鞘內注射siRNA-P300抑制CaMK Ⅱ磷酸化緩解佐劑性關節炎大鼠疼痛與炎癥的作用機制
2023-09-06 12:49:12談俊高巍巍滕志鵬王廣代震宇李煒
河北醫藥 2023年15期
談俊 高巍巍 滕志鵬 王廣 代震宇 李煒
關節炎導致慢性疼痛甚至致殘,對人們的生活造成嚴重負擔[1]。關節炎的特征是疼痛、關節腫脹、畸形和功能喪失[2]。但不明確炎性疼痛的發生機制,本研究利用完全弗氏佐劑(CFA)誘導炎性疼痛模型[3],探討關節炎性疼痛的機制。P300蛋白是組蛋白乙酰轉移酶(HAT)家族的關鍵成員,在真核生物中參與基因表達的調控。研究表明,慢性壓迫損傷(CCI)引起的神經性疼痛中P300激活增加[4],并依賴于鈣調蛋白依賴性蛋白激酶Ⅱ(CaMK Ⅱ)的磷酸化[5]。研究表明,細胞內Ca2+相關級聯反應引起的受體損傷和敏化可能是神經性疼痛的神經生理機制[6],其中CaMK Ⅱ的表達變化在其中扮演重要作用[7,8]。CaMK Ⅱ參與外周和中樞敏化過程[6,9]。還有研究表明,抑制CaMK Ⅱ可以改善CCI模型動物的疼痛敏化[6]。但是目前仍無關于P300與其依賴的CaMK Ⅱ的磷酸化調節方式在關節炎性痛模型中扮演的角色。本文假設P300通過抑制磷酸化的CaMK Ⅱ可能與關節炎性疼痛和抗炎作用有關。因此,本研究構建CFA誘導關節炎大鼠模型,評價鞘內注射P300的沉默質粒(siRNA-P300)對關節炎大鼠痛覺、炎性反應的影響,并探討CaMK Ⅱ磷酸化的作用和機制。
1 材料與方法
1.1 實驗材料 大鼠來源于重慶醫科大學實驗動物中心。二甲基亞砜(DMSO)、腫瘤壞死因子(TNF)-α、白介素(IL)-1β、IL-6的ELISA試劑盒及CFA購于上海碧云天公司;Von Frey TM動態足底觸覺測量儀購于上海玉研科學儀器有限公司;輻射熱刺痛儀購于四川科儀誠科技有限公司。P300、P物質(SP)、c-fos,p-CaMK Ⅱ的抗體購于美國Abcam公司。
1.2 方法
1.2.1 動物的造模、給藥與分組:3月齡雄性SD大鼠60只,體重181~233 g。實驗期間動物飼養于重慶醫科大學動物房,每日光照約12 h,自由飲食和飲水。所有大鼠飼養2周后進入實驗。將大鼠隨機分為A、B、C、D、E、F共6組(n=10):A組:僅為大鼠左后肢踝關節注射等體積0.9%氯化鈉溶液作為對照組。B組:左后肢踝關節注射完全弗氏佐劑(CFA)誘導佐劑性關節炎模型大鼠;C組:鞘內注射陰性對照(siRNA-NC),其余操作與D組一致;D組:左后肢踝關節注射CFA誘導佐劑性關節炎模型大鼠,制備前10 min鞘內注射用于敲低P300的小干擾RNA(siRNA-P300);E組:鞘內聯合注射siRNA-P300和油酸的溶劑二甲基亞砜(DMSO),其余操作與D組一致;F組:鞘內聯合注射siRNA-P300和油酸(CaMKⅡ的激活劑),其余操作與D組一致。建模后1 d、7 d、14 d用電子千分卡尺測左后足跖厚度。
