基于COI基因分析葉爾羌河5種鯉科魚(yú)類遺傳多樣性
2023-09-06 11:16:16張小雨靳文慧王玉濤
安徽農(nóng)業(yè)科學(xué) 2023年16期
張小雨,靳文慧,王 佳,王玉濤
(喀什大學(xué)生命與地理科學(xué)學(xué)院/新疆帕米爾高原生物資源與生態(tài)重點(diǎn)實(shí)驗(yàn)室,新疆喀什 844000)
葉爾羌河位于新疆塔里木盆地的西南面,起源于世界第二大高峰——喬戈里峰,屬于塔里木河水系[1-2]。流域內(nèi)分布的魚(yú)類主要為鯉形目的鯉科和鰍科,土著魚(yú)以高原鰍屬和裂腹魚(yú)屬為主,普通經(jīng)濟(jì)魚(yú)類由鯉魚(yú)、草魚(yú)、鯽魚(yú)等構(gòu)成[3]。目前,與葉爾羌河流域相關(guān)的研究日益增多,主要集中在水利水電工程[4]、環(huán)境科學(xué)和資源利用[5]等方面,但從分子水平對(duì)該流域魚(yú)類資源狀況的研究鮮見(jiàn)報(bào)道。因此選取適宜的方法對(duì)該流域魚(yú)類資源進(jìn)行鑒定及進(jìn)一步的系統(tǒng)進(jìn)化分析極為重要。
王德忠[6]、楊天燕等[7]、陳生熬等[8]均通過(guò)形態(tài)學(xué)的方法分別分析了新疆裂腹魚(yú)的分布特性和遺傳差異以及葉爾羌高原鰍的種群結(jié)構(gòu)。李國(guó)剛等[9]也曾對(duì)塔里木河等水系的土著魚(yú)類資源展開(kāi)調(diào)查,發(fā)現(xiàn)了水系的特有魚(yú)類。隨著種質(zhì)資源鑒定技術(shù)的進(jìn)一步發(fā)展[10]。其中,COI(線粒體細(xì)胞色素C氧化酶亞基 Ⅰ )基因的引物通用性和進(jìn)化速度[11]均處于較高水平,而且它的5′端序列的種間差異顯著高于種內(nèi),所以,mtDNA基因被視為物種鑒定的理想基因條形碼之一[12]。2003年,Hebert等[13]首次提出COI基因可以作為基因條形碼對(duì)物種進(jìn)行有效鑒定的觀點(diǎn)。此后,Gomes等[14]利用COI基因?qū)ε佬蓄愇锓N和魚(yú)類進(jìn)行鑒定,說(shuō)明COI基因作為遺傳標(biāo)記鑒定物種的有效性。閆亞利等[15-17]也選擇COI基因?qū)︳~(yú)類種質(zhì)鑒定進(jìn)行研究。