生態恢復方式對喀斯特生境苔蘚植物物種組成的影響
2023-10-17 06:14:06汪鵬魏虹陳明月張梅曾辰武張學朝
熱帶亞熱帶植物學報 2023年5期
汪鵬, 魏虹, 陳明月, 張梅, 曾辰武, 張學朝
生態恢復方式對喀斯特生境苔蘚植物物種組成的影響
汪鵬, 魏虹*, 陳明月, 張梅, 曾辰武, 張學朝
(三峽庫區生態環境教育部重點實驗室,重慶市三峽庫區植物生態與資源重點實驗室,西南大學生命科學學院,重慶 400715)
為探究生態恢復方式對苔蘚植物的影響,研究了自然棄耕和退耕還林等生態恢復方式對重慶市中梁山喀斯特地區不同生境類型下苔蘚植物物種組成特征的影響。結果表明,研究區有苔蘚植物15科26屬41種,其中蘚類39種,苔類2種,美灰蘚()為主要優勢物種;研究區石生蘚類植物占比較大,生活型以平鋪型和叢集型為主,各生境物種多樣性及相似性指數整體偏低,但相比棄耕雜草地、灌木林和經濟型林地,竹林和馬尾松林中苔蘚植物具有更高的物種多樣性;冗余分析表明,郁閉度和光照強度是影響苔蘚植物分布的關鍵環境因子,喬木林為苔蘚植物的生長提供了更適宜的生境。因此,相比于自然恢復和經濟性林地等恢復方式,退耕還林是喀斯特退化生境更為理想的生態恢復方式。
喀斯特生境;生態恢復方式;苔蘚植物;生物多樣性
喀斯特生境是一種脆弱的生態環境,具有環境容量低、生物量小、生態環境系統變異敏感度高、空間轉移能力強、穩定性差等一系列生態脆弱性特征[1]。人為干擾是喀斯特地區生態環境惡化的主要驅動力,砍伐、勞作等人為干擾破壞了自然生態系統,造成植被退化、水土流失、石漠化等問題的發生和發展[2]。近年來,眾多學者開展了大量關于石漠化退化生態系統植被恢復的研究,生態恢復可以通過構建不同植被類型[3],改善土壤結構、養分條件和微生物菌群等環境條件[4],從而有效防止喀斯特生態系統繼續退化并提升其生態服務功能。研究表明,不同生態恢復方式的恢復效果存在較大差異,有待進行更深入的研究[5]。
植物群落是物種與環境相互作用的產物。復雜的環境條件對物種的生長繁殖具有選擇性和一定的限制性,恢復后形成的植物群落又會影響原有的生境條件[6]。相較于喬灌草類植物,大多數苔蘚植物葉僅1層細胞,且表面無保護性的角質層,對環境變化具有更高的敏感性,對環境變化的生態響應(生長基質、生活型、物種分布及組成)也更明顯[7]。但相較于喬灌草類植物,苔蘚植物受個體小、鑒別困難等原因的限制,在研究中往往被忽略[8]。作為陸地生態系統中重要的生產者,苔蘚植物分布廣泛,部分物種生命力頑強,能夠在極端環境中生存[9]。很多苔蘚植物對干燥環境有極強的適應性,能生活在裸露的巖石和沙丘上,是石漠化地區生態恢復的先鋒植物和拓荒者[10],在生成巖面生物結皮層、改善巖面環境和促進群落演替過程中起著重要作用[11]。因此,喀斯特地區苔蘚植物的生態學功能越來越成為研究者關注的焦點。
重慶中梁山是以褶皺束構造為背景的低山丘陵地貌,多為砂巖組成的脊狀山脊線,背斜低山中部出露有灰巖的地區,形成細長的與山體走向一致的巖溶槽谷,具有典型的喀斯特地貌特征[12]。1960年前后,人為因素造成該區域大量森林砍伐殆盡, 生境退化嚴重。1996年重慶中梁山區域開始進行石漠化治理,形成了由退耕還林恢復的剛竹林、慈竹林及馬尾松林等林地和自然棄耕恢復的灌木、雜草地等恢復生境[13]。本研究以中梁山喀斯特生境苔蘚植物群落為研究對象,探究不同的生態恢復方式對苔蘚植物群落物種組成特征的影響,以期更好地了解和掌握喀斯特地區生態恢復的狀況并指導生態恢復實踐。
1 材料和方法
1.1 研究區概況
