叢枝菌根真菌對酸脅迫下杉木幼苗生長生理的影響
2023-09-28 01:18:04李少青
防護林科技 2023年5期
關鍵詞:生長
李少青
(福建省三明市三元區林業局,福建 三明 365000)
杉木(Cunninghamialanceolata)材質優良、生長快,是我國南方片區重要的造林用材樹種[1]。研究發現,杉木種植會帶來土壤酸化,尤其是多代連作后導致土壤酸化明顯,林木生長量大幅下降,地力衰退嚴重,成為限制杉木人工林高效培育的關鍵因子[2]。土壤pH能影響土壤中養分的存在狀態及轉化利用效率,較低pH是造成土壤酸化的根本原因。調查表明,在酸脅迫下杉木抗逆性低,林分質量較差[3]。
杉木林地中微生物種群豐富[4],地域不同,林齡不同,微生物群落特性及多樣性也不同[5,6]。叢枝菌根真菌作為土壤微生物中普遍存在的群體之一,與80%以上陸地植物建立了良好的共生關系,能促進植物吸收營養,提升對逆境的適應性[7]。陳艷芳、陳鄭洪等發現,杉木連作后,土壤中叢枝菌根真菌多樣性指數、侵染率下降,土壤性質發生改變[8,9]。雷梅等通過接種叢根菌根真菌的方法,有效提升了杉木幼苗對養分的吸收[10]。然而,酸脅迫下叢枝菌根真菌對杉木生長的影響及其生理響應機制方面研究仍鮮見報道。為此,本研究以杉木幼苗為試驗材料,通過菌根接種技術,探討菌根真菌對酸脅迫下杉木生長和相關生理特性的影響,以揭示叢枝菌根真菌在提升杉木耐酸性方面的應用潛力。
1 材料與方法
1.1 試驗材料
在福建三明市杉木優良種源試驗林中,選擇形質優良、無病蟲害成年優樹為采種母樹,于2021年10—11月果實成熟時進行種子采集,待自然曬干、去雜后存放于4 ℃冰箱備用。次年3月將種子取出,經45 ℃溫水浸種12 h后進行沙床播種。出苗后移至裝有等體積比的黃心土、珍珠巖育苗杯中。當苗高15 cm左右時,選擇長勢較好、生長健壯幼苗為試驗材料。
1.2 菌根接種
以北京市農林科學研究院植物營養與資源研究所提供的叢枝菌根化真菌摩西球囊霉(BGC NM01A)和地表球囊霉(BGC GD01C)為混合菌劑,在裝有經高溫高壓滅菌的80%紅壤與15%珍珠巖,以及含5%過磷酸鈣的營養杯(8 cm×12 cm)中進行接種。每個營養杯中放入混合菌劑20 g,約含150孢子的BGC NM01A和500個孢子BGC GD01C。接種菌劑1個月后,采用醋酸墨水染色,取樣側根觀察菌根侵染情況[11]。
1.3 酸脅迫處理
以高、徑生長量無統計學上顯著差異的未接種菌根(N)和菌根侵染明顯(M)的兩種苗木為研究對象,采用水培法,以1/2 Hoagland營養液為培養液,利用2 mol·L-1H2SO4溶液進行pH調整,參照梁海燕等[3]研究結果,設置pH 6.8(P1)、pH 5.5(P2)、pH 4.5(P3)3個酸脅迫處理,共6個處理:NP1、NP2、NP3、MP1、MP2、MP3。每處理設置4重復,每重復50株苗。試驗在控制環境條件下處理30 d(溫度25±1 ℃,相對濕度85%,光照強度180 μmol·m-2·s-1,光照時間16 h,每周更換1次培養液,每日充氣1次,每次30 min)。
1.4 指標測定
幼苗高徑生長增量:以試驗處理期間高徑生長增量表示,用卷尺、游標卡尺分別進行幼苗高度、地徑的測量。
