VI、POD以及OsPOX1是影響水稻種子萌發(fā)的關(guān)鍵指標。除以上指標外,SP、OsP5CS、OsCATC是影響抗旱性材料的其他關(guān)鍵指標,RSL、H2O2、OsALM1是影響旱敏感性材料的其他關(guān)鍵指標。
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農(nóng)業(yè)(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農(nóng)業(yè)(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農(nóng)業(yè)(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
