白化和黃化茶樹品種綠茶游離氨基酸、兒茶素類及咖啡堿差異分析
2023-10-17 06:13:44徐玉婕戴曉晶吳紀忠劉亞芹周漢琛楊霽虹王輝黃建琴丁勇雷攀登
熱帶亞熱帶植物學報 2023年5期
徐玉婕, 戴曉晶, 吳紀忠, 劉亞芹, 周漢琛, 楊霽虹, 王輝,黃建琴, 丁勇, 雷攀登*
白化和黃化茶樹品種綠茶游離氨基酸、兒茶素類及咖啡堿差異分析
徐玉婕1, 戴曉晶2, 吳紀忠2, 劉亞芹1, 周漢琛1, 楊霽虹1, 王輝1,黃建琴1, 丁勇1, 雷攀登1*
(1. 安徽省農業科學院茶葉研究所,安徽 黃山 245000;2. 廣德市農業農村局,安徽 宣城 242200)
為探究以白化和黃化茶樹品種鮮葉為原料制成的綠茶滋味品質和代謝物差異,對廣德市6個白化品種綠茶(奶白茶)和14個黃化品種綠茶(黃金芽茶)進行感官審評和代謝物分析。結果表明,奶白茶滋味鮮爽而收斂性略弱;黃金芽茶滋味收斂性強而鮮度低于奶白茶。游離氨基酸總量以及呈現鮮味、甜味的游離氨基酸在奶白茶中的含量顯著高于黃金芽茶,而貢獻收斂性的兒茶素類化合物和沒食子酸含量以及呈現苦味的咖啡堿含量在奶白茶中顯著低于黃金芽茶。偏最小二乘法判別分析(PLS-DA)表明導致兩種綠茶滋味差異的標志性化合物有7種,分別是茶氨酸、精氨酸、谷氨酸、天冬氨酸、表沒食子兒茶素沒食子酸酯(EGCG)、表兒茶素沒食子酸酯和咖啡堿。味覺活性值(Dot)最高的EGCG在黃金芽茶中的呈味貢獻顯著高于奶白茶。因此,游離氨基酸、兒茶素類化合物、沒食子酸和咖啡堿含量差異導致白化和黃化茶樹品種綠茶滋味不同。
茶樹;白化茶樹;黃化茶樹;綠茶;滋味成分
白化和黃化茶樹是在特定環境下因葉綠體合成受阻,新梢色澤呈現不同程度白化或黃化的葉色突變體,具有一定的遺傳穩定性,是我國重要的茶樹特異種質資源[1–2]。根據其對環境的響應不同, 可分為低溫敏感型和光照敏感型,15 ℃下‘小雪芽’嫩梢為白色,溫度升高,葉片復綠;‘黃金芽’在自然光下發出黃色嫩梢,遮蔭后葉片復綠[3–5]。白化和黃化茶樹品種在發育過程中的特征代謝物積累和代謝調控顯著區別于正常葉色品種,其游離氨基酸及茶氨酸含量顯著富集,而兒茶素和咖啡堿等含量顯著降低[6–8]。因此,以白化和黃化茶樹品種鮮葉為原料制作的綠茶,口感更加鮮爽且苦澀味降低。
茶湯滋味是評價茶葉品質的一個重要因素。游離氨基酸、多酚類、嘌呤堿、有機酸、多糖等滋味物質的含量和比例決定茶湯滋味。不同滋味物質由于其各自的味感閾值及在茶湯中的不同含量而對茶湯滋味構成有不同程度的貢獻[9]。Zhang等[10]的研究表明,黃酮醇--糖苷、單寧和酯型兒茶素是貢獻茶湯澀味的主要化合物,而咖啡堿和非酯型兒茶素會增強茶湯的苦味。此外,l-茶氨酸、琥珀酸、沒食子酸對茶湯鮮味的貢獻較大。茶湯中的滋味物質間存在味覺上的相互作用,表沒食子兒茶素沒食子酸酯能協同提高咖啡堿的苦味和澀味,抑制蔗糖的甜味[11]。l-茶氨酸、琥珀酸和沒食子酸可以增強茶湯中l-谷氨酸鈉的鮮味[12]。豐富的滋味物質和不同化合物的味覺相互作用造成茶湯口感各異。
