超表達CsCslE1_4基因對煙草細胞壁成分的影響
2023-11-15 10:31:26姚新轉李倩倩呂立堂
種子 2023年8期
關鍵詞:煙草
姚新轉, 李倩倩, 呂立堂,2
(1.貴州大學茶學院, 貴陽 550025;2.貴州大學生命科學學院/農業生物工程研究院/山地植物資源保護與種質創新省部共建教育部重點實驗室, 貴陽 550025)
細胞壁是使植物細胞輪廓具有一定大小和硬度、同時具有一定彈性的組織。細胞壁為植物提供水分以及養料、構建植物形態,與細胞分化和植物生長發育密切相關[1]。植物細胞壁在協調抵御植物與外界環境中的生物脅迫和非生物脅迫方面起重要作用[2-3]。研究發現,在許多植物的脆性突變體中發現植物的纖維素含量下降,細胞壁結構發生改變,更容易引起植株斷裂。如在不規則木質部突變體中,細胞壁組織成分的空間構造隨著細胞次生壁中的纖維素含量下降發生改變,導致成熟植株莖稈的機械強度下降[4]。在水稻中發現的bcl-2基因的脆性突變體,出現了莖、葉的脆性,在進一步研究中發現纖維素含量降低[5]。Samuga等[6]對楊樹PtrCslD2基因進行研究,發現PtrCslD2與木質部中纖維素合成有關。Nawaz等[7]研究發現,Csl還參與抵抗非生物應激反應,如冷、熱和鹽脅迫。類纖維素合酶基因的功能以及之間的相互作用關系尚未清楚。因此,研究類纖維素合成酶E(CsIE)基因在纖維發育過程中的作用機制,有利于在分子水平上改良纖維品質,增加植物細胞壁的強度,并影響細胞和植物生長的方向[8]。本實驗通過對CsCslE1_4基因進行功能分析驗證,分析其對細胞壁成分中纖維素、半纖維素和木質素含量的影響,為選育和栽培調控提供理論依據。
1 材料與方法
1.1 試驗材料
供試煙草(NicotianatabacumL.)品種“Xanthin”由貴州大學茶學院實驗室提供,煙草種植條件為溫度(24±1)℃,相對濕度60%,光照16 h,黑暗8 h。RAN提取試劑盒、反轉錄試劑和熒光定量試劑盒購買于北京華越洋生物科技有限公司。
1.2 CsCslE1_4基因過表達載體構建
使用XbaⅠ和BamHⅠ雙酶切質粒載體pSH737,用XbaⅠ和KpnⅠ雙酶切合成的目的片段,以T4DNA連接酶連接,構成植物表達載體,β-葡萄糖苷酸酶基因(β-glucuronidase,GUS) 融合基因作為報告基因。
1.3 茶樹CsCslE1_4基因的遺傳轉化
