具有恐懼和強Allee效應的離散食餌-捕食者模型
2023-10-26 06:18:16胡新利李航航
西安工程大學學報 2023年4期
胡新利,李航航
(西安工程大學 理學院,陜西 西安 710048)
0 引 言
在種群系統中,群聚有利于種群的增長和生存,但種群數量過大或過小將會阻礙其發展,最終會導致種群趨于滅絕,這就是Allee效應[1]。食餌-捕食者模型是基本的種群模型之一,許多學者建立了各類食餌-捕食者模型,并對其進行動力學分析。捕食者除了直接捕殺食餌之外,還會對食餌的覓食、防御和繁殖等行為產生影響,為此,WANG等首次提出具有恐懼效應的食餌-捕食者模型[2],并引起了眾多學者的廣泛關注[3-5]。
在種群動力學中,連續或離散模型是描述種群數量變化的2種常用形式。眾多研究表明[6-7]:當種群具有不重疊的世代時,由差分方程描述的離散模型比連續模型更貼近實際。ZHU[8]和CUI[9]等分別研究了具有Holling Ⅳ功能反應的連續和離散Lotka-Volterra食餌-捕食者模型的定性行為。相比而言,離散系統要比連續系統具有更豐富的動力學行為,能夠產生倍周期分支、Neimark-Sacker分支、混沌和更復雜的動力學行為。KUNDU等研究了恐懼效應在具有線性功能反應的離散食餌-捕食者模型中的影響,并對該模型進行了穩定性分析和分支分析[10]。BANERJEE等研究了具有恐懼效應和Holling Ⅲ型功能反應的離散時滯雙食餌-捕食者模型,得到了平衡點存在的條件,并給出了正平衡點的持久性和全局穩定性的充分必要條件[11]。文獻[12]研究了具有恐懼效應的離散捕食者-食餌模型,并對其進行了穩定性分析,獲得了在正平衡點處發生倍周期分支的條件。CELIK[13]等和DIN[14]分別使用不同的離散方式研究了具有Allee效應的食餌-捕食者模型的分支分析。
1 模型的建立
有研究表明,除了直接捕殺之外,對捕食者產生的恐懼會使食餌的增長率降低40%。因此,文獻[15]提出了一類具有恐懼效應和強Allee效應的連續食餌-捕食者模型,考慮了在繁殖過程中,食餌已經受到交配誘導的強Allee效應的影響,并考慮了由于對捕食者的恐懼而使食餌的生長速度降低,具體模型如下:
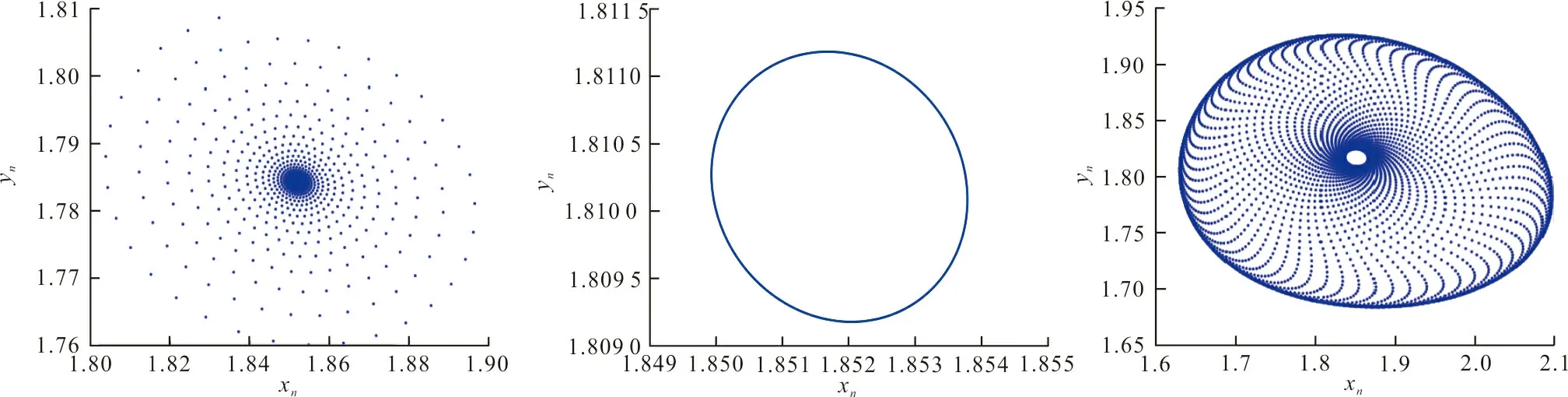
式中:r、A、f、K、b、β和d均為正常數;r為食餌種群的內稟增長率;A為食餌種群滅絕的Allee閾值(0 文獻[15]主要討論了系統(1)所有平衡點的穩定性條件,并分析了系統(1)在平衡點處發生Hopf分支。但對于離散系統會產生更豐富的動力學行為,為此,對系統(1)進行離散化。為了避免離散系統存在負解,利用文獻[16]中的分段常數變元的方法,得到連續系統(1)對應的離散系統 式中:xn和yn分別是食餌和捕食者在n世代的種群密度,其他參數同系統(1)。類似的離散化方式參見文獻[17-18]。 系統(2)的平衡點可以通過代數方程 求得。通過求解方程(3),可以得到如下定理。 定理1 系統(2)有以下4個平衡點。 1) 食餌和捕食者均滅絕的平衡點E0(0,0)。 2) 食餌存在捕食者滅絕的平衡點E1(K,0),E2(A,0)。 a21=bβyexp(bβx-d),a22=exp(bβx-d)。 