1.2.2 行為學檢測:①利用Von Frey TM動態足底觸覺測量儀檢測大鼠的機械性縮足閾值(PWMT)。于建模前1 d,建模后1 d、7 d、14 d檢測各組大鼠的疼痛刺激閾值,每次檢測于上午8∶30~11∶30進行,波間隔設置為10,頻率為10.0 Hz,強度1.000 V。測定時保持被檢測肢體溫度在37℃左右。將大鼠放入有機玻璃籠中,底部為1 cm×1 cm網格的鐵絲網,安靜20 min后開始測量。測痛儀的Von Frey纖維絲由下向上刺激大鼠右側后肢足底,設定刺激力度在20 s內由0 g逐漸增強至26 g,當大鼠出現迅速縮足時刺激力度會停止;若未出現縮足反應,壓力值增到26 g后也會自動停止。屏幕顯示的壓力值即為痛閾值,每只大鼠測3次。每2次測量間隔5 min,取其平均值。②另外,使用熱輻射痛檢測儀分別檢測大鼠左后足的熱縮足潛伏期(PWTL),與機械異位痛同時進行測試。將大鼠置于3 mm厚的有機玻璃箱(20 cm×40 cm×35 cm)中,適應30 min環境后確定PWTL。儀器參數設定紅外強度為50,終止時間為20 s。紅外線熱源被放置在腳下并打開。爪子拔出后切斷電源,自動記錄照射時間。每次刺激間隔5 min,以避免潛在的組織損傷。
1.2.3 組織檢測:動物腹腔內注射1%戊巴比妥鈉麻醉,鞘內注射20 μl亞甲藍確認導管位置。開胸灌流沖洗心臟,然后灌流固定40~60 min。大鼠在造模-1 d、1 d、7 d、14 d時取材制成切片。暴露脊柱棘突和橫突,選L4和L5作入路。剪刀剪去部分棘突形成“V”形切口,暴露黃韌帶和脊髓組織。咬去黃韌帶和硬脊膜,繼續咬去兩側的橫突。鉗下骨質暴露脊髓并取出,剪去脊神經節兩端的脊神經干。將脊髓浸泡在4%多聚甲醛固定36 h,在30%蔗糖中脫水24 h。用O.C.T.進行包埋,在-20℃下切6 μm厚手動切片。吸附在載玻片上保存。用免疫熒光方法檢測定位定性P300在脊髓中的表達。冷凍切片經過血清封閉并于室溫回溫,孵育兔抗大鼠的P300的一抗(1∶300)于濕盒中4℃過夜,復溫30 min,隨后避光下孵育山羊抗兔IgG二抗(黃紅色DyLight 549,1∶200)1 h。使用甘油:0.01 mmol/L(1∶1)封片。熒光顯微鏡下拍照保存。
1.2.4 Western blot檢測大鼠神轉錄因子和TRP、鈉通道蛋白:冰浴中勻漿處理脊髓組織,冰上孵育20 min,隨后離心15~20 min,收集上清,用酶標儀測定570 nm處的吸光值;計算樣品中的蛋白濃度。行SDS-PAGE電泳(先45 V,后90 V),冰浴電泳1.5 h;使用NC膜進行轉膜。使用Western Blot封閉液封閉用于檢測的NC膜,室溫封閉1 h,孵育SP(1∶500)、c-fos(1∶500)、磷酸化的CaMKⅡ(1∶800)、P300(1∶800)的一抗,室溫孵育30 min后,放于4℃過夜;孵育二抗,室溫孵育40 min,最后進行顯色和曝光。
1.2.5 ELISA檢測血清中TNF-α、L-1β、IL-6的表達:收集14 d大鼠血清。ELISA試劑盒檢測TNF-α、L-1β、IL-6的表達水平。酶標儀檢測A450的光密度。
1.3 觀察指標 6組大鼠機械痛閾值,熱縮足潛伏期,左后足跖厚度,脊髓SP、c-fos、P300、p-CaMK Ⅱ、CaMK Ⅱ的蛋白表達水平,血清中TNF-α、L-1β、IL-6的含量。
2 結果