研究區域位于重慶市近郊北碚區中梁山龍鳳槽谷鳳凰村附近,屬亞熱帶濕潤季風氣候區,冬暖夏熱,雨量豐沛,常年平均氣溫16.8 ℃,年降水量為1 200~1 300 mm,海拔360~700 m[14]。該地區開展退耕還林工程后,在山體兩側恢復形成了連片的馬尾松()林,并有部分柏木()林和刺槐()林;在居民區周邊及山腳則為栽種的剛竹()和慈竹()林。灌木林與草地多為農田棄耕后自然恢復形成,其中灌木層以白櫟()、女貞()幼苗占優勢;草本層植物有白茅()、鐵芒萁()等。區內人類活動以農業生產活動為主,主要栽種包括獼猴桃()、核桃()等經濟林木和玉米()、土豆()等農作物[15]。
1.2 樣地及樣方設定
參照相關研究對退化生境恢復方式的劃分方法進行樣地選擇[16–17]。經野外調查,確定了棄耕地、灌木林地、馬尾松林、竹林和獼猴桃林5種生境, 分別對應自然棄耕、退耕還林和人工經濟林等不同恢復方式,涵蓋了研究地區主要生境類型。每種生境各選擇典型樣地3個(表1),樣地面積均為10 m×20 m。調查樣地地理坐標為(106°25′29.97″~106°25′56.61″ E,29°46′06.39″~29°46′32.74″ N)。棄耕地為棄耕后自然恢復2~5 a的混生雜草地,主要分布狗尾草、青蒿、烏蘞梅、接骨木等。灌木林地為棄耕后自然恢復5~10 a形成的灌草混生群落,主要分布白櫟、欒樹(幼)、女貞(幼)、懸鉤子、小飛蓬等。馬尾松林為退耕還林人工恢復約20 a左右,建群種為馬尾松林,林下雜生棕櫚、刺五加、懸鉤子等。竹林為退耕還林人工恢復約20 a左右,建群種為剛竹,散生極少數女貞、黃檀,林下幾乎無其他植被。獼猴桃林經濟型林地,林下分布小飛蓬、馬唐、狗尾草等,栽種10 a左右。
參照《生物多樣性觀測技術導則》[18]設置苔蘚植物樣方,每個樣地均設置9個樣方。由于研究區苔蘚植物主要分布在大小不一的巖石表面,且平鋪生長的蘚類長勢較好,本研究將每個樣方大小調整為20 cm×20 cm,以便更好地比較不同基質上苔蘚植物蓋度差異。為反映苔蘚植物群落對干旱生境的適應,于2020年6月—8月重慶夏季高溫期(氣溫> 35 ℃)開展了3次野外調查, 每次調查5個樣地(各生境類型1個),共調查15個樣地,合計135個樣方。
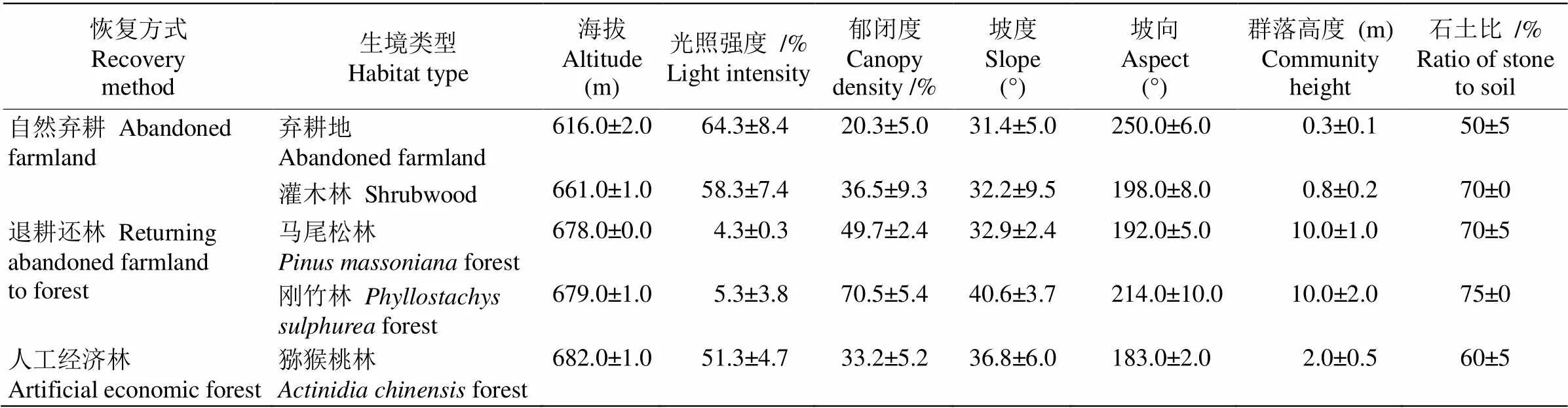
表1 樣地的環境特征(n=3)
1.3 環境因子測定及植物樣本采集
為盡量減少溫度和濕度的影響,在晴朗天氣對樣地進行采樣。采樣前,將GPS (HOLUX,m-241plus)校準后直立調查樣方中心位置,待穩定后讀數記錄,獲取樣地經緯度、海拔信息;將兩個照度計(EXTECH,LT45)分別放置于調查樣方中心位置及樣地外全光照處,同時記錄3次讀數,通過樣地內光照與全光照比值獲取相對光照數據;利用羅盤獲取樣地坡度、坡向等信息;利用目測法估算各個樣地的其他高等植物的群落高度和郁閉度。將樣方中出現的每種苔蘚植物編號,利用篩網(篩孔1 cm× 1 cm)測定各物種蓋度,記錄其生長基質后采樣裝袋,共采集植物樣本300余份。
1.4 樣本鑒定
將樣本自然風干保存后進行形態解剖,利用SZ810體式顯微鏡及BK6000生物顯微鏡對采集的苔蘚植物的細胞、葉形、葉尖、孢子體、葉基、中肋等形態進行觀察和鑒定。憑證樣本保存在西南大學三峽庫區生態環境教育部重點實驗室。
1.5 物種多樣性計算
由于苔蘚植物個體數目不易測量,因此以苔蘚植物的蓋度代替多度進行計算,本研究參考張金屯主編的《數量生態學》[19]進行物種多樣性分析。重要值()體現了物種在群落中的重要性,=()/2,式中,為苔蘚植物的生態重要值,為相對蓋度;為相對頻度。
