重金屬污染物的微生物修復策略
2023-10-08 03:13:52王永剛朵建文冷非凡陳吉祥孫尚琛
安徽農業科學 2023年18期
關鍵詞:研究
王 瑾,王永剛,朵建文,冷非凡,陳吉祥,孫尚琛*
(1.蘭州資源環境職業技術大學,甘肅蘭州 730030;2.蘭州理工大學,甘肅蘭州 730050)
目前,由于快速發展的工業活動、礦山開采、高價態金屬廢棄物傾倒、化肥的過度使用等,重金屬在環境中的遷移加劇、遷移速度大大加快,越來越多的含金屬殘留物被排放到地表進入土壤系統和地下水,并通過食物鏈富集在人體中,進而對人體和生態系統造成嚴重危害[1-2]。當重金屬在土壤中的積累量超過土壤的自凈能力后會改變陸地生態系統中棲息動植物的種類、分布及數量,影響植物根系微生物的群落結構和酶活性,阻礙植物的光合作用、呼吸作用等[3]。當過量的重金屬進入水生系統后,會嚴重阻礙水生動物DNA的合成,導致胚胎畸變、影響性別和身長等[2]。而水生藻類一旦積累了過量的重金屬會導致其代謝和生理功能紊亂,光合作用被抑制,更有甚者導致藻類死亡,改變天然環境中藻類的數量和分布[3]。另外,進入人體的重金屬借助體內某些有機成分可結合成金屬絡合物或螯合物富集在人體內,當積累量超過人體新陳代謝最大限值后,會嚴重損害人體的呼吸系統、消化系統、生殖系統等,甚至造成人體癌變和死亡[3-5]。
基于過量重金屬對生態系統的諸多危害,重金屬已被公認為是對自然動植物群造成嚴重危害的最持久和最復雜的污染因子。目前,對于重金屬污染環境的防治與治理,已發展起來的方法有包括離子交換法、吸附法和膜分離法在內的物理法,沉淀法、氧化還原法等在內的化學法和依賴于微生物的生物法。因微生物具有可培養性強、環境友好、操作簡單等特點,微生物修復策略被認為是在重金屬污染物治理方面最有前景的方式之一。一方面,微生物對某些特定的金屬具有強選擇性,且因其體積小的特點可以為重金屬的高效結合提供大的比表面積[5]。另一方面,微生物吸附是一個獨立和可逆的過程,存活的和死亡的微生物都可以用作金屬吸收的生物吸附劑[6]。此外,微生物還可以通過離子交換、氧化還原、電化學處理、化學沉淀等方式對重金屬進行固定和解毒[1]。所有這些改變重金屬遷移性和毒性的轉變都具有巨大的生物地球化學意義并具有生物修復潛力。另外,在重金屬脅迫下,微生物的聚集和生物膜的形成使微生物能夠維持更長的時間,進一步表明微生物修復機制比傳統方法更經濟和有效。
“微生物-重金屬”是一個較為復雜的生物學命題,該研究從重金屬對微生物的毒性作用、微生物對重金屬脅迫的響應等多方面進行綜述,以期為“微生物-重金屬”的相互作用提供更為全面的理解,為生物修復策略的進一步應用奠定理論基礎。
1 重金屬對微生物的影響
重金屬通常對微生物產生雙向作用,如某些低濃度的重金屬在微生物蛋白質的合成、酶活性構型的維系、氧化還原等過程中發揮作用[7]。但當重金屬的濃度超過微生物生長代謝所需的閾值后會對微生物產生不利影響。