2.1 大鼠機械痛閾值變化 動物行為學觀察大鼠的MWT。與A組比較,B組造模后1 d、7 d、14 d的MWT值明顯下調,差異均有統計學意義(P<0.05);與B組比較,C組在造模后1 d、7 d、14 d的MWT的變化差異無統計學意義(均P>0.05);與C組比較,D組在造模后1 d、7 d、14 d的MWT均上調,差異均有統計學意義(P<0.05)。與E組比較,F組在造模后1 d、7 d、14 d的MWT均下調,差異均有統計學意義(均P<0.05);與D組比較,E組在造模后1 d、7 d、14 d 的MWT差異無統計學意義(均P>0.05)。見表1。
表1 6組大鼠機械痛閾值 n=10,MWT/g,
2.2 大鼠熱縮足潛伏期變化 動物行為學觀察大鼠的PWTL。與A組比較,B組造模后1 d、7 d、14 d的PWTL明顯縮短,差異均有統計學意義(P<0.05);與B組比較,C組在造模后1 d、7 d、14 d 的PWTL變化差異無統計學意義(均P>0.05);與C組比較,D組在造模后1 d、7 d、14 d 的PWTL均增加,差異均有統計學意義(均P<0.05)。與E組比較,F組在造模后1 d、7 d、14 d 的PWTL均縮短,差異均有統計學意義(均P<0.05);與D組比較,E組在造模后1 d、7 d、14 d 的PWTL差異無統計學意義(均P>0.05)。見表2。
表2 6組大鼠輻射熱潛伏期 n=10,PWIL/S,
2.3 大鼠脊髓SP和c-fos的變化 檢測大鼠造模后14 d脊髓的SP和c-fos的表達變化。A組SP和c-fos的表達水平明顯上調,差異均有統計學意義(P<0.05);與B組比較,C組SP和c-fos的表達差異無統計學意義(P>0.05);與C組比較,D組SP和c-fos的表達水平下調,差異均有統計學意義(P<0.05)。與E組比較,F組SP和c-fos的表達水平上調,差異均有統計學意義(均P<0.05);但與D組比較,E組SP和c-fos的水平差異無統計學意義(均P>0.05)。見表3。
表3 6組大鼠脊髓SP和c-fos的相對表達 n=10,
2.4 大鼠左后足跖腫脹程度變化 測量大鼠左后足跖厚度變化。與A組比較,B組造模后1 d、7 d、14 d的足跖厚度增加,差異均有統計學意義(P<0.05);與C組比較,D組在造模后1 d、7 d的足跖厚度減少,差異均有統計學意義(均P<0.05)。見表4。
表4 6組大鼠左后足跖厚度 n=10,cm,
2.5 大鼠血清TNF-α、L-1β、IL-6的變化 檢測大鼠造模后14 d血清TNF-α、L-1β、IL-6的變化。與A組比較,B組造模后TNF-α、L-1β、IL-6的水平上調,差異均有統計學意義(P<0.05);與B組比較,C組TNF-α、L-1β、IL-6的水平變化差異無統計學意義(P>0.05);與C組比較,D組TNF-α、L-1β、IL-6 的水平下調,差異均有統計學意義(P<0.05)。與E組比較,F組TNF-α、L-1β、IL-6上調,差異均有統計學意義(P<0.05);與D組比較,E組TNF-α、L-1β、IL-6的水平差異無統計學意義(均P>0.05)。見表5。
表5 大鼠血清炎性因子水平 n=10,
2.6 大鼠脊髓CaMK Ⅱ的磷酸化水平變化