多樣性是反映群落內部物種數和物種相對多度的一個指標,主要表明群落本身的物種組成和個體數量分布的特征。本試驗選取Patrick豐富度指數()、Simpson多樣性指數()、Shannon-Wiener多樣性指數()和Pielou均勻度指數()。Patrick指數,Simpsom指數=1-Σ(N/)2, Shannon-Wiener指數=-Σ(P×lnP), Pielou指數=/ln(), 式中,為物種數,N為第個物種的蓋度,為所有物種的蓋度之和,P為第個物種的相對蓋度。
本試驗采用Sorenson多樣性指數衡量不同生境中苔蘚植物物種組成的變化,=2/(),式中,為兩個生境共有物種數,、分別為兩個生境的物種數。
1.6 數據處理
采用Excel對原始數據進行整理,利用SPSS軟件中單因素方差分析法分析不同生境中苔蘚植物多樣性指數的差異顯著性;利用CANOCO 4.5對物種和標準化后的環境數據進行冗余分析;利用R語言中mantel test檢驗環境因子對苔蘚物種分布的影響,具體為vagan包vedist()函數計算相似性距離,ape包中mantel()函數進行mantel test分析,其中物種數據使用Barry-Curtis距離,環境因子用Euclid距離。
2 結果和分析
2.1 物種組成特征
對野外調查獲取的300余份標本進行詳細鑒定,整個調查區域共有15科26屬41種苔蘚植物,其中蘚類39種,苔類2種。由圖1可知,研究區內叢蘚科(Pottiaceae)的屬、種數量最多,有7屬11種;其次為灰蘚科(Hypnaceae)有4屬4種、羽蘚科(Thui- diaceae)有3屬4種和青蘚科(Brachytheciaceae)有2屬6種,這些科苔蘚植物的分布較為廣泛。
對不同生境中苔蘚植物科屬種數量的統計結果表明,退耕還林的馬尾松林和竹林下苔蘚植物具有較高的科、屬、種數,而人工果園獼猴桃林下的苔蘚植物科屬種數較少(表2)。將苔蘚植物中物種數最多的科定義為優勢科,物種數最多的屬定義為優勢屬。除馬尾松林和灌木林中苔蘚植物優勢科分別為灰蘚科和真蘚科(Bryaceae)外,其他3種生境中苔蘚植物優勢科均為叢蘚科,青蘚屬()及扭口蘚屬()為優勢屬。

圖1 苔蘚植物的科屬種數
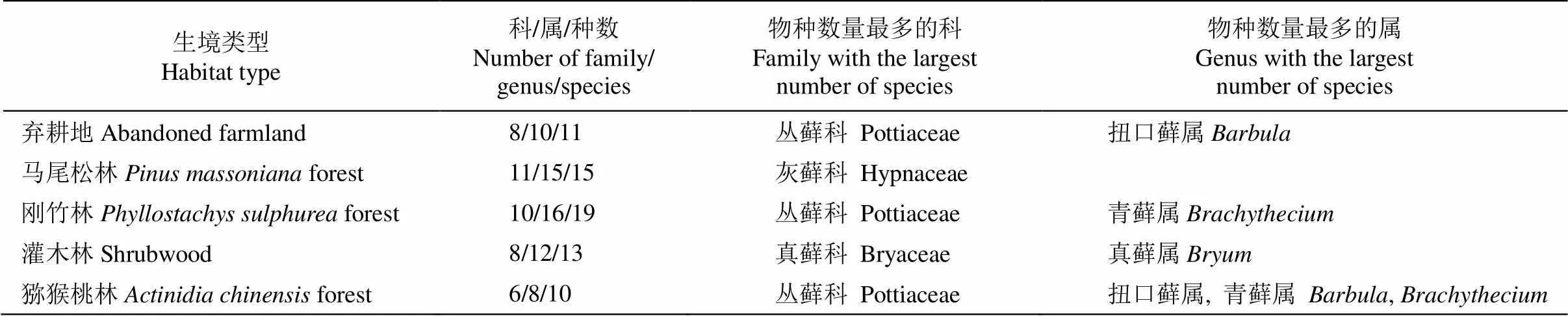
表2 不同生境中苔蘚植物科屬概況
本研究將樣地中重要值()≥0.10的苔蘚植物定義為優勢物種。由表3可知,不同生境中苔蘚植物優勢種和優勢程度均有較大差異,棄耕地、馬尾松林、竹林均有5種優勢物種,而灌木林和獼猴桃林分別有3和2種。其中美灰蘚在不同樣地中均占優勢, 但優勢程度具有一定差異,其在棄耕地、竹林、灌木林和獼猴桃林中均占主要優勢,而在馬尾松林中則是大灰蘚()占主要優勢。從物種生長方式來看,竹林中優勢苔蘚植物多以匍匐生長,不規則分枝為主,如美灰蘚、薄壁卷柏蘚();而其他生境中的優勢物種莖單一不分枝的蘚類則兼并叢集生長,如狹葉扭口蘚()、紅蒴立碗蘚()。美灰蘚在所有樣地中均表現出較好的適應性,具有較高的重要值。
2.2 生活型特征
從生長基質來看,中梁山喀斯特地區地表苔蘚植物有石生、土生和混生(同時生長在不同基質上) 3種。各生境均以混生苔蘚為主(占比≥50%);棄耕地中土生苔蘚植物占比較高(接近30%),而其他生境中土生苔蘚植物占比均相對偏低(圖2),這可能與樣地石土比較低有關,土壤面積占比較大為土生苔蘚提供了更多的生長環境。
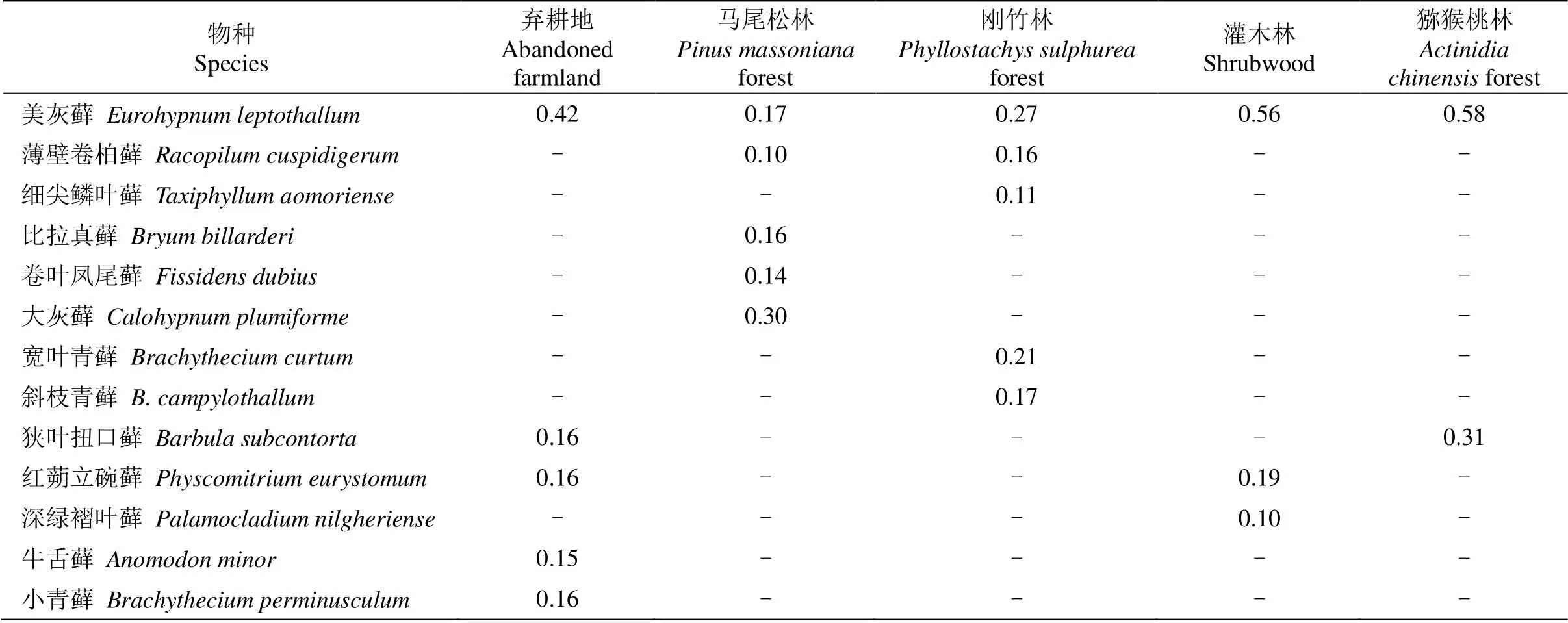
表3 部分優勢苔蘚植物的重要值

圖2 不同生境中苔蘚植物的生長基質。A: 棄耕地; B: 馬尾松林; C: 剛竹林; D: 灌木林; E: 獼猴桃林。下同
依據苔蘚植物生活型的劃分標準,地表苔蘚植物主要有3類生活型,即平鋪型、叢集型和交織型。由圖3可知,研究區苔蘚植物主要由平鋪型和叢集型組成,交織型較少。叢集型蘚類在棄耕地中最多,馬尾松林中最少,交織型并未在棄耕地中出現而在其他樣地中所占比例相近且均較低,而平鋪型蘚類植物在馬尾松林和竹林中占比較大,而在其余樣地中占比相近且較低。