從宏觀角度來講,重金屬會影響微生物的群落結構和生物多樣性,這主要取決于重金屬和微生物的類型。張瀚丹等[8]研究發現,Cr和Ni的濃度與放線菌門(Actinobacteria)和奇古菌門(Thaumarchaeota)的豐度呈負相關,但與擬桿菌門(Bacteroidetes)的豐度呈正相關,Cd、Pb和Hg的濃度與芽單胞菌門(Gemmatimonadetes)呈正相關,但總體上重金屬會導致微生物生物多樣性的下降。何一凡等[9]的研究表明,在受Cr、Pb和Zn污染的土壤中,硝化菌(Nitrobacterium)、擬桿菌(Bacteroidetes)和疣微菌(Verrucomicrobia)的豐度與重金屬濃度呈負相關,但綠彎菌(Chloroflexi)的生長卻受顯著的促進。另外,受低濃度的Cd、Cu、Zn、Pb復合污染的土壤中,細菌、真菌和放線菌的生物量均較污染前顯著增加,表明低濃度的重金屬會刺激微生物群落結構的增加。但隨著重金屬濃度的增加,其對微生物群落結構的影響由刺激變為毒害作用,相應微生物的群落結構相應降低。
從微觀角度來講,高濃度的重金屬離子會破壞微生物細胞壁的完整性、干擾DNA和RNA的合成、誘導部分酶的構型發生變化、阻礙輔因子與生物分子的結合,甚至導致細胞死亡(圖1)[10]。Ranquet等[11]研究發現,隨著大腸桿菌(E.coli)細胞內的Co2+濃度增加,tRNA甲基硫轉移酶、烏頭酸酶和鐵還原酶3種Fe-S酶的活性逐漸降低。另外,Co2+還會干擾微生物細胞的氧化還原及酶的水解過程、抑制細胞分裂。Das等[12]研究發現,在低濃度(<130 μmol/L)的Au4+環境中,米根菌(Rhizopusoryzae) 會通過合成金屬還原硫蛋白對Au4+進行解毒,以維持細胞的正常生長,但當Au4+的濃度超過130 μmol/L后,米根菌的生長受到明顯抑制。另有研究表明,高濃度的Cr6+會通過靜電結合的方式與大腸桿菌和枯草芽孢桿菌(Bacillussubtilis)DNA中的磷酸基團結合,阻礙轉錄和復制過程,導致細菌發生誘變[13]。
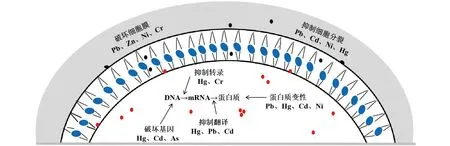
圖1 重金屬對微生物的影響示意Fig.1 Schematic diagram of the effect of heavy metals on microorganisms
2 微生物對重金屬脅迫的響應
盡管重金屬脅迫會顯著降低微生物的多樣性和豐度,及對微生物的生長代謝造成一定毒害,但在長期的重金屬環境中,微生物會形成一定的抗性機制來響應重金屬的脅迫,以維持自身正常的生長代謝。如微生物細胞可利用細胞壁表面的負電荷官能團對重金屬離子進行螯合,以避免過量的重金屬進入細胞質[14]。其次,在重金屬的刺激下,微生物的谷胱還原蛋白和硫氧還原蛋白基因被激活,進而可實現對重金屬離子的氧化還原。另外,微生物細胞還可利用一些存在于細胞膜上的轉運蛋白實現對重金屬離子的吸附、轉運及外排等。總體來說,微生物對重金屬脅迫的響應機制可分為細胞外屏障、細胞外螯合、細胞內絡合、細胞內外排、金屬離子還原5類。
2.1 細胞外屏障通常,重金屬離子必須先穿透細胞壁才可進入細胞膜,但因細胞壁獨特的生理結構,尤其是革蘭氏陽性菌細胞壁肽聚糖層厚而致密,且鑲嵌有磷壁酸的特點,細胞壁在抵御外界刺激過程中發揮分子篩的功能,其可阻止分子量大于600 Da的化合物進入細胞質[15]。另外,一些革蘭氏陽性菌如括紅球菌(Rhodococcus)、分枝桿菌(Mycobacterium)、諾卡氏菌(Nocardia)、棒狀桿菌(Corynebacterium)、戈登氏菌(Gordonia)等由于其細胞壁中富含霉菌酸,使得微生物細胞表面疏水性和滲透性極差,進而阻止了重金屬離子的滲透[16]。而革蘭氏陰性菌由于存在于其細胞外膜的磷脂和脂多糖不斷地旋轉和側移,且脂多糖的酰基鏈比磷脂鏈更硬,排列更緊密,致使其對疏水性化合物的滲透性更低,從而使得革蘭氏陰性菌在細胞外對重金屬有較強抗性[17]。