2.6.1 免疫熒光結果顯示:與A組比較,B組P300的表達水平上調,差異有統計學意義(P<0.05);與B組比較,C組P300水平變化差異無統計學意義(P>0.05);與C組比較,D組P300的水平下調,差異有統計學意義(P<0.05)。與E組比較,F組的P300上調,差異有統計學意義(P<0.05);與D組比較,E組的P300的水平無統計學意義(P>0.05)。
2.6.2 蛋白免疫印跡檢測大鼠造模后14 d脊髓p-CaMK Ⅱ的表達變化:A組的p-CaMK Ⅱ的表達水平明顯上調,差異有統計學意義(P<0.05);與B組比較,C組p-CaMK Ⅱ表達差異無統計學意義(P>0.05);而與C組比較,D組的p-CaMK Ⅱ的表達水平下調,差異有統計學意義(P<0.05)。與E組比較,F組的p-CaMKII表達水平上調,差異有統計學意義(P<0.05);但與D組比較,E組的p-CaMK Ⅱ的水平無統計學意義(P>0.05)。見圖1、2,表6。
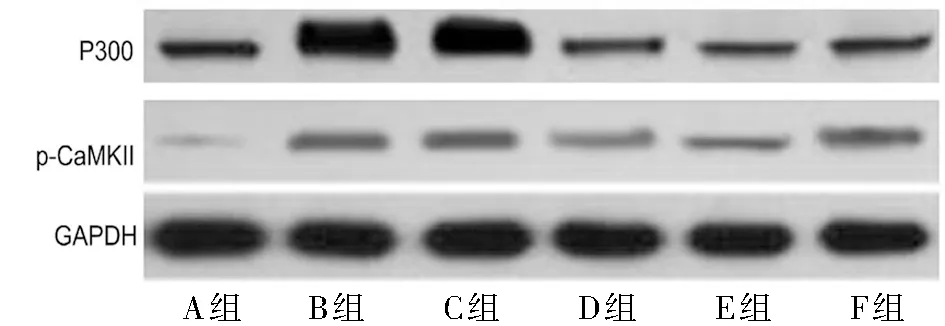
圖1 蛋白免疫印跡檢測大鼠脊髓的P300和p-CaMK Ⅱ的表達

A組 B組 C組
表6 大鼠P300蛋白和CaMKII的磷酸化的相對表達水平 n=10,
3 討論
關節炎是目前導致嚴重威脅人類健康的關節疾病,每年關節炎的新發病例數和致殘總數都在增加。關節炎患者會產生不同程度的疼痛,降低生活質量[1]。關節炎的炎性痛機制較為復雜,容易出現異常性疼痛和痛覺過敏[10],由于其病因的多樣性和復雜性,炎性導致的疼痛已成為關節炎機制研究的關鍵[11]。
本研究使用CFA模擬臨床的關節炎性疼痛。觀察到與對照組大鼠比較,CFA誘發的關節炎性疼痛大鼠中的機械性異常性疼痛持續1個月,另外,本研究也觀察到了CFA注射后的慢性熱痛覺過敏現象持續半月。因此本研究著重研究了1 d、7 d、14 d大鼠的關節炎性疼痛機制。我們還觀察到CaMK Ⅱ參與到了佐劑性關節炎大鼠的疼痛和炎性反應。
首先,動物行為學結果顯示,CFA關節炎大鼠表現出明顯疼痛反應,其中機械痛覺過敏的癥狀持續至少28 d,而熱痛覺過敏持續至少維持15 d。因此,本實驗結果表明,CFA誘導下,大鼠關節的促炎反應加劇,不僅可以誘導機械痛覺過敏還可以誘導熱痛覺過敏。這與Avrampou 等[12]的研究一致。其次,內鈣的相關級聯反應導致神經痛覺敏化,而且研究表明CaMK Ⅱ的磷酸化水平升高作為痛覺敏化的關鍵表征之一[13],因此我們研究了CFA關節炎大鼠脊髓中CaMK Ⅱ的磷酸化水平。蛋白免疫印跡結果觀察到,在CFA刺激后的14 d,大鼠脊髓組織中CaMK Ⅱ的磷酸化水平明顯增加。然而,大鼠在術前接受鞘內注射siRNA-P300,可觀察到明顯的磷酸化CaMK Ⅱ水平被抑制的現象,此結果表明,沉默P300抑制了CFA關節炎大鼠脊髓組織中CaMK Ⅱ的磷酸化水平。
作為HAT家族成員,P300蛋白在以往被證實可以參與CCI神經性痛覺過敏,我們的結果證實,P300在CFA模型大鼠脊髓中表達顯著增加,表明P300可能對CFA疼痛的機制具有潛在的調節作用。另外,P300激活后可以引起CaMK Ⅱ的磷酸化[4,14,15]。在外周和中樞敏化過程中CaMK Ⅱ的磷酸化扮演重要作用[9]。研究表明,CCI神經性痛覺過敏大鼠在造模前接受鞘內注射CaMK Ⅱ抑制劑m-AIP可導致CaMK Ⅱ的磷酸化被抑制,因此表現出痛覺過敏行為減輕的現象,維持了7 d[6]。