2.3 苔蘚植物多樣性特征
5種植被類型中,馬尾松林、竹林和棄耕地中苔蘚植物豐富度顯著高于灌木林和獼猴桃林(0.05),整體趨勢為竹林>馬尾松林>棄耕地>灌木林>獼猴桃林;Simpsom和Shannon-Wiener多樣性指數的變化趨勢一致,均竹林>馬尾松林>棄耕地>獼猴桃林>灌木林,其中灌木林和獼猴桃林顯著低于其他3種類型;馬尾松林、竹林和棄耕地表現出較高的物種均勻度(圖4)。
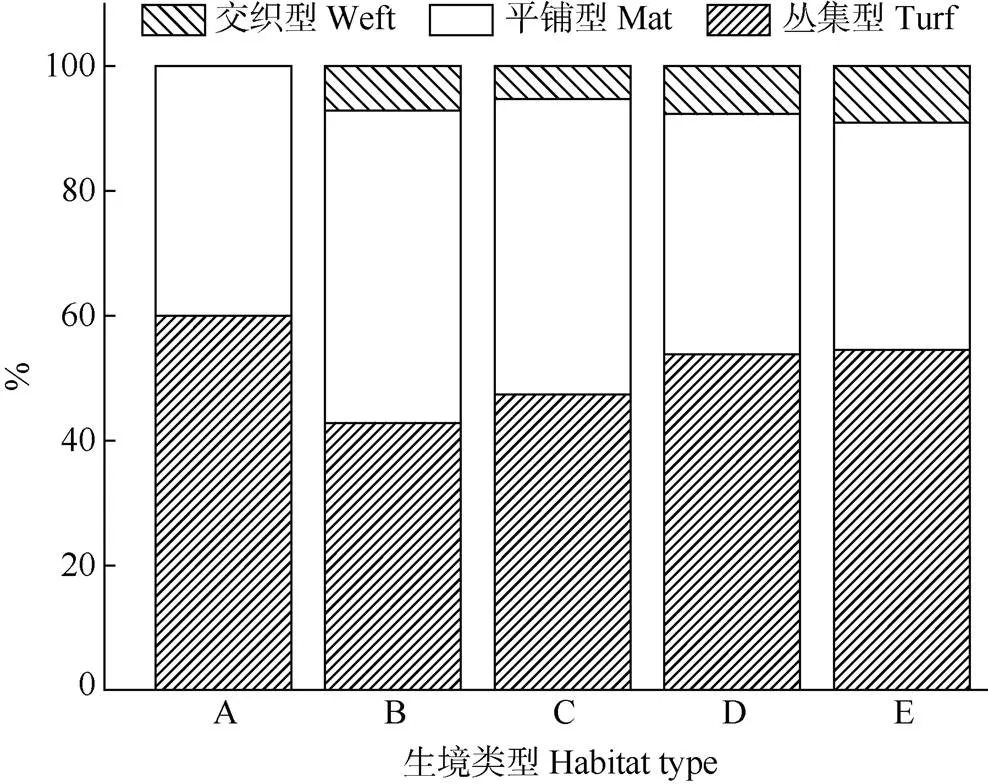
圖3 不同生境苔蘚植物生活型
比較各樣地中苔蘚植物物種組成的相似性,馬尾松林與灌木林中苔蘚植物相似性系數最高, 為0.429,呈中等相似水平;馬尾松與獼猴桃林相似性系數為0.240,呈極不相似水平;其他樣地間的相似性指數為0.250~0.400,屬于偏中低度不相似水平(表4),說明研究區苔蘚植物物種相似性整體偏低,不同恢復方式對樣地中苔蘚植物分布影響不同。
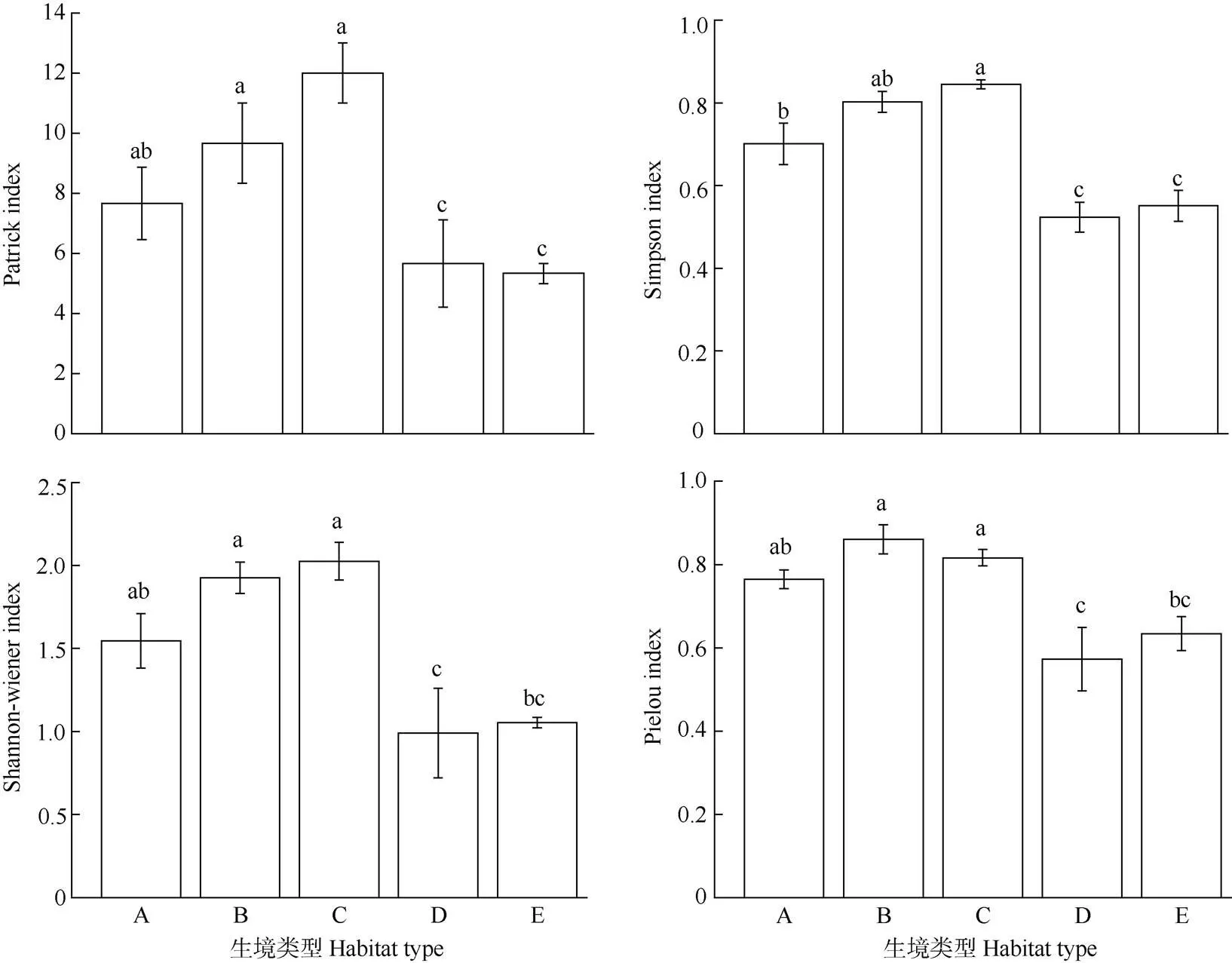
圖4 不同生境苔蘚植物的α多樣性指數。柱上不同字母表示樣地間差異顯著(P<0.05)。
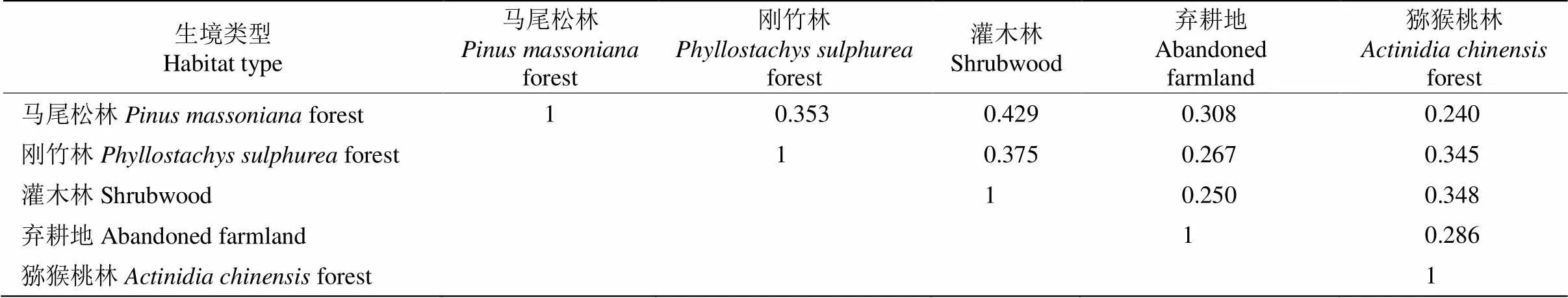
表4 不同生境苔蘚植物的Sorenson多樣性指數
2.4 環境因子對苔蘚植物分布的影響
為了更好地探究環境因子對不同恢復方式下苔蘚植物物種分布的影響,對樣地光照強度、郁閉度、海拔、坡度、坡向、群落高度等環境因子進行了共線性檢測,剔除了2>0.7且<0.05的環境因子(群落高度)。將研究區內頻度≥3的苔蘚物種與環境因子數據矩陣進行DCA分析,結果表明,第一軸梯度長度值為2.76,因此,本研究可采用線性模型進行進一步的RDA分析。RDA排序結果進行置換檢驗,顯著性小于0.05 (=0.002),說明排序結果可以接受環境因子對物種分布的解釋量(表5)。排序軸前4軸的方差解釋量為42.62%,累積方差解釋比例達95.75%;其中第一軸的特征值為0.179,第二軸的特征值為0.129。圖5為冗余分析后的RDA圖, 紅色箭頭代表環境變量,每個箭頭指向環境變量值最急劇增加的方向;箭頭之間的角度表示各環境變量間的相關性;通過將1個環境變量的箭頭投影到沿著該變量箭頭方向的假想線上,來讀取1個環境變量與其他環境變量的近似相關性。藍色箭頭代表物種,每個箭頭指向對應物種值的最大增加方向;箭頭之間的角度表示物種之間的相關性,當角度銳利時,近似相關性為正,當角度大于90°時,近似相關性為負;箭頭的長度是衡量該物種是否適合的標準。更定量地說,我們可以通過將其他物種的箭頭投射到覆蓋該物種箭頭的假想線上,來讀取一個物種與其他物種的近似相關性。如果投影線終止于坐標原點(零點),則相關性預計為零。通過將物種箭頭尖端垂直投影到覆蓋環境變量箭頭的線上,可以近似估算物種和環境變量之間的相關性。投影點在箭頭指示的方向上落得越遠,相關性越高,坐標原點(零點)附近的投影點表明相關性接近于零;如果投影點位于相反方向,則預測相關性為負。圖中的環境因子所對應的射線越長,表明該因子對研究對象的影響程度越大。由圖5可知,苔蘚植物分布與生境光照強度和郁閉度具有較強的相關性,美灰蘚()、密枝青蘚()、小青蘚()和扭口蘚()與光照相關性較大,與海拔、郁閉度呈負相關;而細尖鱗葉蘚、薄壁卷柏蘚和卷葉濕地蘚則與海拔和郁閉度相關性較大而與光照呈負相關。
采用Mantel檢驗環境因子對苔蘚植物分布的影響, 結果表明,光照對苔蘚植物分布具有極顯著影響,郁閉度對苔蘚植物分布具有顯著影響,而海拔、坡度、坡向對苔蘚植物分布的影響較小(表6)。