該研究基于COI基因序列對(duì)葉爾羌河流域5種鯉科魚(yú)類的遺傳多樣性進(jìn)行分析,探討COI基因作為分子標(biāo)記對(duì)葉爾羌河魚(yú)類進(jìn)行鑒定的有效性和適應(yīng)性,有助于對(duì)葉爾羌河魚(yú)類資源狀況展開(kāi)調(diào)查,為該流域魚(yú)類資源的合理開(kāi)發(fā)及利用提供參考。
1 材料與方法
1.1 試驗(yàn)材料從塔什庫(kù)爾干縣、麥蓋提縣等地的葉爾羌河流域采集魚(yú)類樣品共50尾。參照《新疆魚(yú)類志》[18]對(duì)采集的魚(yú)類樣本進(jìn)行形態(tài)學(xué)初步鑒定,拍照記錄每個(gè)樣本,并剪取魚(yú)鰭組織浸泡于95%無(wú)水乙醇,分組編號(hào)后保存于-80 ℃冰箱。
1.2 DNA提取、擴(kuò)增及測(cè)序利用傳統(tǒng)的酚-氯仿法,提取葉爾羌河5種魚(yú)類魚(yú)鰭或魚(yú)肉組織的DNA。該研究相關(guān)引物采用魚(yú)類通用引物(FISH-F1:5′-TCAACCAACCACAAAAAGACATTGGCAC-3′;FISH-R1:5′-TAGACTTCTGGGTGGCCAAAGAATCA-3′;FISH-F2:5′-TCGACTAATCATAAAGGATATCGGCAC-3′;FISH-R2:5′-ACTTCAGGGTGACCGAAGAATCAGAA-3′)[19]擴(kuò)增COI片段。PCR反應(yīng)體系為20 μL,包括2×TaqPCR Mix 12 μL、上下游引物(10 mmol/L)各0.4 μL、DNA模板1 μL、ddH2O加至20 μL;PCR擴(kuò)增程序:94 ℃預(yù)變性2 min,94 ℃變性30 s,58 ℃復(fù)性30 s,72 ℃延伸1 min,以上步驟經(jīng)35個(gè)循環(huán)之后,再72 ℃延伸5 min。PCR擴(kuò)增產(chǎn)物通過(guò)1%瓊脂糖凝膠電泳進(jìn)行檢測(cè),合格后送樣測(cè)序。