白化/黃化品種葉片中化合物含量的差異與茶樹生長發育過程中的代謝調控密切相關。Lu等[13]對白化葉片和對照葉片進行RNA測序和差異表達基因分析,結果表明白化葉片中葉綠體的缺失可能破壞了碳氮代謝的平衡,導致游離氨基酸大量積累,而多酚含量較低。低溫敏感型品種‘安吉白茶’在白化期間可溶性蛋白含量降低,游離氨基酸含量升高[14]。Dong等[15]的研究表明,白化誘導之后的黃色葉片比正常綠葉中香氣代謝物含量顯著降低。白化茶樹品種‘小雪芽’白化期咖啡堿、兒茶素類化合物的含量明顯低于綠葉期[16]。這些代謝物積累的不同造成所制成品茶的品質特征具有差異。
本研究比較了白化品種鮮葉所制的奶白茶和黃化品種鮮葉所制的黃金芽茶滋味差異,并分析了二者的主要滋味化合物含量,并進一步探究造成白化和黃化茶樹品種綠茶滋味不同的差異化合物,為研究白化和黃化茶樹品種綠茶品質形成的機理提供理論基礎。
1 材料和方法
1.1 樣品與試劑
供試綠茶樣品均于2021年3月—4月產自安徽省廣德市不同地區,原料為一芽一、二葉,按鮮葉來源分為奶白茶(白化品種, NB1~NB6)和黃金芽茶(黃化品種, HJY1~HJY14)。均按照理條殺青-攤涼-理條做形-干燥的加工工藝制成。
游離氨基酸標準品(德國SYKAM公司);茶氨酸標準品(北京百靈威科技有限公司);兒茶素單體和咖啡堿標準品(美國SIGMA公司);色譜級85% 磷酸(ROE公司);色譜級甲醇和乙腈(Thermo Fisher公司);屈臣氏蒸餾水(規格500 mL/瓶);高效液相色譜儀(Chromaster,日本日立公司);氨基酸分析儀(S-433D,德國SYKAM公司)。
1.2 感官審評方法
審評小組由9名專業審評人員組成,對審評人員進行味覺訓練,并對20個綠茶樣品進行預審評。參照張翔等[17]的方法,將綠茶滋味劃分為鮮度、濃度、強度和協調度4項因子,評分標準見表1。參照國家茶葉感官審評標準GB/T 23776—2018,準確稱取3.0 g綠茶樣品,置于150 mL審評杯中, 用100 ℃沸水沖泡4 min,濾出茶湯后由9名專業審評人員進行感官審評。

表1 綠茶滋味因子感官評分標準
1.3 茶湯的制備
參照周漢琛等[18]的方法制備待測茶湯,準確稱取0.250 0 g磨碎茶樣裝入茶包,熱封口后置于具塞25 mL三角瓶,加8 mL沸水,90 ℃水浴7 min,重復提取3次后定容至25 mL。提取液分別稀釋10和4倍,并用0.45m微孔濾膜過濾,用于兒茶素組分與咖啡堿、游離氨基酸含量分析。每個樣品均3次重復。
1.4 游離氨基酸含量的測定