利用農桿菌介導的葉盤法進行煙草遺傳轉化[9]。在培養皿中抗性篩選后,通過PCR以及GUS組織染色鑒定得到陽性轉基因植株,用于后續試驗。
1.4 CTAB法提取煙草總RNA以及cDNA制備
以煙草葉片為材料,采用CTAB法提取總RNA。取0.1 g樣品,研磨成細粉迅速轉移至RNA提取緩沖液中。水浴10 min,冷卻至室溫,離心后上清液轉移至新離心管中,加入等體積的水飽和酚和氯仿(比例25∶24)和10 mL NaCl,離心,加入等體積的異丙醇和2倍體積的NaCl,-80 ℃過夜沉淀。離心去上清,乙醇清洗沉淀,加適量RNA Free ddH2O溶解,即得總RNA。根據試劑盒說明書操作將RNA反轉錄為cDNA用于下一步熒光定量PCR。
1.5 轉CslE1_4基因煙草表達量檢測
利用CsCslE1_4-F:ATGTCACTGTCCCATCTCTTTG,CsCslE1_4-R:CGCCTAAGCTGTATCCGTATTT,以煙草β-actin基因作參照,設計引物Nt-actin-F:TGAGATGCACCACGAAGCTC,Nt-actin-R:CCAACATTGTCACCAGGAAGTG。根據試劑盒上的使用說明進行熒光定量分析。每個樣本設置3個重復。采用2-△△Ct法計算基因的相對表達量。
1.6 煙草細胞壁成分測定
1.6.1細胞壁多糖成分的提取
參照文獻[10]的測定方法,稱取0.05 g(干重)樣品使用磷酸緩沖液研磨至勻漿,反復用磷酸緩沖液洗滌研缽和研棒,離心。沉淀中依次加入氯仿-甲醇(體積比1∶1)去除脂類、DMSO-H2O(體積比9∶1)提取淀粉,蒸餾水清洗,殘渣即為粗細胞壁,備用。果膠以半乳糖醛酸含量制作標準曲線,比色法測定糖醛酸吸光值并計算相應的果膠含量;半纖維素含量以木糖為標準糖制作標準曲線,比色法測定戊糖,并計算相應半纖維素含量;纖維素含量的測定以葡萄糖含量制作標準曲線,采用蒽酮-硫酸法計算相應的纖維素含量,比色法測定己糖。
1.6.2木質素含量測定
參照劉佳佳[11]的乙酰溴法,以280 nm處的吸光值表示每克鮮樣中木質素的相對含量。每個樣品3個生物學重復。
1.7 數據處理
實驗數據使用Excel和GraphPad Prism5.0軟件進行分析處理。
2 結果與分析
2.1 CslE1_4基因的克隆與轉基因株系的獲得