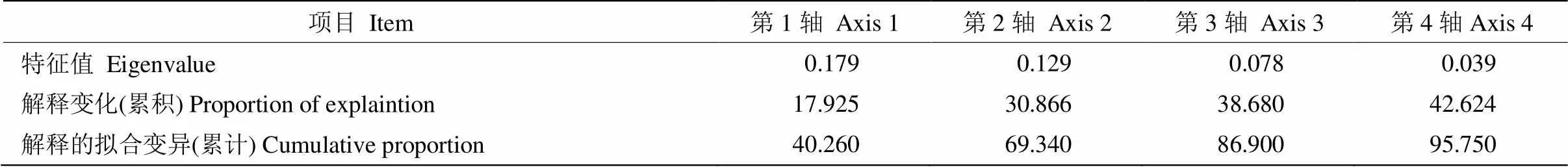
表5 冗余分析結果
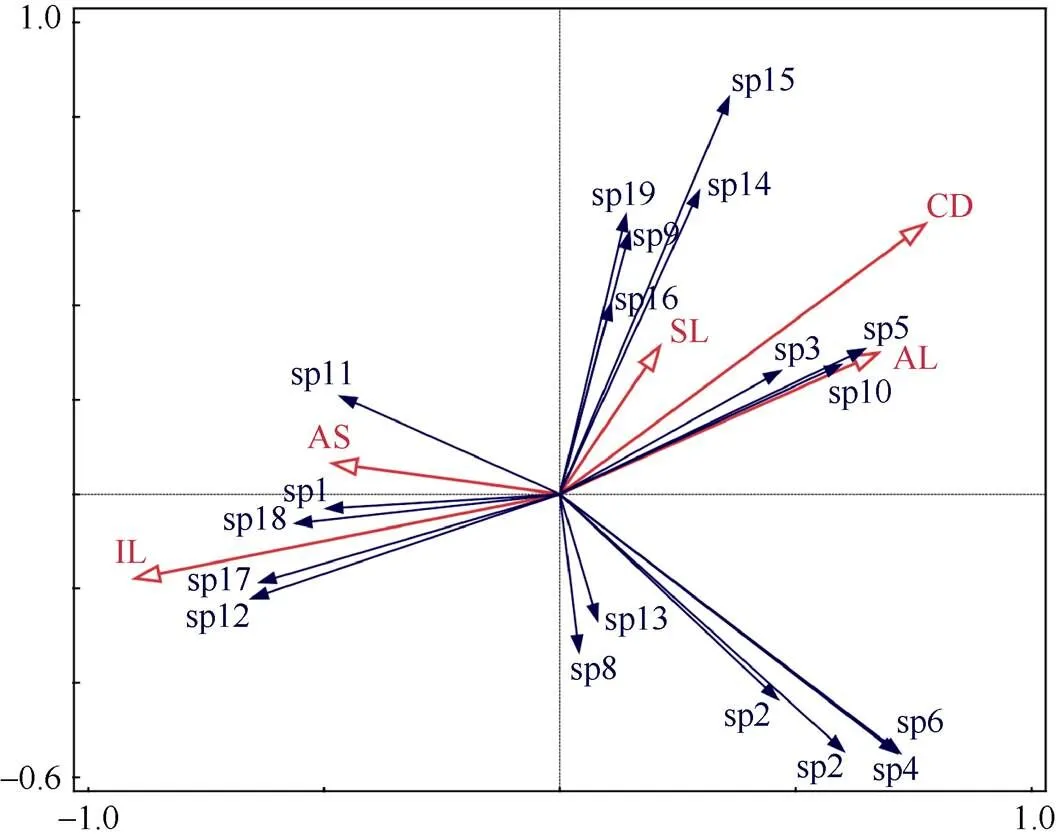
圖5 環境因子對苔蘚植物分布的影響。sp1: 美灰蘚; sp2: 粗枝蘚; sp3: 細尖鱗葉蘚; sp4: 大灰蘚; sp5: 薄壁卷柏蘚; sp6: 卷葉鳳尾蘚; sp7: 比拉真蘚; sp8: 毛口蘚; sp9: 皺葉毛口蘚; sp10: 卷葉濕地蘚; sp11: 狹葉扭口蘚; sp12: 扭口蘚; sp13: 多枝縮葉蘚; sp14: 寬葉青蘚; sp15: 斜枝青蘚; sp16: 密枝青蘚; sp17: 小青蘚; sp18: 紅蒴立豌蘚; sp19: 長柄絹蘚; IL: 光照強度; AS: 坡向; AL: 海拔; CD: 郁閉度; SL: 坡度。
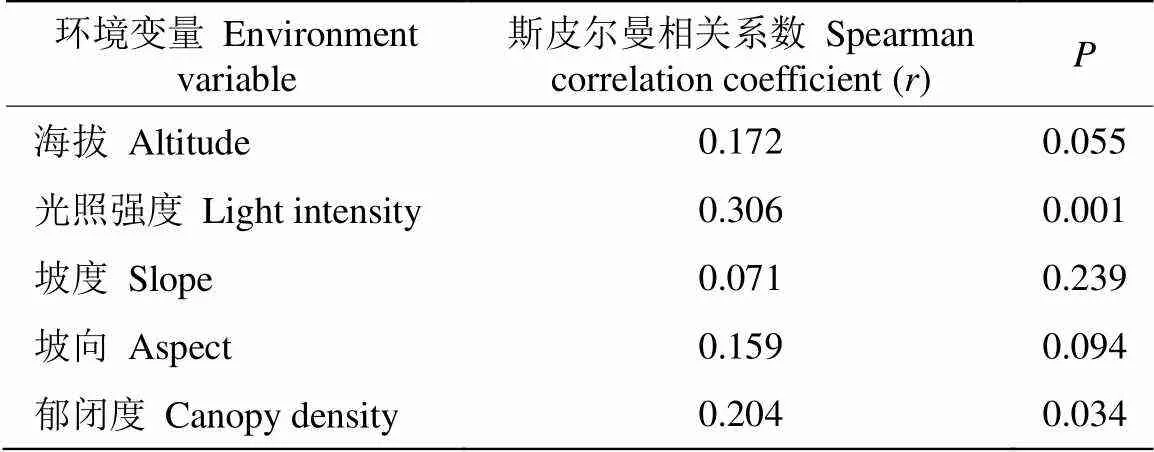
表6 Mantel test檢驗環境因子與苔蘚物種的相關性
3 結論和討論
喀斯特石漠化生境的特殊性,對植物種類具有強烈的篩選性(耐旱性、石生性)[20],本研究共調查到41種苔蘚植物。相較于其他喀斯特生境,如貴州溝谷[21]、猴耳天坑[22]等,本研究區苔蘚物種多樣性整體偏低,可能與樣地石漠化程度整體偏高有關(土石比50%~75%)[23]。苔蘚植物是群落演替的先鋒植物,對生境有較強的適應能力,研究區苔蘚植物生活型以叢集型和平鋪型為主,已形成了一套適應特殊環境的生活型和形態特征,可以更好地適應喀斯特干旱環境。叢集型苔蘚植物主要是通過密集生長,有效減緩空氣在葉片表面的流動,從而有利于降低植物體內水分的蒸騰[24];而平鋪型苔蘚植物具有更高的生物量和更好的吸水和持水能力[25],對石漠化生境的改善具有較好的生態意義。研究表明, 郁閉度較低的生境中,如灌木林、棄耕地和獼猴桃林中叢集型蘚類占比更大,說明叢集型蘚類可能更適應光照強度較高的生境。
在石漠化恢復過程中,由于采用了不同生態恢復方式,地上植被對石漠化環境產生了不同的影響[26]。本研究選取了自然棄耕、退耕還林和人工經濟林3種典型恢復方式,除海拔外的其他環境因子均表現出明顯差別,樣地間苔蘚植物相似性系數普遍偏低,在一定程度上反映了不同的恢復方式對生境具有不同的生態影響。而冗余分析表明,光照和郁閉度是影響研究區苔蘚植物分布的主要環境因子。光照為植物生長生存提供了必須的能源條件, 但是由于喀斯特地形環境復雜,植被類型多樣,導致光照強度差異明顯。而苔蘚植物由于光合色素含量等生理特征差異對光照條件表現出不同程度的耐受性,大部分苔蘚植物對光照的需求量低,能在低光照條件下完成滿足自身需求的光合作用。但也有部分物種對光照具有較高耐受性,分布于光強較大的區域[27–28]。郁閉度不僅可以影響群落中的光環境,還可以影響群落土壤水分及溫度,從而影響地被植物的生長發育[29],在喀斯特地區,水土保持能力相對其他生態系統較弱,郁閉度對群落微環境的影響更為顯著[30]。大量研究表明,郁閉度是影響苔蘚植物分布和物種多樣性的重要環境因子[31–33]。在本研究中,各生境巖石裸露率較高,巖面對水分的節流能力較弱,無法很好地為苔蘚植物提供生理水分,高郁閉度的竹林和馬尾松林能有效減少林下空氣流動,限制水分蒸發,因此對苔蘚植物分布與生存具有顯著影響。退耕還林的生態恢復方式為苔蘚植物提供了更理想的生境。
從重要值來看,不同樣地中苔蘚植物優勢物種不同,適應性較強的美灰蘚也表現出不同占比,表明不同的生態恢復措施影響了地被苔蘚植物的生長和分布。作為先鋒物種,苔蘚植物敏感地反映了生境的變化,不同物種對環境因子的適應體現出不同生態恢復方式的恢復效果差異。細尖鱗葉蘚、薄壁卷柏蘚可能更適應高郁閉度低光照環境,而狹葉扭口蘚,紅蒴立碗蘚則在低郁閉度高光照環境中廣泛存在,這可能與苔蘚植物對水分需求差異以及耐旱機制不同有關。相較于其他平鋪型蘚類,細尖鱗葉蘚和薄壁卷柏蘚的分枝密集和重疊程度較低,與空氣接觸的表面積較大,需要較高的濕度環境才能維持自身正常生長狀態,而狹葉扭口蘚和紅蒴立碗蘚則可能是因為個體相對較小,對水分需求量較低,因而在低郁閉度區域也能廣泛生長。