參照周漢琛等[19]的方法測定,色譜柱為LCAK07/Li,柱溫為38 ℃~74 ℃,梯度升溫;流動相A為pH 2.9檸檬酸鋰溶液,流動相B為pH 4.2檸檬酸鋰溶液,流動相C為pH 8.0檸檬酸鋰溶液;洗脫泵流速為0.45 mL/min,衍生泵流速為0.25 mL/min, 進樣量50L。以標準品響應面積計算游離氨基酸含量。
1.5 兒茶素組分和咖啡堿含量的測定
采用高效液相色譜法測定,色譜柱為InfintyLab Proshell 120 SB-C18 (2.7m, 150 mm×4.6 mm, Agi- lent公司)。流動相A為0.04%磷酸溶液,流動相B為乙腈,流動相C為甲醇,流速1.0 mL/min,紫外檢測器波長280 nm;柱溫40 ℃。梯度洗脫程序為:初始流動相90% A+7.5% B+2.5% C,保持1 min; 85% A+11.2% B+3.8% C,保持22 min; 15% A+85% B, 保持13 min; 90% A+7.5% B+2.5% C,保持25 min。以標準品繪制標準曲線,計算兒茶素及咖啡堿含量。
1.6 數據的統計分析
利用SPSS 20.0軟件進行ANOVA計算,采用Duncan (D)方差分析法對樣品中代謝物含量差異進行分析(<0.05);利用SMICA.P軟件對主要滋味化合物進行偏最小二乘法判別(partial least squares discriminant analysis, PLS-DA)分析;采用TB tools對樣品中化合物組分含量進行熱圖分析。
2 結果和分析
2.1 感官審評結果分析
由專業審評人員對奶白茶和黃金芽茶的干茶、葉底、湯色進行審評(圖1)。兩種綠茶外形相似,呈朵形,嫩綠微黃,HJY4、HJY5的干茶色澤偏綠。奶白茶的湯色嫩綠明亮,黃金芽茶湯色嫩綠或淺黃綠色;2種綠茶芽較肥壯,奶白茶葉底呈現玉白色,黃金芽茶葉底呈現玉黃色。NB6較其他奶白茶和黃金芽茶葉底和湯色偏黃。
對奶白茶和黃金芽茶茶湯的4項滋味因子進行審評,二者在滋味上呈現不同的特征(表2)。奶白茶的鮮味特征更加明顯,鮮度評分為(8.61±0.38)~ (9.67±0.26),黃金芽茶為(5.69±1.24)~(8.24±1.15)。黃金芽茶的強度特征更加明顯,即收斂性更強,強度評分為(6.14±0.94)~(7.99±0.5),奶白茶為(3.82± 0.64)~(4.61±0.66)。奶白茶和黃金芽茶的濃度評分分別為(5.96±1.06)~(6.8±1.1)和(5.76±0.83)~(7.63±0.62),協調度評分分別為(8.18±0.56)~(9.19±0.48)和(6.92± 0.56)~(8.84±0.48)。
2.2 關鍵滋味化合物含量分析
對奶白茶和黃金芽茶的關鍵滋味成分游離氨基酸、兒茶素類化合物以及咖啡堿的含量進行測定,結果表明2種綠茶的主要滋味成分的含量存在差異。