1.3 數(shù)據(jù)處理使用Chromas軟件查看正反雙向測(cè)序結(jié)果峰圖,然后使用DNAStar軟件包進(jìn)行序列的拼接、比對(duì)和校對(duì);利用DnaSP 5.10軟件計(jì)算葉爾羌河5種魚(yú)類群體單倍型,并統(tǒng)計(jì)變異位點(diǎn)數(shù)、單倍型多樣性指數(shù)(Hd)、核苷酸多樣性指數(shù)(Pi)、核苷酸差異數(shù)(K)等;采用MEGA 7.0軟件計(jì)算序列核苷酸組成和遺傳距離等,并結(jié)合下載的12條同源序列(具體信息見(jiàn)表1)基于NJ法構(gòu)建系統(tǒng)進(jìn)化樹(shù)。
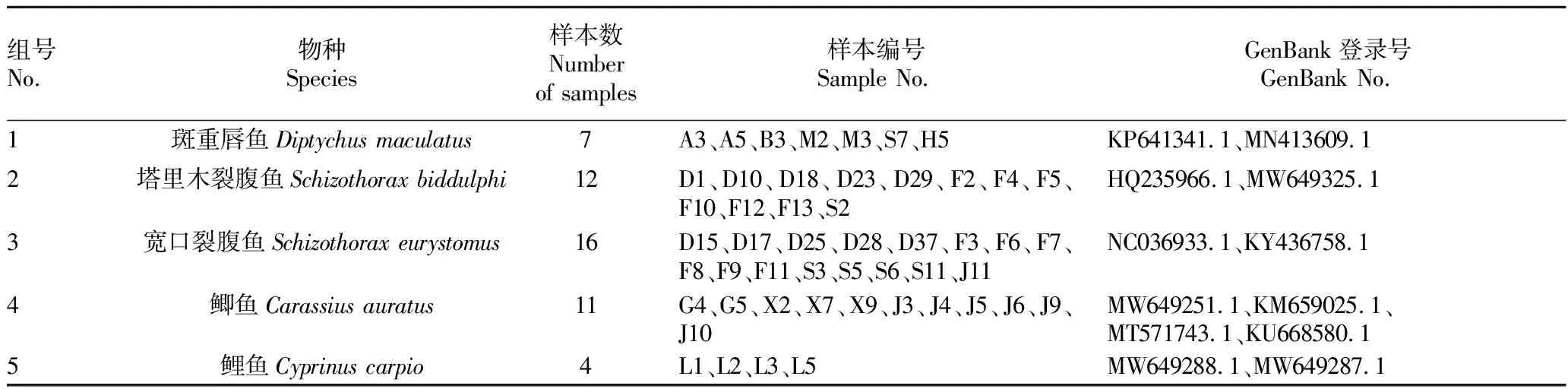
表1 樣本名錄及相關(guān)信息
2 結(jié)果與分析
2.1COI基因序列特征該研究通過(guò)比對(duì)和分析葉爾羌河5種魚(yú)類樣本的COI基因片段序列,再結(jié)合從NCBI下載的12條已知的線粒體基因序列,最終保留675 bp的序列。分析結(jié)果表明(表2),葉爾羌河流5種鯉科魚(yú)類COI基因序列的平均堿基含量分別為T 28.4%、C 28.2%、A 25.6%、G 17.8%,其中鳥(niǎo)嘌呤含量最低,胸腺嘧啶含量最高,且5種魚(yú)類的COI基因片段序列的A+T含量明顯高于G+C,展現(xiàn)出顯著的A/T偏倚性。該研究序列中的變異位點(diǎn)146個(gè),沒(méi)有發(fā)現(xiàn)堿基的插入/缺失,平均轉(zhuǎn)換位點(diǎn)和顛換位點(diǎn)分別為41和13,平均堿基轉(zhuǎn)換/顛換(R值)為3.2。
2.2 遺傳多樣性分析對(duì)葉爾羌河5種魚(yú)類COI基因的Hd、Pi和K值進(jìn)行統(tǒng)計(jì)(表3),共發(fā)現(xiàn)變異位點(diǎn)146個(gè),其中包括了145個(gè)簡(jiǎn)約性信息位點(diǎn),1個(gè)單一多態(tài)位點(diǎn)。整體的Hd、Pi和K值分別為0.838、0.079 39和53.589。在單倍型多樣性指數(shù)(Hd)中,塔里木裂腹魚(yú)最高(Hd=0.509),鯉魚(yú)其次(Hd=0.500),均表現(xiàn)出較高的單倍型多樣性,而鯽魚(yú)(Hd=0.473)、斑重唇魚(yú)(Hd=0.286)和寬口裂腹魚(yú)(Hd=0.118)的單倍型多樣性指數(shù)則較低;在核苷酸多樣性指數(shù)(Pi)中,鯽魚(yú)最高(Pi=0.003 18),寬口裂腹魚(yú)最低(Pi=0.000 17),表明葉爾羌河5種鯉科魚(yú)類群體的核苷酸多樣性均處于較低的水平。