苔蘚植物在不同生境中的分布特點,直接影響各生境中苔蘚植物多樣性特征。在本研究中,不同恢復方式對苔蘚植物多樣性具有明顯影響,退耕還林形成的竹林和馬尾松林中苔蘚植物具有更高的多樣性,這與林地郁閉度較大,能有效保持林下空氣濕度、土壤濕度有關[34]。相對于棄耕后自然恢復形成的灌木林,棄耕農田苔蘚植物多樣性相對較高,這可能是與棄耕農田石土比相對較低,土生類苔蘚植物較多有關[35–37]。獼猴桃林中苔蘚植物物種數最低,可能是受到了較為強烈的人為活動的影響,施肥、農藥和耕地等農業活動,均可能在一定程度上影響苔蘚植物的生長環境,其內在聯系有待進行更深入的研究。而獼猴桃林中苔蘚植物多樣性指數和均勻度指數較灌木林高,主要是因為美灰蘚和狹葉扭口蘚優勢程度突出導致,進一步體現這兩個物種對研究區環境具有較強的適應性。
綜上,研究區不同恢復方式形成的生境中苔蘚植物物種組成、優勢種、生活型特征及多樣性指數均表現出一定的差異,說明不同的生態恢復方式對喀斯特地區苔蘚植物物種組成特征具有明顯影響。相比自然恢復和人工經濟林,退耕還林更適宜苔蘚植物的生長,更有助于提高該區域苔蘚植物多樣性。美灰蘚是研究區的主要優勢物種,對生境有更好的適應能力,可作為改善喀斯特環境的優先物種之一。
[1] SONG T Q, PENG W X, DU H, et al. Occurrence, spatial-temporal dynamics and regulation strategies of karst rocky desertification in southwest China [J]. Acta Ecol Sin, 2014, 34(18): 5328–5341. [宋同清,彭晚霞, 杜虎, 等. 中國西南喀斯特石漠化時空演變特征、發生機制與調控對策 [J]. 生態學報, 2014, 34(18): 5328–5341. doi: 10.5846/ stxb201405090929.]
[2] DAI Q H, YAN Y J. Research progress of karst rocky desertification and soil erosion in southwest China [J]. J Soil Water Conserv, 2018, 32(2): 1–10. [戴全厚, 嚴友進. 西南喀斯特石漠化與水土流失研究進展 [J]. 水土保持學報, 2018, 32(2): 1–10. doi: 10.13870/j.cnki. stbcxb.2018.02.001.]
[3] LIU Y G, LIU C C, WEI Y F, et al. Species composition and commu- nity structure at different vegetation successional stages in Puding, Guizhou Province, China [J]. Chin J Plant Ecol, 2011, 35(10): 1009– 1018. [劉玉國, 劉長成, 魏雅芬, 等. 貴州省普定縣不同植被演替階段的物種組成與群落結構特征 [J]. 植物生態學報, 2011, 35(10): 1009–1018. doi: 10.3724/SP.J.1258.2011.01009.]