游離氨基酸是茶葉呈現鮮味的重要物質,其中茶氨酸是茶葉中特有的游離氨基酸,也是茶葉中含量最高的游離氨基酸,占游離氨基酸總量的50%~ 70%[20–21]。從圖2可見,奶白茶和黃金芽茶中共測定了16種游離氨基酸,奶白茶的游離氨基酸總量和茶氨酸含量顯著高于黃金芽茶(<0.05)。
兒茶素類化合物是茶葉苦澀味和收斂性的重要來源,從奶白茶和黃金芽中共檢測了6種兒茶素,分別為表沒食子兒茶素(epigallocatechin, EGC)、兒茶素(catechin, C)、表兒茶素(epicatechin, EC)、表沒食子兒茶素沒食子酸酯(epigallocatechin gallate, EGCG)、沒食子兒茶素沒食子酸酯(gallocatechin gallate, GCG)和表兒茶素沒食子酸酯(epicatechin gallate, ECG),奶白茶的兒茶素總量、酯型兒茶素含量顯著低于黃金芽茶(<0.05),非酯型兒茶素含量的差異不顯著(> 0.05)(圖3)。

表2 滋味評分
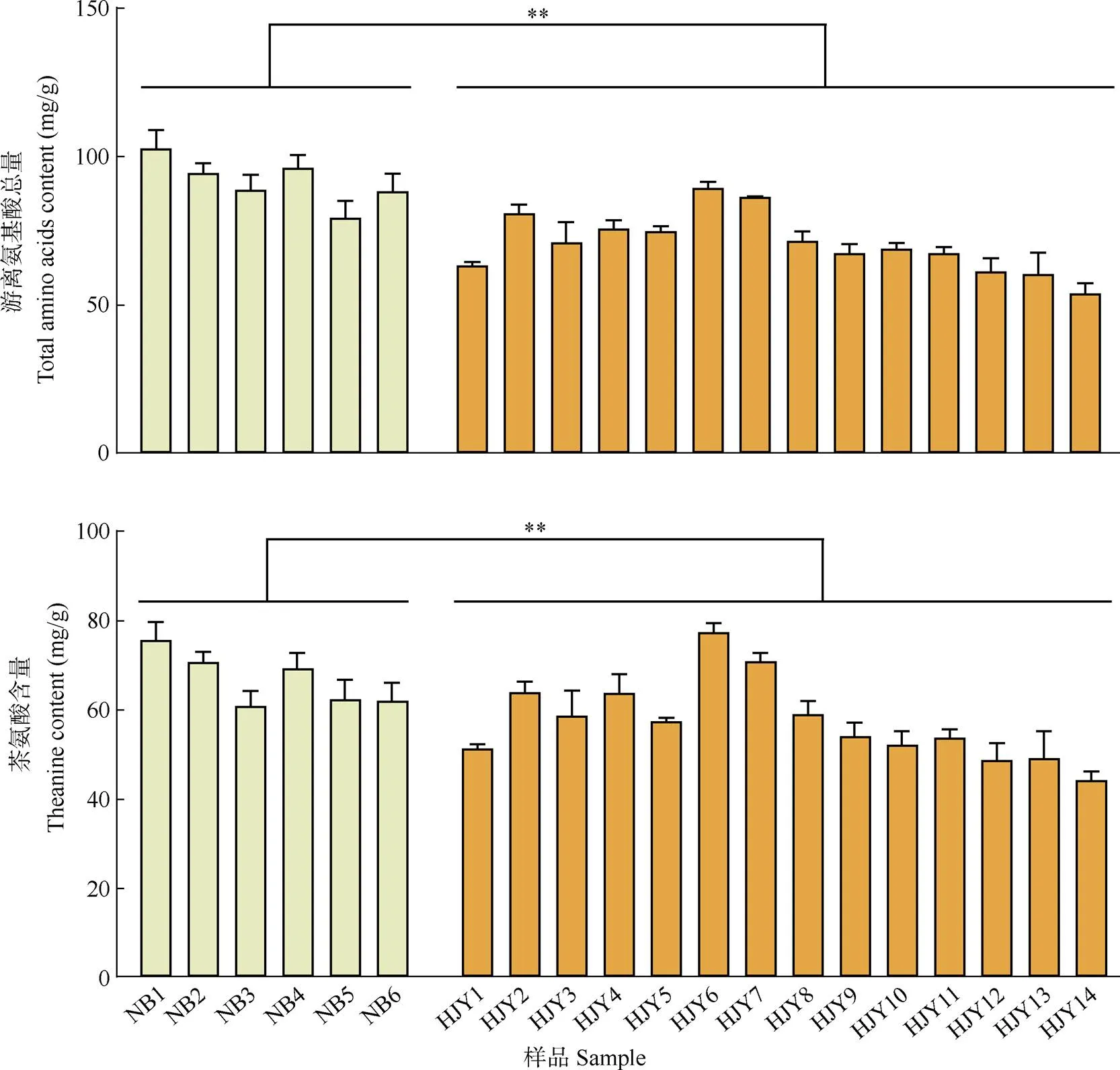
圖2 綠茶中游離氨基酸總量和茶氨酸含量。**: P<0.01。
對6種奶白茶和14種黃金芽茶中的滋味物質含量均值進行分析(圖4)。奶白茶中游離氨基酸總量、茶氨酸總量顯著高于黃金芽茶(<0.05),黃金芽茶的兒茶素總量以及酯型兒茶素總量顯著高于奶白茶(< 0.05), 非酯型兒茶素的含量沒有顯著差異(>0.05), 其中酯型兒茶素的含量差異最大,奶白茶為27.78 mg/g, 而黃金芽茶達到57.10 mg/g,是奶白茶的2倍多。
對16種游離氨基酸、6種兒茶素類化合物、沒食子酸和咖啡堿含量進行熱圖分析(圖5),結果表明奶白茶和黃金芽茶的游離氨基酸組分和兒茶素類化合物組分存在顯著差異。鮮味氨基酸(茶氨酸、天冬氨酸、谷氨酸)和甜味氨基酸(蘇氨酸、甘氨酸、丙氨酸)在奶白茶中的含量明顯高于黃金芽茶。而苦味氨基酸在黃金芽茶中含量更高,比如纈氨酸、異亮氨酸、亮氨酸3種苦味氨基酸在HJY5、HJY10中含量明顯高于6種奶白茶。酯型兒茶素EGCG和ECG在奶白茶中的含量明顯低于黃金芽茶,酯型兒茶素GCG和兒茶素單體C在奶白茶中的含量明顯高于黃金芽。沒食子酸和咖啡堿在奶白茶中的含量明顯低于黃金芽茶,除了HJY12、HJY13和HJY14的咖啡堿含量略低于奶白茶。
2.3 關鍵滋味化合物的差異分析
通過偏最小二乘法判別法(PLS-DA)分析兩種綠茶滋味化合物差異,以VIP>1且<0.05篩選差異化合物, 結果表明,兩種綠茶在游離氨基酸和兒茶素類化合物(包括沒食子酸和咖啡堿)存在顯著差異(圖6: A, B)。通過PLS-DA共篩選出7個差異化合物,包括4個游離氨基酸、2個兒茶素類化合物和咖啡堿。差異游離氨基酸分別為茶氨酸、精氨酸、谷氨酸、天冬氨酸,其中茶氨酸在兩種綠茶中的差異最為顯著,VIP值達到2.5 (圖6: C);產生差異的兒茶素類化合物分別為酯型兒茶素EGCG和ECG,其中,EGCG的差異最為顯著,VIP值為2 (圖6: D)。