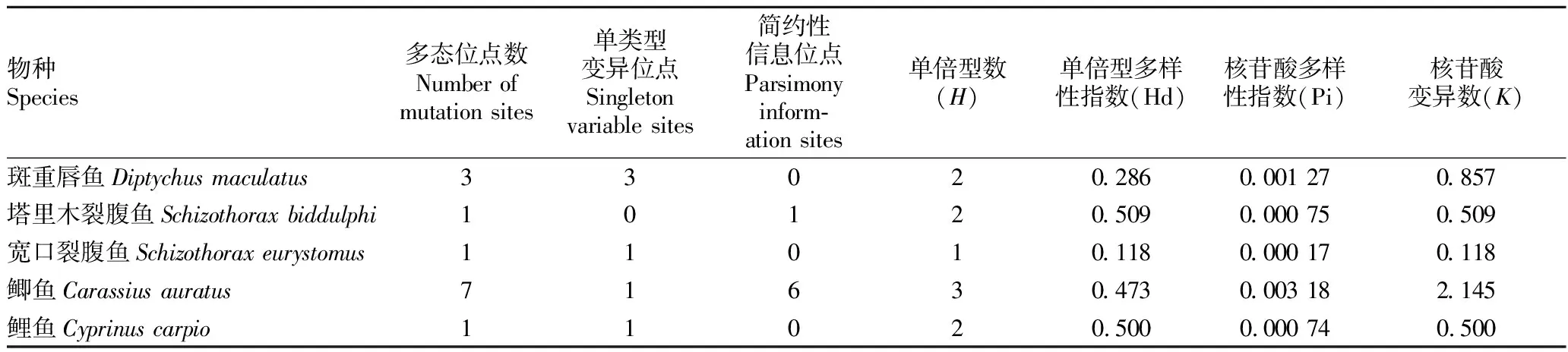
表3 葉爾羌河5種魚(yú)類COI基因序列遺傳多樣性參數(shù)
通過(guò)MEGA 7.0軟件對(duì)葉爾羌河流域5種魚(yú)類的遺傳距離進(jìn)行統(tǒng)計(jì),結(jié)果表明(表4),該研究5種魚(yú)類的種內(nèi)遺傳距離為0~0.003 2,平均值為0.001 6,其中鯽魚(yú)的種內(nèi)遺傳距離最大(0.003 2);種間距離為0.002 4~0.150 5,平均值為0.088 8。其中,斑重唇魚(yú)和鯽魚(yú)的種間遺傳距離最遠(yuǎn)(0.150 5);塔里木裂腹魚(yú)和寬口裂腹魚(yú)的種間遺傳距離最小(0.002 4),推測(cè)與兩者親緣關(guān)系較近有關(guān)。

表4 基于COI基因序列的葉爾羌河流域魚(yú)類群體遺傳距離
2.3 單倍型分析采用DnaSP 5.10軟件檢測(cè)該研究中50個(gè)樣本的COI片段,一共獲得146個(gè)多態(tài)性位點(diǎn),10個(gè)單倍型。其中,Hap1、Hap2在斑重唇魚(yú)中特有;Hap3、Hap5在塔里木裂腹魚(yú)中特有;Hap4在寬口裂腹魚(yú)中特有;Hap6、Hap7、Hap8在鯽魚(yú)中特有;Hap9、Hap10在鯉屬的鯉魚(yú)中特有。
2.4 系統(tǒng)進(jìn)化樹(shù)的構(gòu)建以黃顙魚(yú)(Tachysurusfulvidraco,MH317141.1)作為外群,采用鄰接法(neighbor-joining method,NJ)來(lái)構(gòu)建系統(tǒng)發(fā)育樹(shù)。如圖1所示,該研究中的5種魚(yú)類在系統(tǒng)發(fā)育樹(shù)上的位置清晰,共分為4大支。其中,第一組寬口木裂腹魚(yú)(Schizothoraxeurystomus)和塔里木裂腹魚(yú)(Schizothoraxbiddulphi)聚為一支;第二組鯉魚(yú)(Cyprinuscarpio)聚為一支;第三組斑重唇魚(yú)(Diptychusmaculatus)聚為一支;第四組銀鯽(Carassiusgibelio)、鯽魚(yú)(Carassiusauratus)聚為一支。其中,鯽屬、重唇魚(yú)屬、鯉屬分別單獨(dú)形成一支,裂腹魚(yú)屬的寬口裂腹魚(yú)和塔里木裂腹魚(yú)則共聚為一大支,表明兩者的親緣系較近。