[4] HU F, DU H, ZENG F P, et al. Dynamics of soil nutrient content and microbial diversity following vegetation restoration in a typical karst peak-cluster depression landscape [J]. Acta Ecol Sin, 2018, 38(6): 2170–2179. [胡芳, 杜虎, 曾馥平, 等. 典型喀斯特峰叢洼地不同植被恢復對土壤養分含量和微生物多樣性的影響 [J]. 生態學報, 2018, 38(6): 2170–2179. doi: 10.5846/stxb201703180458.]
[5] CHENG H T, LI Q F, WANG X M, et al. Effects of different vegetation restoration strategies on soil penetrability of karst ecosystem in Guizhou Province [J]. J Soil Water Conserv, 2020, 34(6): 110–116. [程漢亭, 李勤奮, 王曉敏, 等. 不同植被恢復策略對貴州喀斯特生態系統土壤滲透特性的影響 [J]. 水土保持學報, 2020, 34(6): 110–116. doi: 10.13870/j.cnki.stbcxb.2020.06.017.]
[6] LI Q K, MA K P. Advances in plant succession ecophysiology [J]. Acta Phytoecol Sin, 2002, 26(S1): 9–19. [李慶康, 馬克平. 植物群落演替過程中植物生理生態學特性及其主要環境因子的變化 [J]. 植物生態學報, 2002, 26(S1): 9–19.]
[7] CUTLER N. Nutrient limitation during long-term ecosystem develop- ment inferred from a mat-forming moss [J]. Bryologist, 2011, 114(1): 204–214. doi: 10.1639/0007-2745.114.1.204.
[8] YUAN C J, XIONG K N, RONG L, et al. Research progress on the biodiversity during the ecological restoration of karst rocky deserti- fication [J]. Earth Environ, 2021, 49(3): 336–345. [袁成軍, 熊康寧, 容麗, 等. 喀斯特石漠化生態恢復中的生物多樣性研究進展 [J]. 地球與環境, 2021, 49(3): 336–345. doi: 10.14050/j.cnki.1672-9250. 2021.49.061.]
[9] SARDANS J, PE?UELAS J. Drought changes nutrient sources, content and stoichiometry in the bryophyteHedw. growing in a Mediterranean forest [J]. J Bryol, 2008, 30(1): 59–65. doi: 10.1179/174328208X281987.
[10] KORANDA M, MICHELSEN A. Mosses reduce soil nitrogen availa- bility in a subarctic birch forest via effects on soil thermal regime and sequestration of deposited nitrogen [J]. J Ecol, 2021, 109(3): 1424– 1438. doi: 10.1111/1365-2745.13567.
[11] MENG W P, DAI Q H, RAN J C. A review on the process of bryophyte karstification [J]. Chin J Plant Ecol, 2019, 43(5): 396–407. [蒙文萍, 戴全厚, 冉景丞. 苔蘚植物巖溶作用研究進展 [J]. 植物生態學報, 2019, 43(5): 396–407. doi: 10.17521/cjpe.2019.0020.]
[12] WEI X P. The study on the characteristics and mechanism of soil erosion in karst valley area, Chongqing [D]. Chongqing: Southwest University, 2011. [魏興萍. 巖溶槽谷區水土流失特征及機理研究 [D]. 重慶: 西南大學, 2011.]
[13] ZHENG H R, LUO H X, ZOU Y Q, et al. Quantifying vegetation restoration in a karst rocky desertification area in Chongqing based on Geo-informatic Tupu [J]. Acta Ecol Sin, 2016, 36(19): 6295–6307. [鄭惠茹, 羅紅霞, 鄒揚慶, 等. 基于地學信息圖譜的重慶巖溶石漠化植被恢復演替研究 [J]. 生態學報, 2016, 36(19): 6295–6307. doi: 10. 5846/stxb201411122233.]
[14] ZHANG Y Z, JIANG Y J, LI Y, et al. Effects of tunnel excavation on slope runoff and soil erosion in a karst trough valley [J]. Acta Ecol Sin, 2019, 39(16): 6126–6135. [張遠矚, 蔣勇軍, 李勇, 等. 隧道工程對喀斯特槽谷區坡面產流及土壤侵蝕的影響 [J]. 生態學報, 2019, 39 (16): 6126–6135. doi: 10.5846/stxb201812052655.]
[15] WANG Q R, JIANG Y J, HAO X D, et al. Relationship between surface soil pollen and modern vegetation in karst trough area of Zhongliang Mountain, Chongqing City, SW China [J]. Carsol Sin, 2021, 40(3): 466–475. [汪啟容, 蔣勇軍, 郝秀東, 等. 重慶中梁山巖溶槽谷區表土孢粉與現代植被的關系研究 [J]. 中國巖溶, 2021, 40(3): 466–475. doi: 10.11932/karst2021y20.]
[16] YANG Z J, WEN C, YANG L, et al. Effects of different restoration methods on ecosystem multifunctionality in the semi-arid loess small watershed [J]. Acta Ecol Sin, 2020, 40(23): 8606–8617. [楊智姣, 溫晨,楊磊, 等. 半干旱黃土小流域不同恢復方式對生態系統多功能性的影響 [J]. 生態學報, 2020, 40(23): 8606–8617. doi: 10.5846/stxb2020 02010187.]
[17] ZHAO Y, ZHANG W, HU P L, et al. Responses of soil organic carbon fractions to different vegetation restoration in a typical karst depression [J]. Acta Ecol Sin, 2021, 41(21): 8535–8544. [趙元, 張偉, 胡培雷, 等. 桂西北喀斯特峰叢洼地不同植被恢復方式下土壤有機碳組分變化特征 [J]. 生態學報, 2021, 41(21): 8535–8544. doi: 10.5846/ stxb202101140151.]
[18] Ministry of Environmental Protection of the People’s Republic of China. HJ 710.2—2014 Technical guidelines for biodiversity monitoring: Lichens and bryophytes [S]. Beijing: China Environmental Press, 2015. [中華人民共和國環境保護部. HJ 710.2—2014 生物多樣性觀測技術導則地衣和苔蘚 [S]. 北京: 中國環境科學出版社, 2015.]
[19] ZHANG J T. Quantitative Ecology [M]. 2nd ed. Beijing: Science Press, 2011: 86–122. [張金屯. 數量生態學 [M]. 第2版. 北京: 科學出版社, 2011: 86–122.]
[20] LI J F, WANG Z H, ZHANG Z H. Bryophyte diversity and the effect of soil formation along with water conservation in karst rocky deserti- fication region [J]. Res Environ Sci, 2013, 26(7): 759–764. [李軍峰, 王智慧, 張朝暉. 喀斯特石漠化山區苔蘚多樣性及水土保持研究 [J]. 環境科學研究, 2013, 26(7): 759–764. doi: 10.13198/j.res.2013. 07.68.lijf.005.]
[21] TAN H Y. Study on bryophytes species diversity in karst ravine from Guizhou Province [D]. Guiyang: Guizhou University, 2017. [談洪英. 貴州喀斯特溝谷苔蘚植物物種多樣性研究 [D]. 貴陽: 貴州大學, 2017.]
[22] LI X F, ZHANG Z H, LI C Y, et al. Diversity of bryophytes in under- ground forest of Monkey-ear Tiankeng [J]. Acta Bot Boreali-Occid Sin, 2018, 38(12): 2324–2333. [李小芳, 張朝暉, 李承義, 等. 貴州猴耳天坑地下森林苔蘚植物多樣性特征研究 [J]. 西北植物學報, 2018, 38(12): 2324–2333. doi: 10.7606/j.issn.1000-4025.2018.12.2324.]