定義1[19]平衡點E(x,y)處的雅可比矩陣J(x,y)對應的特征方程為 λ2-trJ(x,y)λ+detJ(x,y)=0 (4) 令λ1和λ2是上述方程(4)的2個根,有: 1) 若|λ1|<1且|λ2|<1,則E(x,y)是匯且局部漸近穩定; 2) 若|λ1|<1且|λ2|>1(或|λ1|>1且|λ2|<1),則E(x,y)是鞍點且不穩定; 3) 若|λ1|>1且|λ2|>1,則E(x,y)是源且不穩定; 4) 若|λ1|=1或|λ2|=1,則E(x,y)是非雙曲的。 通過簡單計算,系統(2)在平衡點E0(0,0)、E1(K,0)和E2(A,0)處的雅可比矩陣對應的特征方程的根分別為: 1)λ1=exp(-rA),λ2=exp(-d); 因此,可以得到如下定理。 定理2 1) 系統(2)的平衡點E0(0,0)是匯且局部漸近穩定。 2) 系統(2)的平衡點E1(K,0)具有以下性質: 3) 系統(2)的平衡點E2(A,0)具有以下性質: 為了考慮系統(2)唯一正平衡點E3(x*,y*)的穩定性,需要引進以下引理。 引理1[20]考慮二次特征多項式 F(λ)=λ2-α1λ+α2 式中:α1和α2為實系數。因此,特征多項式對應的特征方程的2個特征根都位于開的單位圓盤內的充分必要條件為|α1|<1+α2<2。 P(λ)=λ2-trJλ+detJ (5) 其中, 應用引理1得到如下定理。 當特征多項式(5)對應特征方程的特征值是一對模為1的復共軛根,則系統(2)在唯一正平衡點E3(x*,y*)處會產生一個Neimark-Sacker分支,且此條件可以寫成集合形式,即 ΩNS={(A,f,b,β,d,r,K):(trJ)2- 4detJ<0,detJ=1}。 現在討論系統(2)的所有參數在集合ΩNS的小鄰域內變化時,在唯一正平衡點E3(x*,y*)處發生的Neimark-Sacker分支。 其中|K*|?1表示小的擾動參數。令x=X-x*,y=Y-y*,則映射方程(7)轉化為 其中: g1(x,y)=a13x2+a14xy+a15y2+b1x3+ b2x2y+b3xy2+b4y3+O((|x|+|y|)4) g2(x,y)=a23x2+a24xy+d1x3+ d2x2y+O((|x|+|y|)4)。 λ2-T(K*)λ+D(K*)=0 (9) 其中: 因此,可以得到: 假設T(0)≠0,1,即 并且考慮下面的變換 在式(10)的轉換下,線性化系統(8)的標準形式可以寫為 其中: O((|u|+|v|)4)。 式中:x=a11u;y=(φ-a11)u-ψv。定義非零實數 其中: 例1 設系統(2)中的參數值為r=0.78,A=0.15,f=0.7,b=0.12,β=0.46,d=0.1,K=3.2且初始值x0=1.81,y0=1.96。 系統(2)在唯一正平衡點E3(x*,y*)處雅可比矩陣對應的特征多項式為P(λ)=λ2-1.949 3λ+0.986 6,則|1.949 3|<1+0.986 6<2。 因此,系統(2)在唯一正平衡點E3(1.811 6,1.969 8)是局部漸近穩定的(定理3),如圖1所示。 (a) 系統(2)的xn圖 (b) 系統(2)的yn圖 (c) 系統(2)的相圖圖1 系統(2)唯一正平衡點E3的局部漸近穩定性Fig.1 The local asymptotic stability diagram of the unique positive equilibrium point E3 of system (2) 例2 設系統(2)中的參數值為r=0.75,A=0.11,f=0.94,b=0.12,β=0.45,d=0.1,3 λ2-1.964 596 587 250 718λ+1=0 (12) 此外,特征方程(12)的根為: λ1=0.982 3+0.187 3i, λ2=0.982 3-0.187 3i。 