本研究在大鼠脊髓中利用P300的siRNA成功降低P300的表達。觀察到與CaMK Ⅱ抑制劑m-AIP類似的結果;但是P300被沉默后,我們還觀察到14 d痛覺過敏行被減輕的效果,觀察到脊髓中的疼痛標志物SP和c-fos表達同樣被抑制;而在P300沉默的基礎上繼續使用CaMK Ⅱ激活劑油酸后,痛覺過敏行為反而部分增強,且SP和c-fos表達部分上調,但是油酸對P300的表達未產生顯著影響。因此本研究結果表明,在脊髓中沉默P300可能通過抑制CaMKII的磷酸化抑制CFA大鼠的炎性痛覺過敏。
另外,與對照組比較,CFA組大鼠的足跖厚度增加,而預先注射siRNA-P300,CFA大鼠的足跖厚度在7 d內被明顯抑制,表明siRNA-P300可以抑制關節炎大鼠的關節水腫。我們還檢測大鼠血清TNF-α、L-1β、IL-6的變化。本研究結果顯示siRNA-P300降低了TNF-α、L-1β、IL-6水平,然而注射油酸后,大鼠血清TNF-α、L-1β、IL-6的水平都增加。因此,本研究證明關節大鼠鞘內注射siRNA-P300有抗炎作用,抑制CaMKII的磷酸化是siRNA-P300抗炎作用的關鍵機制之一。
本研究深入探討了關節炎性疼痛的機制,并觀察到脊髓SP、c-fos、P300蛋白、CaMKⅡ的磷酸化水平表達均增加,與觀察到的CFA大鼠疼痛行為相對同步,這表明CaMK Ⅱ可能介入與疼痛有關信號通路的調節。研究表明,大鼠中皮下注射甲醛后,CaMK Ⅱ磷酸化在脊髓背角顯著增加,時間與發炎性疼痛出現的時間高度一致[16]。這進一步證實CaMK Ⅱ可作為炎性痛的標志物或治療靶點。另有相關機制研究證實,CaMK Ⅱ經歷自身磷酸化并激活天冬氨酸NMDA受體,以促進痛覺過敏的持續形成[17,18]。以上這些結果表明,在臨床治療關節炎性疼痛中,連續給CaMK Ⅱ磷酸化的抑制劑可能更有效,而沉默P300對CaMK Ⅱ磷酸化的抑制作用更明顯,因而可作為新的關節炎性疼痛的治療靶點。
綜上所述, P300介導的CaMK Ⅱ磷酸化在關節炎性疼痛和炎性反應中起著重要作用。通過使用siRNA-P300抑制CaMK Ⅱ的磷酸化,可在一定的時間內有效改善大鼠的疼痛行為和炎性反應,為關節炎性疼痛的臨床治療提供了新思路。
猜你喜歡
新少年(2022年9期)2022-09-17 07:10:54
音樂探索(2022年2期)2022-05-30 21:01:37
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
小天使·一年級語數英綜合(2020年6期)2020-12-16 02:56:41
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
人大建設(2019年12期)2019-05-21 02:55:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
北極光(2014年8期)2015-03-30 02:50:51
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
中國火炬(2010年8期)2010-07-25 11:34:30