2.4 關鍵滋味化合物與呈味貢獻分析
味覺活性值(does-over-threshold, Dot)是指某一滋味成分濃度與其滋味閾值的比值,一般認為Dot>1的滋味物質對茶湯呈味有顯著貢獻。采用Dot值對化合物在茶湯中的滋味貢獻進行量化,滋味閾值來源于Scharbert等[22]和Hufnagel等[23]。表3為Dot值大于1的化合物,分別為EGCG、ECG、EGC、EC、GA和CAF,這表明茶湯中EGCG、ECG、EGC和CAF的濃度均高于滋味閾值。Dot值最高的化合物是EGCG和CAF,分別為8.11~39.35和11.55~19.66,表明EGCG、CAF為茶湯中苦澀味的主要貢獻物質。此外,ECG和EGC對茶湯滋味也有較大貢獻。EGCG和ECG在黃金芽中的Dot值明顯高于奶白茶,二者在奶白茶中的Dot值分別為8.11~19.74和1.38~3.09;在黃金芽茶中分別為21.01~39.35和4.50~7.59,表明酯型兒茶素EGCG和ECG對黃金芽茶湯收斂性貢獻更大。此外,GA呈現酸味和澀味, 在黃金芽茶中的Dot值均大于1,而在奶白茶中均小于1。
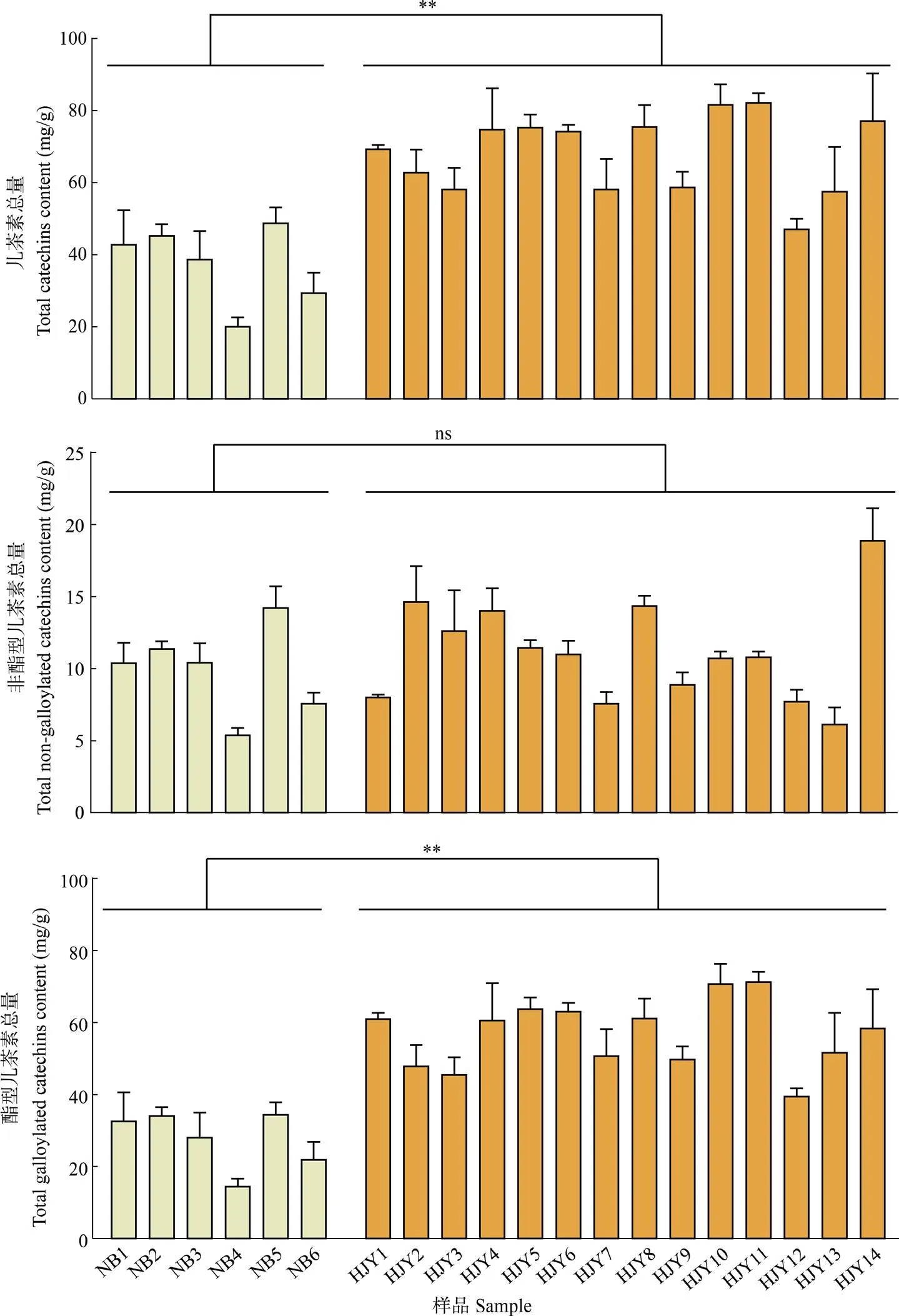
圖3 綠茶中兒茶素類化合物、非酯型兒茶素和酯型兒茶素含量。**: P<0.01; ns: P>0.05。
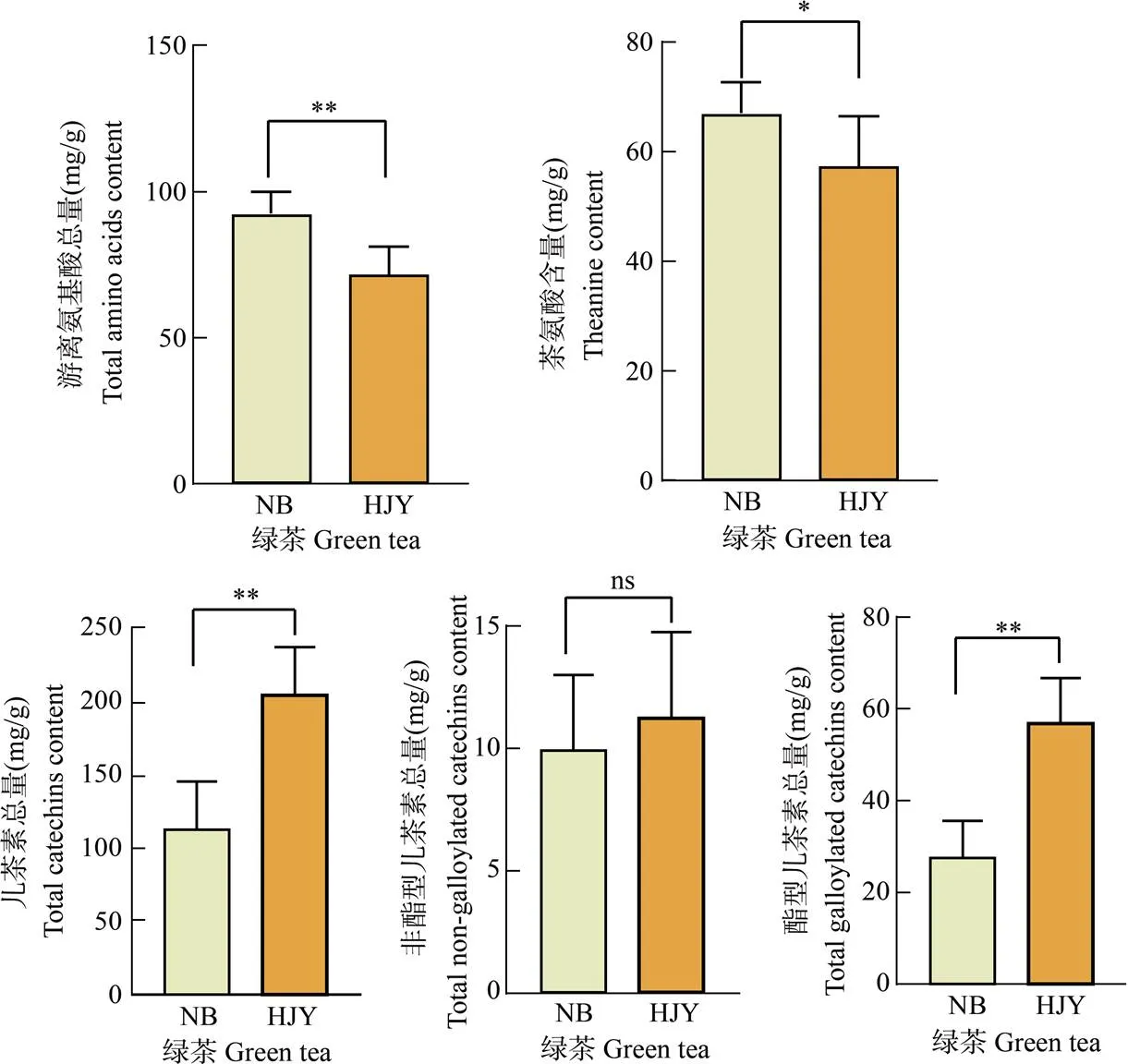
圖4 綠茶中滋味物質含量。*: P<0.05; **: P<0.01
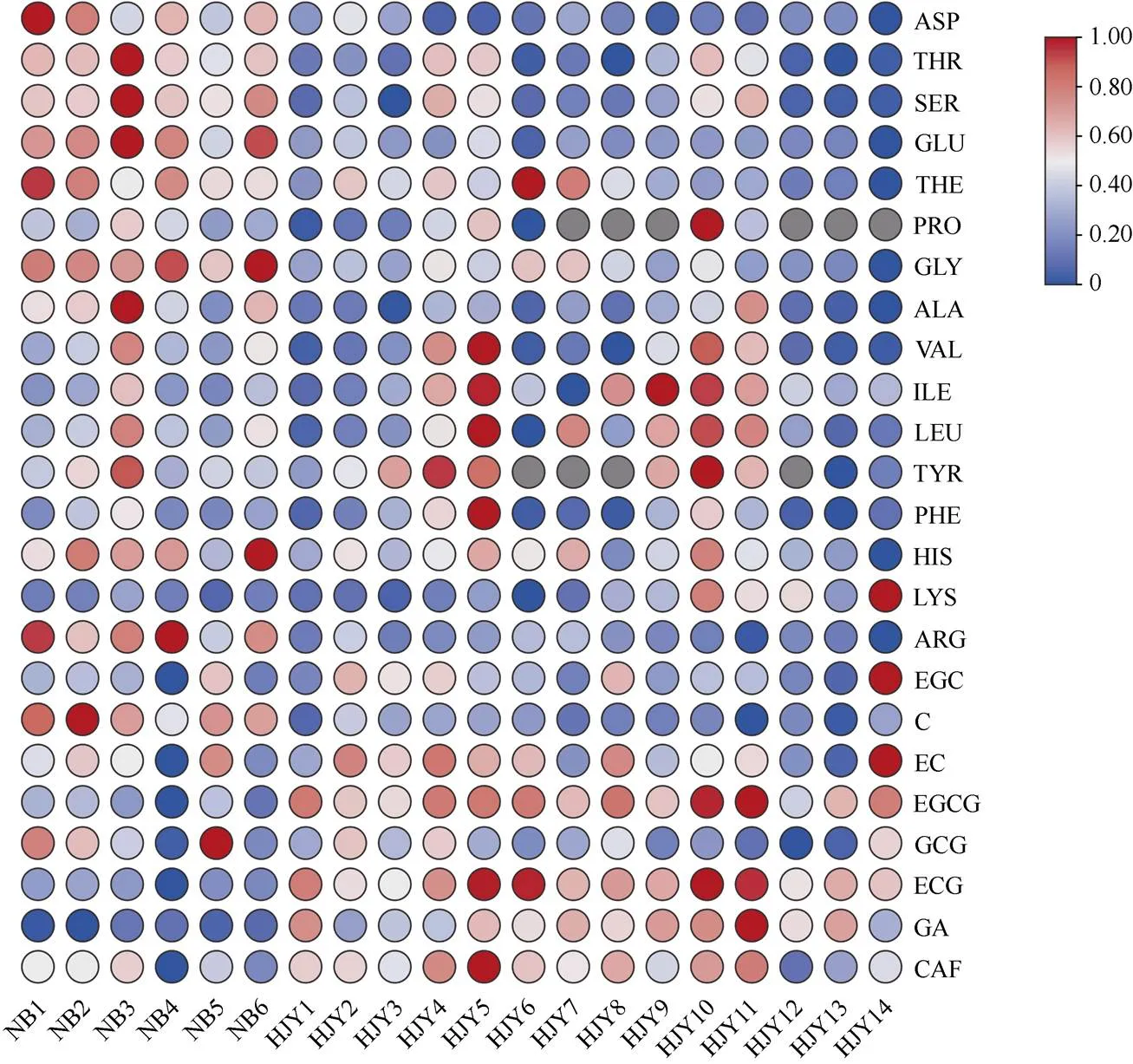
圖5 綠茶中游離氨基酸和兒茶素類化合物含量的熱圖分析。THE: 茶氨酸; ASP: 天冬氨酸; THR: 蘇氨酸; SER: 絲氨酸; GLU: 谷氨酸; GLY: 甘氨酸; ALA: 丙氨酸; VAL: 纈氨酸; ILE: 異亮氨酸; LEU: 亮氨酸; PHE: 苯丙氨酸; HIS: 組氨酸; LYS: 賴氨酸; ARG: 精氨酸; EGC: 表沒食子兒茶素; C: 兒茶素; EC: 表兒茶素; EGCG: 表沒食子兒茶素沒食子酸酯; GCG: 沒食子兒茶素沒食子酸酯; ECG: 表兒茶素沒食子酸酯: GA: 沒食子酸; CAF: 咖啡堿。下同

圖6 綠茶中游離氨基酸和兒茶素類化合物的PLS-DA分析(A, B)和VIP值(C, D)

表3 Dot值>1的化合物
3 結論和討論