細胞質膜的通透性在阻止重金屬離子進入細胞質的過程中亦發揮重要作用。當微生物細胞受重金屬脅迫后,細胞膜的脂肪酸組成、鏈長和磷脂組成會發生相應的變化以調整細胞膜的通透性,進而保證了細胞膜的活力[16-17]。在研究不同濃度的Cu2+、Pb2+、Zn2+和Cr6+對嗜根寡養單胞菌(Stenotrophomonasrhizophila)活性和細胞膜通透性的影響時發現,0~120 mg/L的金屬離子對細菌的生長均無影響,在Cu2+、Pb2+和Zn2+環境中的細菌的細胞膜通透性隨著金屬離子濃度的增加而增加,但當Cr6+濃度超過80 mg/L后,細胞膜通透性逐漸降低[15]。另外,細胞膜上大量脂肪酸鏈的存在也會阻礙重金屬離子進入細胞質[17]。細胞莢膜作為一些細菌表面的特殊結構,在微生物細胞響應重金屬的刺激中同樣發揮重要作用,其可作為一種擴散屏障抵抗重金屬進入細胞質。
2.2 細胞外螯合利用微生物在生長代謝過程中分泌的螯合蛋白將重金屬在細胞外進行螯合是降低重金屬毒性,維持細胞正常生長的方式之一。目前,已證明可胞外螯合重金屬的蛋白有鐵載體、草酸鹽、磷酸鹽和硫化物等,而細菌只產生細胞外鐵載體,是一種含有異羥肟酸鹽、兒茶酚酸鹽、羧酸鹽的多肽[18]。研究發現,銅綠假單胞菌(P.aeruginosa)分泌2種鐵載體,其中綠膿桿菌螯鐵蛋白pyochelin可以螯合Cd2+、Co2+、Cu2+、Pb2+、Ga3+、Hg2+、Zn2+、Mn2+、Ni2+、Sn2+、Tb3+等金屬離子[19],另一種鐵載體熒光嗜鐵素pyoverdines可螯合U6+、Np5+等金屬離子[20]。另外,鐵載體還可以促進部分植物對重金屬離子的吸收,Dimkpa等[21]的研究證明,唐德鏈霉菌(Streptomycestendae)F4分泌的鐵載體去鐵胺desferrioxamineB、coelichelin和desferrioxamine E可使向日葵對Cd2+的吸收能力顯著增加,來源于銅綠假單胞菌(P.aeruginosa)的熒光嗜鐵素pyoverdine和綠膿桿菌螯鐵蛋白pyochelin能顯著促進玉米對Cr6+和Pb2+的吸收[22]。
另外,通過一些硫酸鹽還原菌在胞外與重金屬形成硫化物沉淀是實現細胞外螯合的另一種途徑。如有研究證明,硫還原泥土桿菌(Geobactermetallireducens)可將Mn4+還原為Mn2+,將U6+還原為U4+,進而使兩種金屬離子的毒性減弱[23]。金屬還原地桿菌(Geobactersulfreducens)可將強度性的Cr6+還原為弱毒性的Cr3+[24]。