光合參數:各處理每重復選取植株中上部5片功能葉進行活體測定。其中,葉綠素含量采用便攜式葉綠素速測儀測定,光合作用參數采用LI-6400光合作用儀測定。
光合產物含量:可溶性糖和淀粉含量測定均采用蒽酮比色法。
抗氧化指標:超氧化物歧化酶(SOD)、過氧化物酶(POD)、過氧化氫酶(CAT)活性及丙二醛(MDA)含量采用王尚堃[12]方法進行測定。
1.5 數據處理
數據采用SPSS19.0統計軟件進行不同處理間差異顯著性分析(P<0.05),圖表采用Excel制作。
2 結果與分析
2.1 對幼苗生長的影響
由圖1可以看出,供試酸脅迫處理下,隨著pH降低,杉木幼苗的苗高、地徑增量呈明顯的下降趨勢。從叢枝菌根真菌的影響看,除P1處理下,接種(M)與未接種(N)菌根幼苗高徑生長無顯著差異外,在P2和P3處理下,均以接種菌根幼苗的高徑增量較大。這說明,在pH4.5~5.5范圍內,杉木幼苗生長受到了明顯的抑制作用,而通過菌根接種能有效改善苗木的生長情況。
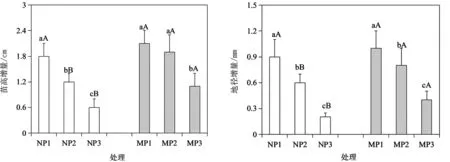
圖1 叢枝菌根真菌對酸脅迫下杉木幼苗生長的影響
2.2 對幼苗光合能力的影響
光合作用是實現植物生長的關鍵生理代謝活動,直接影響著植物生長表現。從表1杉木幼苗光合參數變化來看,在P1處理下,除凈光合速率(Pn)在接種菌根后顯著增加外,葉綠素含量、蒸騰速率(Tr)、水分利用效率(WUE)、氣孔導度(Gs)和胞間二氧化碳濃度(Ci)并未受到叢枝菌根真菌的影響。但在P2和P3處理下,接種菌根后幼苗葉片中葉綠素含量、Pn、WUE、Ci顯著增大,而Tr、Gs顯著減小。試驗結果表明,在酸脅迫下叢枝菌根真菌有效提升了杉木幼苗光合能力。

表1 叢枝菌根真菌對酸脅迫下杉木幼苗光合參數的影響
可溶性糖和淀粉是植物通過光合作用形成的兩大主要光合產物,直接反映植物光合能力。為進一步探究叢枝菌根真菌對酸脅迫下杉木幼苗光合能力的影響,分析了供試處理幼苗葉片中可溶性糖和淀粉含量的變化。由圖2可知,在供試pH4.5~6.8范圍內,隨著pH減小,杉木幼苗葉片中可溶性糖與淀粉含量逐漸降低,但在接種菌根后,各酸處理幼苗葉片中可溶性糖與淀粉含量均顯著增加。這說明,叢根菌根能顯著提升杉木幼苗的光合代謝能力。
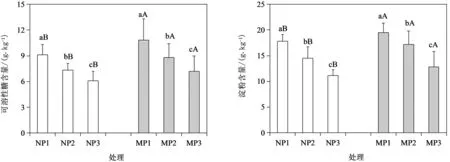
圖2 叢枝菌根真菌對酸脅迫下杉木幼苗光合產物的影響
2.3 對幼苗抗氧化能力的影響
膜系統保護酶活性變化反映著植株遭受逆境脅迫傷害的情況,通常抗逆能力越強抗氧化酶活性越高。表2試驗結果表明,pH越小,葉片中膜脂過氧化物丙二醛(MDA)含量越高,代表供試杉木幼苗受到的酸脅迫傷害程度越重。從酶活性變化看,隨著pH減小,除SOD活性在接種菌根后表現為逐漸增大的趨勢外,POD、CAT活性及未接種菌根的SOD活性均表現為先上升后下降的趨勢。從接種與未接種菌根幼苗間MDA含量及酶活性差異看,除P1處理為無明顯變化外,在P2、P3處理下,接種菌根后幼苗MDA含量下降,SOD、POD、CAT活性上升,這說明叢枝菌根真菌明顯提升了杉木幼苗抗氧化能力,耐酸性增強。
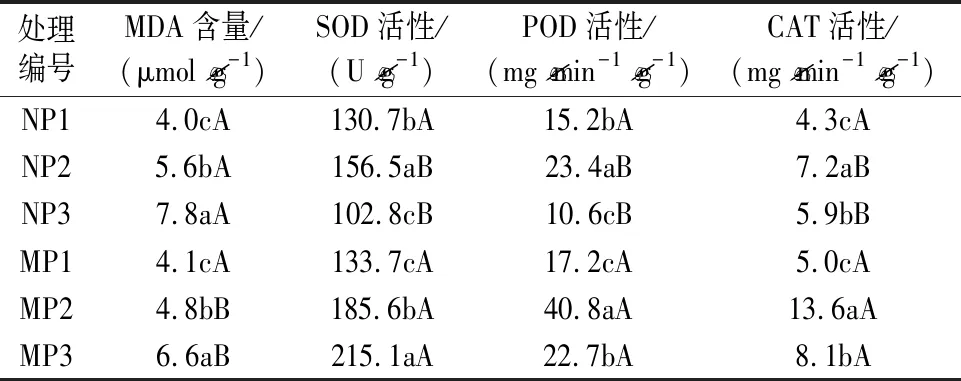
表2 叢枝菌根真菌對酸脅迫下杉木幼苗抗氧化能力的影響