本研究將奶白茶(白化品種)和黃金芽茶(黃化品種)的茶湯滋味分為鮮度、濃度、強度、協調度4個因子,通過感官審評進行打分量化滋味特征,結果表明二者在茶湯滋味上存在差異。兩種綠茶在茶湯滋味的鮮度和強度(收斂性)明顯不同,奶白茶茶湯滋味的鮮爽度高于黃金芽,而黃金芽茶茶湯滋味的收斂性強于奶白茶。進一步檢測了導致二者滋味差異的關鍵化合物,結果表明奶白茶中游離氨基酸含量顯著高于黃金芽茶,其具有不同的呈味特性[24],呈現鮮味和甜味的游離氨基酸含量差異最顯著,如茶氨酸、天冬氨酸、谷氨酸,蘇氨酸、絲氨酸,且最主要的差異化合物是茶氨酸,其在茶湯滋味中貢獻鮮味。貢獻澀味和苦味的兒茶素類化合物和咖啡堿含量在奶白茶中顯著低于黃金芽茶,且主要差異化合物為酯型兒茶素EGCG、ECG和咖啡堿,酯型兒茶素具有較強的苦澀味,收斂性強,是茶湯澀味的重要來源[25]。這些滋味成分的差異可能是導致奶白茶茶湯更加鮮爽而黃金芽茶茶湯收斂性更強的主要因素。
茶湯的滋味是多種化合物綜合協調的結果,不同滋味化合物在茶湯中的含量和呈味閾值不同,因此對茶湯滋味品質的貢獻也存在明顯差異[26]。Dot值反映對兩種綠茶茶湯滋味起主要貢獻的是EGCG和咖啡堿。黃金芽茶中EGCG和ECG的Dot值顯著高于奶白茶,可能是導致黃金芽茶湯比奶白茶茶湯收斂性更強、口感更厚的重要化合物。沒食子酸在黃金芽茶中的Dot值也明顯高于奶白茶,其在奶白茶中的Dot值均小于1,而在黃金芽茶中均大于1。Chen等[27]的研究表明沒食子酸能夠增強茶湯中EGCG的收斂性,因此,沒食子酸含量的差異可能對茶湯的收斂性產生影響。
研究表明,葉綠體的缺失使得參與光合作用和碳固定途徑的基因表達被下調,影響了植物體內的碳氮平衡,促進積累更多的氨基酸[13]。另有研究表明,兒茶素類化合物在白化茶樹白化期下降到最低水平[22],且白化、黃化茶樹品種的葉色變化與苯丙烷代謝、類黃酮和類胡蘿卜代謝變化密切相關[3,28],這表明茶樹葉片葉色的改變影響了內源代謝物的積累,如兒茶素的合成,使得所制綠茶滋味發生改變。
綜上所述,白化和黃化品種茶樹特征代謝物積累存在差異,比如游離氨基酸、兒茶素類化合物、沒食子酸和咖啡堿等,使得其鮮葉加工的綠茶滋味有差異。后續研究中,可進一步分析主要滋味成分的代謝途徑在2種茶樹品種中的差異。
[1] WANG S L, MA C L, HUANG D J, et al. Analysis of genetic diversity and construction of DNA fingerprints of chlorophyll-deficient tea cultivars by SSR markers [J]. J Tea Sci, 2018, 38(1): 58–68. [王松琳, 馬春雷, 黃丹娟, 等. 基于SSR標記的白化和黃化茶樹品種遺傳多樣性分析及指紋圖譜構建 [J]. 茶葉科學, 2018, 38(1): 58–68. doi: 10.3969/j.issn.1000-369X.2018.01.006.]