2.3 細胞內絡合微生物細胞經外界刺激后,可在胞內分泌大量的金屬硫蛋白(MTs)、金屬伴侶蛋白和谷胱甘肽(GSH)等活性大分子物質,這類物質因其結構特點對重金屬有極強的結合能力(圖2)[4]。其中MTs是一類富含Cys的金屬結合蛋白,其高級結構由含4個金屬離子結合位點的羧基端和含3個金屬離子結合位點的氨基端組成,能高效結合不同類型的金屬離子[24]。研究發現,Cd和Zn可誘導聚球藻(Synechococcus)smtA和smtB基因編碼的MT的合成,其中smtA基因編碼的MT中的半胱氨酸殘基幾乎可以結合所有類型的金屬離子,而smtB基因編碼的MT含有4個Zn2+結合位點,負責對Zn2+的特異性結合[25]。Su等[26]通過將人MT基因hMT-1A克隆至大腸桿菌JM109中后發現,與原始菌株相比,表達hMT-1A基因的大腸桿菌突變株對As3+的吸附效率提高了3倍。Zhao等[27]將豌豆的金屬硫蛋白表達基因表達至大腸桿菌JM109中后發現,重組大腸桿菌對Hg2+的吸附效率提高了6.4倍。

圖2 微生物對重金屬的胞內絡合機理示意Fig.2 Schematic diagram of intracellular complexation mechanism of heavy metals by microorganisms
谷胱甘肽(GSH)是由Glu、Cys和Gly組成的含有γ-酰胺鍵和巰基的三肽,在微生物抵抗外界刺激中同樣發揮重要作用。其中Cly上的羧基和Cys上的巰基是谷胱甘肽能穩定的結合金屬離子的主要原因[13]。任吉民等[28]研究發現,在水溶液中,GSH通過其Gly兩端的羧基與Dy3+、Ho3+、Yb3+等稀土離子可形成穩定的遙爪配位結構,進而減弱金屬離子的毒性,該結論亦被Podanyi等[29]證實。另外,劉建華等[30]的研究發現,Cr2+和Cr3+可與GSH中的S原子結合形成9種穩定的配位化合物,而Wang等[31]利用EDS研究Cd2+脅迫對路德維希腸桿菌(E.ludwigii)LY6的影響時發現,菌株表面形成的顆粒物為CdS沉淀,進一步表明GSH在微生物吸附重金屬中具有重要作用。
2.4 細胞內外排許多微生物可通過主動運輸或外排系統將存在于細胞質中的金屬離子排出至細胞外,以避免重金屬對微生物造成毒害。目前,對于重金屬的外排公認的是CDF、RND和ATPase三大外排系統,其家族成員可特異性或非特異性地將重金屬離子通過跨膜轉運運輸至細胞外(圖3)[4]。其中CDF家族成員廣泛存在于細菌、古生菌和真核生物,蛋白結構主要由300~550個氨基酸組成,包括6個跨膜結構域、1個C-端結構域和1個富含組氨酸的結構域[30]。研究發現,嗜根寡養單胞菌JC1基因組中含有2個CDF家族的成員,其中czcD/zitB被預測可能參與Co2+、Cd2+、Zn2+的轉運和外排[15],而其在Ralstoniametallidurans中的同源物已被證明參與Ni2+和Cu2+的轉運[32]。嗜根寡養單胞菌中的另一個CDF家族成員是一個膜蛋白,該蛋白在Achromobacterxylosoxidans中被證明參與Pb2+的轉運[33]。另外,Von Rozycki等[34]的研究表明,CupriavidusmetalliduransCH34基因組中存在3個CDF家族的成員,主要參與Fe2+、Co2+、Ni2+、Zn2+和 Cd2+的轉運。RND家族的蛋白質是位于細胞質膜上的三聚體蛋白,它們與1個三聚體外膜因子OMF和周質蛋白MFP形成蛋白復合物。