[23] TOBIAS M, NIINEMETS ü. Acclimation of photosynthetic charac- teristics of the mossto among-habitat and within- canopy light gradients [J]. Plant Biol, 2010, 12(5): 743–754. doi: 10. 1111/j.1438-8677.2009.00285.x.
[24] CONG C L, LIU T L, KONG X Y, et al. Flora and species diversity of epilithic mosses on rock desertification in the Puding Karst Area [J]. Carsol Sin, 2017, 36(2): 179–186. [從春蕾, 劉天雷, 孔祥遠, 等. 貴州普定喀斯特受損生態系統石生蘚類植物區系及物種多樣性研究 [J]. 中國巖溶, 2017, 36(2): 179–186. doi: 10.11932/karst20170204.]
[25] RICE S K, GAGLIARDI T A, KRASA R A. Canopy structure affects temperature distributions and free convection in moss shoot systems [J]. Am J Bot, 2018, 105(9): 1499–1511. doi: 10.1002/ajb2.1145.
[26] ZHANG J P, ZHANG J G, DUAN A G, et al. Ecological control of rocky desertification in karst areas of southwestern China [J]. Sci Silv Sin, 2008, 44(7): 84–89. [張俊佩, 張建國, 段愛國, 等. 中國西南喀斯特地區石漠化治理 [J]. 林業科學, 2008, 44(7): 84–89. doi: 10. 3321/j.issn:1001-7488.2008.07.015.]
[27] PENTECOST A, ZHANG Z H. Response of bryophytes to exposure and water availability on some European travertines [J]. J Bryol, 2006, 28(1): 21–26. doi: 10.1179/174328206X90431.
[28] KIMMERER R W, DRISCOLL M J L. Bryophyte species richness on insular boulder habitats: The effect of area, isolation, and microsite diversity [J]. Bryology, 2000, 103(4): 748–756. doi: 10.1639/0007- 2745(2000)103[0748:BSROIB]2.0.CO;2.
[29] FAN X X, DU Z X, CHAI S L, et al. Comparison on chlorophyll fluorescence kinetic response of six bryophytes to different light intensity [J]. N Hort, 2018(20): 84–90. [樊曉旭, 杜澤萱, 柴淑麗, 等. 光強脅迫下六種苔蘚植物的葉綠素熒光動力學響應差異 [J]. 北方園藝, 2018(20): 84–90. doi: 10.11937/bfyy.20173946.]
[30] ZHANG L H, QI J Q, LI T T, et al. Effects of stand density on under- story plant diversity and biomass in aplantation in Wenfeng Mountain, Xinjin County [J]. Acta Ecol Sin, 2019, 39(15): 5709–5717. [張柳樺, 齊錦秋, 李婷婷, 等. 林分密度對新津文峰山馬尾松人工林林下物種多樣性和生物量的影響[J]. 生態學報, 2019, 39(15): 5709–5717. doi: 10.5846/stxb201808151747.]
[31] LU X Q. A study of theory and technique on the vegetation restoration and reconstruction of degraded ecosystem in karst mountain area [D]. Nanjing: Nanjing Forestry University, 2006. [盧曉強. 貴州喀斯特退化森林生態系統恢復與重建理論及技術研究 [D]. 南京: 南京林業大學, 2006.]
[32] PAKEMAN R J, BROOKER R W, O’BRIEN D, et al. Using species records and ecological attributes of bryophytes to develop an ecosystem health indicator [J]. Ecol Indic, 2019, 104: 127–136. doi: 10.1016/j. ecolind.2019.04.084.
[33] TURETSKY M R, BOND-LAMBERTY B, EUSKIRCHEN E, et al. The resilience and functional role of moss in boreal and arctic eco- systems [J]. New Phytol, 2012, 196(1): 49–67. doi: 10.1111/j.1469- 8137.2012.04254.x.
[34] ZHOU H Y, ZHANG J, ZHANG D J, et al. Community characteristics of soil fauna for different canopy density of aplantation [J]. Acta Ecol Sin, 2017, 37(6): 1939–1955. [周泓楊, 張健, 張丹桔, 等. 不同郁閉度控制下馬尾松()人工林土壤動物群落特征 [J]. 生態學報, 2017, 37(6): 1939–1955. doi: 10. 5846/stxb201510232148.]
[35] WAITE M, SACK L. How does moss photosynthesis relate to leaf and canopy structure? Trait relationships for 10 Hawaiian species of contrasting light habitats [J]. New Phytol, 2010, 185(1): 156–172. doi: 10.1111/j.1469-8137.2009.03061.x.
[36] HRBá?EK F, CANNONE N, K?A?KOVá M, et al. Effect of climate and moss vegetation on ground surface temperature and the active layer among different biogeographical regions in Antarctica [J]. Catema, 2020, 190: 104562. doi: 10.1016/j.catena.2020.104562.
[37] GARCíA-CARMONA M, ARCENEGUI V, GARCíA-ORENES F, et al. The role of mosses in soil stability, fertility and microbiology six years after a post-fire salvage logging management [J]. J Environ Manage, 2020, 262: 110287. doi: 10.1016/j.jenvman.2020.110287.
Effect of Restoration Methods on Species Composition of Bryophytes in Karst Habitats
WANG Peng, WEI Hong*, CHEN Mingyue, ZHANG Mei, ZENG Chenwu, ZHANG Xuechao
(Key Laboratory of Eco-environments in Three Gorges Reservoir Region (Ministry of Education), Chongqing Key Laboratory of Plant Ecology and Resources Research in Three Gorges Reservoir Region, School of Life Sciences, Southwest University, Chongqing 400715, China)
In order to explore the effects of ecological restoration methods on bryophyte, the species composition in different habitat types in Zhongliangshan Karst area of Chongqing, China under natural abandonment and conversion of farmland to forest, et al. was studied. The results showed that there were 41 species of bryophytes in 15 families and 26 genera, including 39 mosses and 2 liverworts, andwas the dominant species. Lithophytic mosses occupy a large proportion, and their life forms are mainly tiled and clustered. The species diversity and similarity index of all habitats were low on the whole. Bryophytes in bamboo forest and Masson pine forest had higher species diversity than those in abandoned weedy land, shrub land and economic forest. Redundancy analysis showed that canopy density and light intensity were the key environmental factors affecting bryophyte distribution, tree forest provided more suitable habitats for bryophyte growth. Therefore, compared with natural restoration and economic forest restoration, returning farmland to forest was a more ideal ecological restoration method for karst degraded habitats.
Karst habitat; Ecological restoration method; Bryophyte; Biodiversity
10.11926/jtsb.4636
2022-03-11
2022-06-02
國家重點研發計劃項目(2016YFC0502304)資助
This work was supported by the National Key Research and Development Program (Grant No. 2016YFC0502304).
汪鵬(1997年生),男,碩士研究生,主要研究方向為群落生態學。E-mail: 320053854@qq.com
. E-mail: weihong@swu.edu.cn
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
中老年保健(2021年12期)2021-08-24 03:30:40
保健醫苑(2021年7期)2021-08-13 08:48:02
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
學生天地(2020年36期)2020-06-09 03:12:30
小學科學(學生版)(2020年5期)2020-05-25 07:11:32
中國生殖健康(2020年6期)2020-02-01 06:28:50
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
中國生殖健康(2019年11期)2019-01-07 01:28:02
汽車觀察(2018年10期)2018-11-06 07:05:26