[2] LOU Y H, HE W Z, LIU Y, et al. Comprehensive assessment of quality traits of 14 etiolated and albino tea cultivars [J]. J Tea, 2020, 46(2): 84–90. [婁艷華, 何衛中, 劉瑜, 等. 14個黃化、白化變異茶樹品種(系)綜合性狀評價與分析 [J]. 茶葉, 2020, 46(2): 84–90. doi: 10.3969/j. issn.0577-8921.2020.02.004.]
[3] SONG L B, MA Q P, ZOU Z W, et al. Molecular link between leaf coloration and gene expression of flavonoid and carotenoid biosyn- thesis ins cultivar ‘Huangjinya’ [J]. Front Plant Sci, 2017, 8: 803. doi: 10.3389/fpls.2017.00803.
[4] WANG W, GUO Y L. Development and application of albino tea varieties [J]. J Food Safety Quality, 2017, 8(8): 3104–3110. [王蔚, 郭雅玲. 白化茶品種的開發與應用 [J]. 食品安全質量檢測學報, 2017, 8(8): 3104–3110. doi: 10.3969/j.issn.2095-0381.2017.08.044.]
[5] DU Y Y, SHIN S, WANG K R, et al. Effect of temperature on the expression of genes related to the accumulation of chlorophylls and carotenoids in albino tea [J]. J Hort Sci Biotechnol, 2009, 84(3): 365– 369. doi: 10.1080/14620316.2009.11512533.
[6] FENG L, GAO M J, HOU R Y, et al. Determination of quality constituents in the young leaves of albino tea cultivars [J]. Food Chem, 2014, 155: 98–104. doi: 10.1016/j.foodchem.2014.01.044.
[7] LI C F, MA J Q, HUANG D J, et al. Comprehensive dissection of metabolic changes in albino and green tea cultivars [J]. J Agric Food Chem, 2018, 66(8): 2040–2048. doi: 10.1021/acs.jafc.7b05623.
[8] BO J H, ZHANG Y L, GONG L J, et al. Research progress on secondary metabolites of albino tea from the perspective of multiomics [J]. Mol Plant Breeding, 2023, 21(6): 1884–1889. [薄佳慧, 張楊玲, 宮連瑾, 等. 多組學視角下白化茶樹次生代謝產物的研究進展 [J]. 分子植物育種, 2023, 21(6): 1884–1889. doi: 10.13271/j.mpb.021. 001884.]
[9] ZHANG Y N, JI W B, XU Y Q, et al. Rewiew on taste characteristic of catechins and its sensory analysis method [J]. J Tea Sci, 2017, 37(1): 1–9. [張英娜, 嵇偉彬, 許勇泉, 等. 兒茶素呈味特性及其感官分析方法研究進展 [J]. 茶葉科學, 2017, 37(1): 1–9. doi: 10.3969/j.issn. 1000-369X.2017.01.001.]
[10] ZHANG L, CAO Q Q, GRANATO D, et al. Association between chemistry and taste of tea: A review [J]. Trends Food Sci Technol, 2020, 101: 139–149. doi: 10.1016/j.tifs.2020.05.015.
[11] YIN J F, ZHANG Y N, DU Q Z, et al. Effect of Ca2+concentration on the tastes from the main chemicals in green tea infusions [J]. Food Res Int, 2014, 62: 941–946. doi: 10.1016/j.foodres.2014.05.016.
[12] KANEKO S, KUMAZAWA K, MASUDA H, et al. Molecular and sensory studies on the umami taste of Japanese green tea [J]. J Agric Food Chem, 2006, 54(7): 2688–2694. doi: 10.1021/jf0525232.
[13] LU M Q, HAN J Y, ZHU B Y, et al. Significantly increased amino acid accumulation in a novel albino branch of the tea plant () [J]. Planta, 2019, 249(2): 363–376. doi: 10.1007/s00425-018- 3007-6.
[14] CHENG H, LI S F, CHEN M, et al. Physiological and biochemical essence of the extraordinary characters of Anji Baicha [J]. J Tea Sci, 1999(2): 87–92. [成浩, 李素芳, 陳明, 等. 安吉白茶特異性狀的生理生化本質 [J]. 茶葉科學, 1999(2): 87–92. doi: 10.3969/j.issn.1000- 369X.1999.02.002.]
[15] DONG F, ZENG L T, YU Z M, et al. Differential accumulation of aroma compounds in normal green and albino-induced yellow tea () leaves [J]. Molecules, 2018, 23(10): 2677. doi: 10. 3390/molecules23102677.
[16] LI N N, LU J L, LI Q S, et al. Dissection of chemical composition and associated gene expression in the pigment-deficient tea cultivar ‘Xiaoxueya’ reveals an albino phenotype and metabolite formation [J]. Front Plant Sci, 2019, 10: 1543. doi: 10.3389/fpls.2019.01543.
[17] ZHANG X, CHEN X J, DU X, et al. Analysis on taste characteristics and main component contribution of Mengding Ganlu tea [J] J Yunnan Univ (Nat Sci), 2020, 42(4): 783–791. [張翔, 陳學娟, 杜曉, 等. 蒙頂甘露茶滋味特征及主要呈味成分貢獻率分析 [J]. 云南大學學報(自然科學版), 2020, 42(4): 783–791. doi: 10.7540/j.ynu.20190404.]
[18] ZHOU H C, WANG H, LEI P D, et al. Analysis of aromatics, catechins and free amino acids in different strains of ‘Shida Tea’ [J]. J Trop Subtrop Bot, 2018, 26(3): 302–308. [周漢琛, 王輝, 雷攀登, 等. ‘柿大茶’品系間芳香類物質、兒茶素及游離氨基酸差異分析 [J]. 熱帶亞熱帶植物學報, 2018, 26(3): 302–308. doi: 10.11926/jtsb.3809.]