依據可轉運蛋白的類型和作用機理,Nies將負責重金屬轉運的RND家族成員分為Zn2+/Co2+/Cd2+轉運蛋白 (HME1)、Ni2+/Co2+轉運蛋白(HME2)、Zn2+(HME3a)轉運蛋白、Co2+(HME3b)轉運蛋白和 Cu+/Ag+(HME4)轉運蛋白5類,但研究發現嗜根寡養單胞菌基因組中的RND成員中不含特異性負責Ni2+/Co2+轉運的蛋白質,預測到1個參與Mg2+/Co2+轉運的蛋白質[15],這可能與微生物的進化與遺傳背景有關。P-ATPase是一種重要的轉運蛋白家族,可攜帶多種重金屬的底物,從結構上考慮P-ATPase具有多個嚴格保守的Cys序列的殘基用于識別和螯合親水性重金屬離子。研究發現,海氏腸球菌(Enterococcushirae)使用稱為CopA和CopB的CPx型 ATP 酶轉運Cu+和Ag+,惡臭假單胞菌(Pseudomonasputida)S4通過ATP水解從細胞質中輸出Cu2+,隨后在細胞周質中螯合Cu2+。
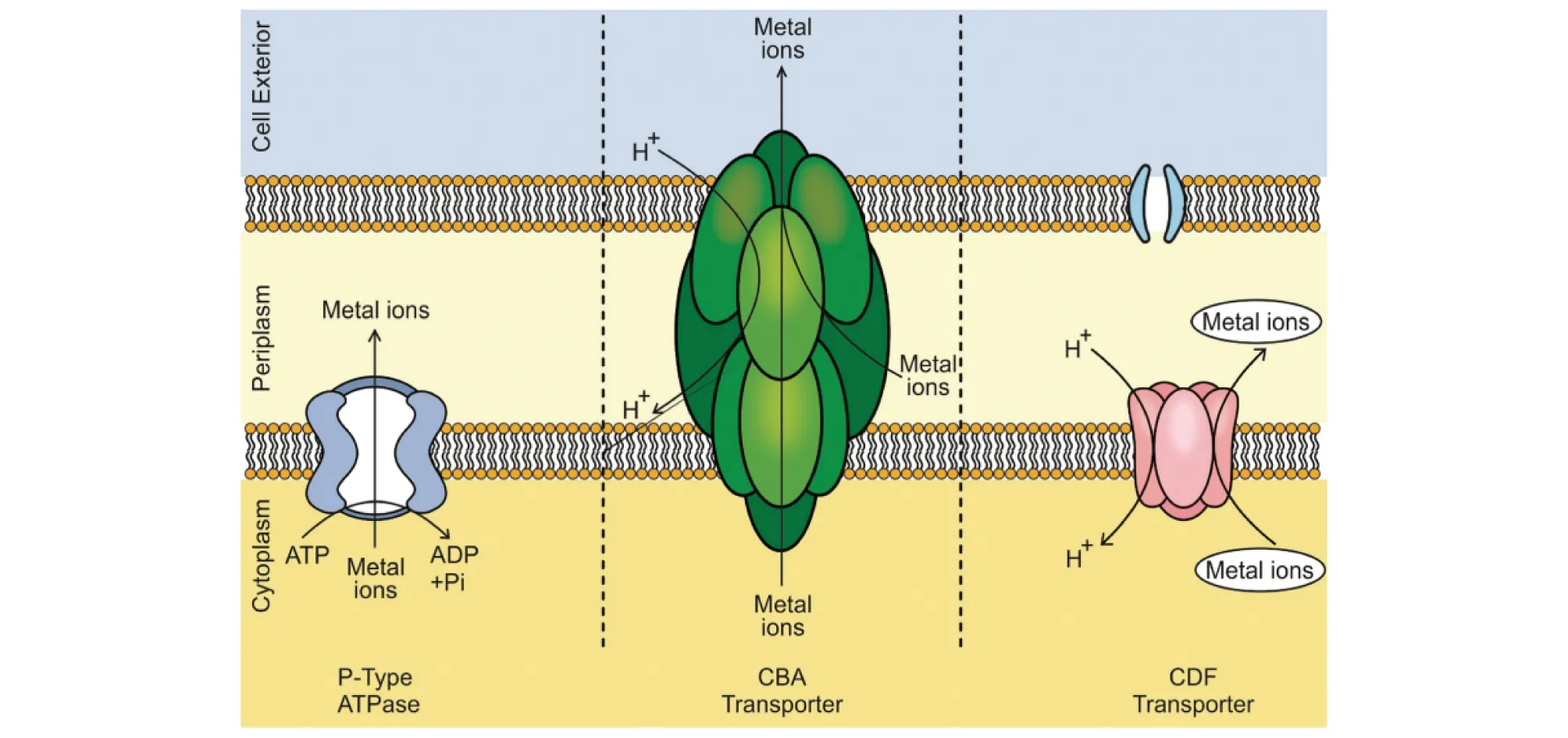
圖3 三大外排系統的作用示意Fig.3 Function diagram of the three efflux systems
除上述3種外排系統外,CHR蛋白、NreB和CnrT系統在解毒Cr6+、Ni2+和Co2+方面也發揮重要作用。CHR介導鉻酸鹽流出并由化學滲透梯度驅動,而CnrT和NreB樣蛋白則由質子動力和CDF蛋白驅動[35]。
2.5 金屬離子的生物還原通過生物還原的方法,降低高價態重金屬離子的化合價,使其呈弱毒性或無毒性,并降低其在自然界中的遷移速率是微生物解毒重金屬的另一途徑。研究表明大多數細菌如地桿菌(Geobacter)、脫硫單胞菌(Desulfuromonas)、微弧菌(Vibrio)等均可利用金屬元素作為終端電子受體。如Thamdrup[35]的研究發現,Geobactermetallireducens可利用Fe(III)、Cr(VI) 和 Mn(IV)作為終端電子受體來保存能量和維持正常生長。Prabhakaran 等[24]的研究表明,砷酸還原菌能將土壤中的As5+還原為As3+,進而使植物根系中的As5+含量減少11.5%左右。此外,細胞色素、氫、乙醇、乳酸鹽等亦可作為電子供體參與微生物對重金屬的還原。研究表明,細胞色素a參與硫酸鹽還原菌對Cr6+的還原,細胞色素c和細胞色素b分別參與陰溝腸桿菌和大腸桿菌對Fe3+和Mn4+的還原。白桿菌屬利用氫、乙醇、乳酸鹽、甲酸鹽等作為電子供體對Fe3+和Mn4+進行還原。
3 展望
微生物具有可培養性強、環境友好、操作簡單等優勢,微生物修復技術目前被認為是重金屬污染場地的修復中最有潛力的方式。現階段,已較清晰地掌握了微生物解毒重金屬的生理機制,這將為微生物修復技術的廣泛應用奠定理論基礎。但目前報道的大多數微生物對重金屬高效的解毒能力均基于實驗室理想的條件下,而一旦應用于工程化,受外界諸多不確定因素的影響,其解毒效率將大幅降低。為此,后期需著力挖掘可廣泛應用于工程化的微生物資源。另外,工程菌的構建是實現微生物修復技術高效、可持續應用的有效途徑,為此在后期的工作中需通過基因組、轉錄組和代謝組結合的方式,挖掘相關微生物基因組中參與重金屬解毒的基因資源,為工程菌的構建提供基因資源。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