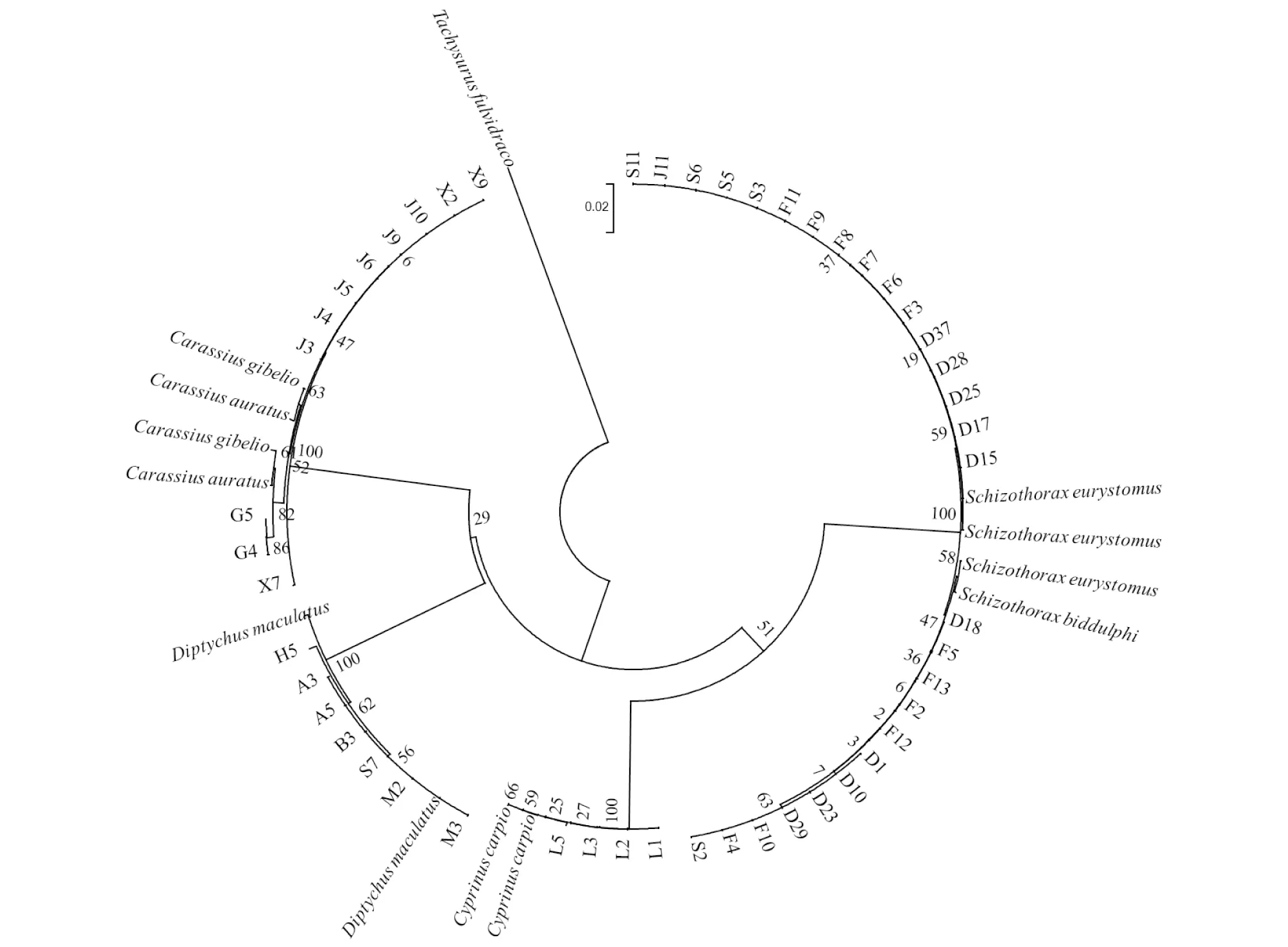
圖1 基于NJ法構(gòu)建的葉爾羌河5種魚(yú)類聚類關(guān)系Fig.1 Clustering relationship of five fish species in Yarkant River based on NJ method
3 結(jié)論與討論
該研究對(duì)葉爾羌河50個(gè)魚(yú)類樣本的線粒體COI基因序列進(jìn)行分析,結(jié)果顯示在堿基組成上,堿基T含量最高(28.4%),堿基G最低(17.8%),且堿基A+T的平均含量(54.0%)明顯高于G+C的平均含量(46.0%),這與張桂寧等[17,20-21]的研究結(jié)果類似,即表現(xiàn)出明顯的A/T堿基偏倚性,與魚(yú)類mtDNACOI基因序列的堿基特征相符。
遺傳多樣性的大小通常通過(guò)Hd和Pi判斷,兩者的數(shù)值越大表示遺傳多樣性越豐富[22]。1998年,Grant等[23]首次提出以0.5作為單倍型多樣性的臨界值,以0.005作為核苷酸多樣性的臨界值。該研究中,塔里木裂腹魚(yú)和鯉魚(yú)均表現(xiàn)出較高的單倍型多樣性(Hd≥0.5)和較低的核苷酸多樣性(Pi<0.005),表明其可能正處于群體的快速擴(kuò)張階段[24],任永麗[20]通過(guò)線粒體COII和ND4基因序列分析3個(gè)塔里木裂腹魚(yú)群體發(fā)現(xiàn)其單倍型多樣性和核苷酸多樣性均較高,推測(cè)可能是由于脊椎動(dòng)物不同線粒體基因序列的進(jìn)化速率不同[25]或與地理隔離有關(guān)。而斑重唇魚(yú)、寬口裂腹魚(yú)和鯽魚(yú)均表現(xiàn)出較低的單倍型多樣性(Hd<0.5)和核苷酸多樣性(Pi<0.005),推測(cè)其可能正經(jīng)歷群體瓶頸效應(yīng)[26]。
該研究中的葉爾羌河5種鯉魚(yú)類的種內(nèi)平均遺傳距離和種間平均遺傳距離分別是0.001 6和0.088 8,符合Hebert等[13]提出的種間遺傳距離需要大于種內(nèi)遺傳距離的10倍以上,這是利用mtDNACOI基因作為有效鑒定物種的前提。