[19] ZHOU H C, LIU Y Q, LEI P D. Metabolites profiling of green tea processed from ‘Huangshanbaicha No. 1’ cultivar at different albino stages [J]. J Trop Subtrop Bot, 2022, 30(2): 187–194. [周漢琛, 劉亞芹, 雷攀登. 不同白化期的‘黃山白茶’代謝物差異分析 [J]. 熱帶亞熱帶植物學報, 2022, 30(2): 187–194. doi: 10.11926/jtsb.4461.]
[20] LIU Y Q, ZHOU H C, WANG H, et al. Dynamic changes of cellulase and main quality components in processing of black tea [J]. Sci Technol Food Ind, 2020, 41(6): 66–70. [劉亞芹, 周漢琛, 王輝, 等. 紅茶加工過程中纖維素酶和主要品質成分的動態變化 [J]. 食品工業科技, 2020, 41(6): 66–70. doi: 10.13386/j.issn1002-0306.2020.06. 011.]
[21] ZHANG Y, DU X, WANG X S. Research progress of theanine in tea () [J]. Food Res Dev, 2007, 28(11): 170–174. [張瑩, 杜曉, 王孝仕. 茶葉中茶氨酸研究進展及利用前景[J]. 食品研究與開發, 2007, 28(11): 170–174. doi: 10.3969/j.issn.1005-6521.2007.11. 052.]
[22] SCHARBERT S, HOFMANN T. Molecular definition of black tea taste by means of quantitative studies, taste reconstitution, and omission experiments [J]. J Agric Food Chem, 2005, 53(13): 5377–5384. doi: 10. 1021/jf050294d.
[23] HUFNAGEL J C, HOFMANN T. Quantitative reconstruction of the nonvolatile sensometabolome of a red wine [J]. J Agric Food Chem, 2008, 56(19): 9190–9199. doi: 10.1021/jf801742w.
[24] TSENG Y H, LEE Y L, LI R C, et al. Non-volatile flavour components of[J]. Food Chem, 2005, 90(3): 409–415. doi: 10. 1016/j.foodchem.2004.03.054.
[25] XIONG L G, LI J, LI Y H, et al. Dynamic changes in catechin levels and catechin biosynthesis-related gene expression in albino tea plants (L.) [J]. Plant Physiol Bioch, 2013, 71: 132–143. doi: 10.1016/j.plaphy.2013.06.019.
[26] SHEN Q, ZHANG X Q, XU F F, et al. Changes of odorous substance and taste attributes of the Zheng’an Baicha in different times [J]. Sci Technol Food Ind, 2020, 41(24): 31–35. [沈強, 張小琴, 許凡凡, 等. 不同時期正安白茶呈味物質變化及滋味評價 [J]. 食品工業科技, 2020, 41(24): 31–35. doi: 10.13386/j. issn1002-0306.2020030278.]
[27] CHEN Y H, ZHANG Y H, CHEN G S, et al. Effects of phenolic acids and quercetin-3--rutinoside on the bitterness and astringency of green tea infusion [J]. NPJ Sci Food, 2022, 6(1): 8. doi: 10.1038/s41538- 022-00124-8.
[28] MA Q P, LI H, ZOU Z W, et al. Transcriptomic analyses identify albino-associated genes of a novel albino tea germplasm ‘Huabai 1’ [J]. Hort Res, 2018, 5(1): 54. doi: 10.1038/s41438-018-0053-y.
Differences of Free Amino Acids, Catechins and Caffeine Between Albino and Etiolated Tea Varieties
XU Yujie1, DAI Xiaojing2, WU Jizhong2, LIU Yaqin1, ZHOU Hanchen1, YANG Jihong1, WANG Hui1, HUANG Jianqin1, DING Yong1, LEI Pandeng1*
(1. Tea Research Institute, Anhui Academy of Agricultural Sciences,Huangshan 245000, Anhui, China; 2. Guangde Agricultural and Rural Bureau,Xuancheng 242200, Anhui, China)
Green teas processed from shoots of albino tea varieties have an umami taste and premium qualities. In order to explore the taste quality and metabolite differences of green tea made from fresh leaves of albino and etiolated tea varieties, sensory evaluation and metabolite analysis were studied on 6 albino green tea varieties (milk white tea, NB) and 14 etiolated green tea varieties (golden tea, HJY) in Guangde City. Sensory evaluation showed that NB green teas taste fresh and convergence was slightly weak; the taste of HJY green tea had strong convergence and the freshness was lower than that of NB green tea. The total amount of free amino acids and the content of free amino acids with fresh and sweet taste in NB green teas were significantly higher than those in HJY green teas, while the contents of catechinsand gallic acids contributing to convergence and the contents of bitter caffeine in NB green tea were significantly lower than those in HJY green teas. The partial least squares discriminant analysis showed that there were 7 signature compounds leading to the taste difference between the two green teas, which were theanine, arginine, glutamic acid, aspartate, epigallocatechin gallate (EGCG), epicatechin gallate and theobromine. The contribution of EGCG with the highest dose-over-threshold (Dot) to the flavor of HJY green tea was significantly higher than that of NB green tea. Therefore, the differences in contents of free amino acids, catechins, gallic acids and theophylline caused the different taste of albino and etiolated tea varieties.
; Albino tea; Etiolated tea; Green tea; Taste compound
10.11926/jtsb.4642
2022-03-28
2022-07-09
安徽省農業科學院青年英才計劃(QNYC-202119);安徽省“廣德黃金芽茶產業+金融+科技”發展試點項目;國家現代農業產業技術體系建設專項(CARS-19)資助
This work was supported by the Project for Youth Talent in Anhui Academy of Agricultural Sciences (Grant No. QNYC-202119), the Project for “Industry + Finance + Technology of Guangde Huangjinya Tea” Development Pilot in Anhui, and the Special Project for Building the National Modern Agricultural Industrial Technology System (Grant No. CARS-19).
徐玉婕(1996年生),女,碩士研究生,研究方向為茶葉加工與品質化學。E-mail: xyujie96@163.com
. E-mail: lpteagle@126.com