3 結論與討論
植物群落與土壤微生物群落之間存在著緊密聯系,通過共同互生關系調節著生態系統的平衡[13]。研究證明,杉木林中叢枝菌根真菌群落豐富,林齡不同,群落組成也不同。但整體上看,在8~35年生杉木林中,以球囊霉屬的豐度最高(28.77%~54.85%),而其他真菌的豐度相對較低(0.01%~2.72%)[6]。目前,有關叢枝菌根真菌對杉木幼苗的影響鮮見報道,雷梅等[10]通過接種摩西球囊霉方法實現了杉木幼苗對養分吸收效率的提升。本試驗通過模擬自然狀態下叢枝菌根真菌中群落組成,以優勢種群球囊霉屬中地表球囊霉和摩西球囊霉混合菌株為材料進行接種,有效改善了酸脅迫下杉木幼苗的生長效果,這為今后改善杉木耐酸性,實現杉木人工林高效培育提供了可靠的參考。但鑒于自然界中杉木林地叢枝菌根真菌的多樣性,是否仍需進一步優化真菌種類的組成有待試驗驗證。
在pH4.5~6.8范圍內,pH越小,杉木幼苗高徑生長增量越低,這反映了當pH為4.5~5.5時,杉木受到了明顯的酸脅迫傷害,這與以往有關pH在6.0以下時,杉木會遭受脅迫傷害的研究結論是一致的[3,14]。在pH為6.8時,叢枝菌根真菌對杉木幼苗生長無明顯影響,這與雷梅等[10]的研究不同,這可能與苗期杉木生長緩慢,而試驗處理時間較短(僅1個月)有關。但在pH為4.5~5.5時,叢枝菌根真菌表現出了明顯的促生長效果,這暗示了相較正常生長條件,在逆境脅迫下叢枝菌根真菌更能有效提升杉木的抗逆性。從菌根的作用機制看,其本質在于提升共生植株對養分的吸收能力,從而緩解環境脅迫導致的營養失衡問題,保障植株生長所需養分。下一步研究叢枝菌根真菌對杉木營養元素的吸收、運輸、轉化方面的影響,對今后實現杉木精準施肥,解決土壤酸化導致杉木生長量下降的技術瓶頸有望提供科學、可靠的理論指導。
叢枝菌根真菌顯著提高了杉木幼苗的光合能力,光合產物增加。從各光合參數對菌根的響應看,以凈光合速率反應最為靈敏,即在pH為4.5~6.8時,接種菌根后幼苗葉片的凈光合速率均顯著增大。這表明,叢枝菌根真菌對杉木幼苗光合能力的改變主要是通過提升凈光合速率,而非通過減少蒸騰失水等方式實現的。影響植物光合速率的影響因子有很多,除溫度、濕度、光照等外在環境因子外,還與植株中營養元素、酶活性等內在因子有關[15,16]。這暗示了,基于光合速率增強的科學施肥方法或能增強杉木耐酸能力。
以往大量研究表明,膜系統保護酶具有雙重生理指示意義[17,18]。在逆境脅迫下,植物組織中早期酶活性升高,代表其自身對逆境的自我調節能力,活性越強,表明遭受脅迫傷害越嚴重,而隨著脅迫處理時間延長,酶活性降低,表明其膜系統遭到破壞,酶活性越低,脅迫傷害越大。本試驗研究發現,隨著pH增加,未接種菌根幼苗葉片中SOD、POD、CAT活性表現為先上升后下降,這與以往研究是相似的。但在接種菌根后,SOD卻表現為隨pH增加而增大的趨勢。這暗示了,叢枝菌根真菌對酸脅迫下杉木幼苗的抗性提升可能與SOD活性增加有關。SOD活性與微量元素Cu、Zn密切相關,但叢枝菌根真菌是否主要通過增強植株對土壤中Cu、Zn吸收利用率,從而實現對植株抗氧化性、光合能力的增加,以及對幼苗生長的促進,有待進一步證實。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