但是該研究中的塔里木裂腹魚(yú)與寬口裂腹魚(yú)群體之間的遺傳距離最小(0.002 4),遠(yuǎn)低于Herber等[27]提出的物種間最小遺傳距離(0.02),說(shuō)明該研究中基于COI基因作為分子標(biāo)記未能成功區(qū)分塔里木裂腹魚(yú)和寬口裂腹魚(yú),這與海薩·艾也力汗等[26]基于COI基因分析塔里木和流域裂腹魚(yú)屬的研究結(jié)果一致。此外,通過(guò)分析系統(tǒng)進(jìn)化樹(shù)表明,在屬的階元上,該研究中的鯉科4屬5種魚(yú)類,即重唇魚(yú)屬、裂腹魚(yú)屬、鯽屬和鯉屬分別聚為一支,分類明確。但是裂腹魚(yú)屬的塔里木裂腹魚(yú)和寬口裂腹魚(yú)的分類上并不是很清晰。總的來(lái)說(shuō),以mtDNACOI基因作為分子標(biāo)記,對(duì)葉爾羌河魚(yú)類進(jìn)行種質(zhì)鑒定和多樣性分析是可行的,但是對(duì)于裂腹魚(yú)屬最好搭配其他線粒體DNA序列作為分子標(biāo)記來(lái)分析效果更好,如D-loop區(qū)[28]、Cytb[29]、COII[30]等。
該研究只分析了葉爾羌河流域5種鯉科魚(yú)類的遺傳多樣性,后續(xù)可以進(jìn)一步對(duì)葉爾羌河流域不同區(qū)域開(kāi)展更為詳細(xì)的資源調(diào)查與遺傳多樣性評(píng)估,從而豐富對(duì)葉爾羌河流域魚(yú)類資源狀況的了解,也有助于瀕危魚(yú)種保護(hù)、資源合理開(kāi)發(fā)利用和生態(tài)環(huán)境改善等。

- 安徽農(nóng)業(yè)科學(xué)的其它文章
- 鄉(xiāng)村振興背景下涉農(nóng)學(xué)科專業(yè)平臺(tái)團(tuán)隊(duì)課程一體化建設(shè)的探索與實(shí)踐
——以凱里學(xué)院園藝專業(yè)為例 - 地方涉農(nóng)院校“新農(nóng)科”建設(shè)探索與實(shí)踐
——以遼東學(xué)院農(nóng)學(xué)院為例 - 農(nóng)村生態(tài)資源價(jià)值實(shí)現(xiàn)路徑及其法治保障研究
- 大學(xué)生掛職視角下徐州市銅山區(qū)家庭農(nóng)場(chǎng)創(chuàng)新發(fā)展研究
- 共同富裕背景下閑置宅基地有償退出利用機(jī)制研究
- 新安江流域水生態(tài)環(huán)境形勢(shì)分析及對(duì)策研究