且|λ1,2|=1。因此, (r,A,f,b,β,d,K)= (0.75,0.11,0.94,0.12,0.45,0.1,3.362 17)∈ΩNS。 系統(2)發生的分支圖和最大Lyapunov指數圖如圖2所示。 (a) xn的分支圖 (b) yn的分支圖 (c) 最大Lyapunov指數圖圖2 系統(2)分支圖和最大Lyapunov指數Fig.2 Bifurcation diagram and maximum Lyapunov exponents of system(2) 從圖2可以看出:當K 分支參數在K∈[3,4.2]內變化時的相圖見圖3。當K=3.3時,平衡點E3(1.851 9,1.784 4)是局部漸近穩定的,見圖3(a)。當K=3.3 時,形成光滑的閉合不變曲線,見圖3(b)。圖3(c)、(d)表明這些閉合不變曲線的半徑隨著K的增大而增大。此外,當K=4 時,閉合不變曲線消失,見圖3(e),產生混沌吸引子,見圖3(f)。 (a) K=3.3 (b) K=3.362 17 (c) K=3.38 對具有恐懼效應和強Allee效應的離散食餌-捕食者模型進行動力學分析。相比連續系統能夠發生Hopf分支之外,離散系統具有更豐富的動力學性質。以K作為分支參數,系統能夠在正平衡點處發生Neimark-Sacker分支和更復雜的動力學行為,使得捕食者和食餌種群的穩定性發生改變,最終可能導致種群之間過度擁擠或者完全崩潰。從數值模擬可以直觀地看出系統正平衡點的局部漸近穩定性、Neimark-Sacker分支等豐富的動力學行為。2 平衡點的存在性和穩定性

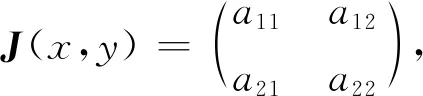
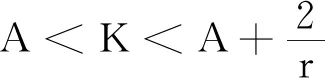

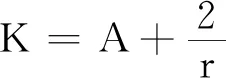
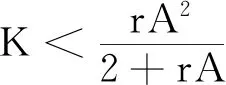
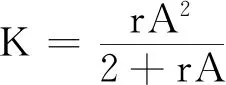


3 Neimark-Sacker分支

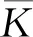
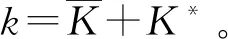



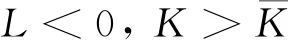
4 數值模擬


5 結 語
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38工業設計(2022年8期)2022-09-09 07:43:20核科學與工程(2021年4期)2022-01-12 06:30:26軍民兩用技術與產品(2021年10期)2021-03-16 06:05:30北京測繪(2020年12期)2020-12-29 01:33:58今日農業(2020年19期)2020-12-14 14:16:52中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50數學物理學報(2020年2期)2020-06-02 11:29:24家庭影院技術(2017年9期)2017-09-26 03:41:45中學物理·高中(2016年12期)2017-04-22 11:53:03