以茶樹葉片cDNA為模板,以特異性引物擴增得到CslE1_4序列,克隆得到的CslE1_4序列與過表達載體pSH737連接,構成植物表達載體pSH737-CsCslE1_4(圖1)。
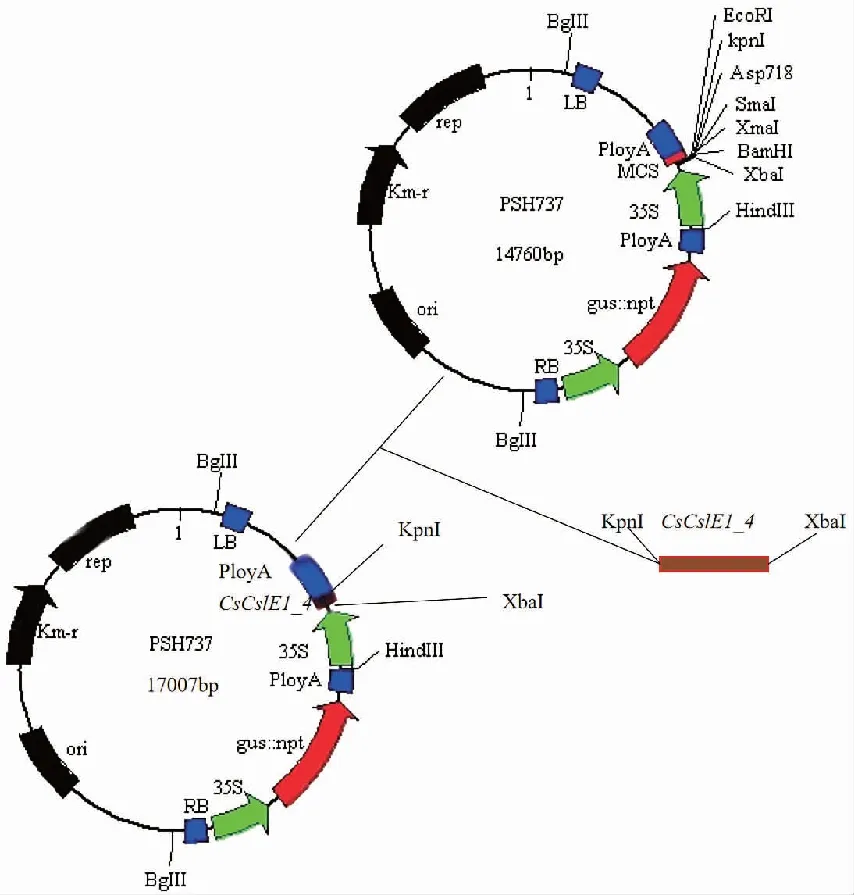
圖1 CsCslE1_4基因過表達載體構建圖譜
通過農桿菌介導法,將含有CsCslE1_4重組質粒的農桿菌菌液對煙草葉片遺傳轉化,獲得18株轉基因煙草(圖2 A~圖2 D)。對煙草進行GUS組織化學染色以及PCR鑒定,結果(圖2 E、F)發現,在GUS染液中,野生型煙草植株并未檢測到GUS活性,在抗性煙草植株中檢測到GUS活性;經PCR鑒定,陽性轉基因植株有一條大小約120 bp的條帶(圖2 F),陰性對照中并未擴增出目的條帶,說明CslE1_4基因已經成功整合到煙草中。
2.2 轉CslE1_4基因煙草株系的表達量分析
通過RT-qPCR分析轉基因株系中CslE1_4基因的表達量(圖3)。結果發現,18株轉基因煙草株系中CslE1、CslE6、CslE13、CslE17的表達量分別是野生型的19.06倍、13.50倍、6.73倍和5.18倍,差異極顯著(圖3),其他株系中CslE16表達量最低。
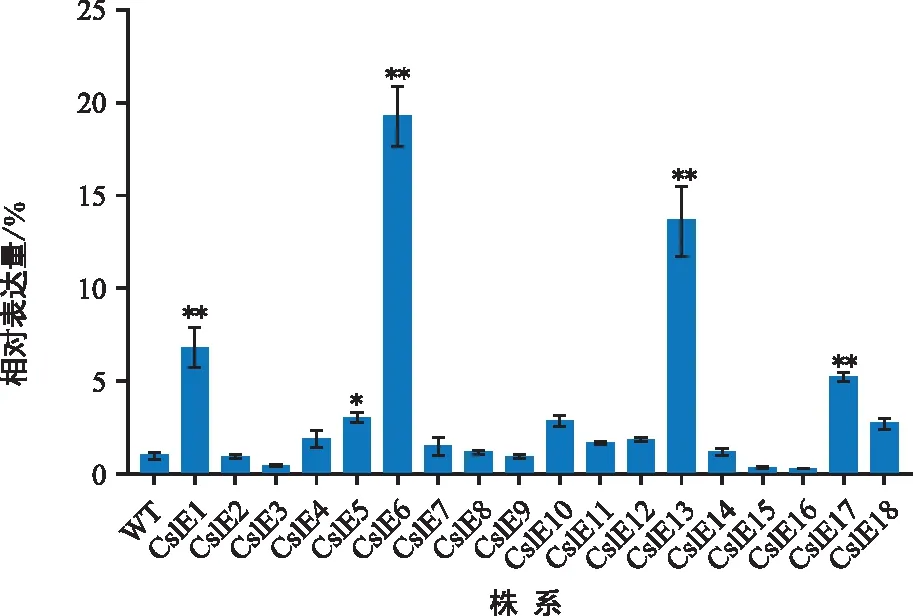
注:WT為野生型煙草;CslE為轉基因煙草,誤差線表示3個重復的標準偏差;“*”表示差異顯著(p<0.05);“**”表示差異極顯著(p<0.01)。下同。
2.3 纖維素含量
對轉CslE1_4基因煙草的葉片與莖段的纖維素含量進行測定,結果(圖4)顯示,除株系CslE4外,轉基因株系莖中纖維素含量都顯著高于同株系葉中的纖維素含量,其中差異最大的是株系CslE2,莖段纖維素含量是葉片纖維素含量的7.17倍。同時,與野生型相比,轉基因株系莖的纖維素含量是野生型的1.43~5.48倍,葉中纖維素含量除株系CslE2外,其余株系纖維素含量是野生型的1.08~2.49倍。由此可見,大多數轉基因株系,其纖維素含量相對較高,部分可能是由于CslE1_4基因表達低。
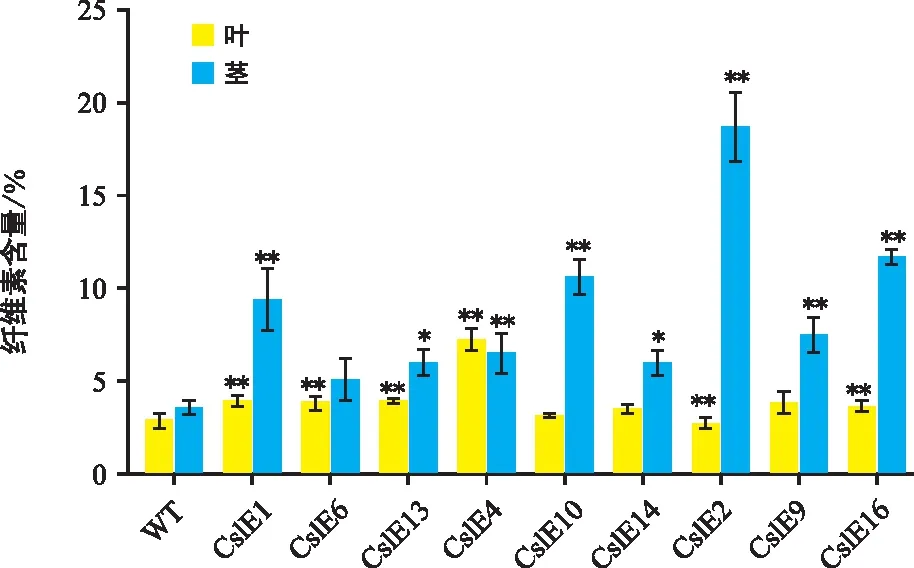
圖4 轉基因煙草的纖維素含量
2.4 半纖維素含量
轉CslE1_4基因煙草的葉片和莖段的半纖維素含量測定結果(圖5)顯示,每個株系的半纖維素含量變化與纖維素含量趨勢基本一致。除株系CslE4外,野生型和轉基因株系莖中的半纖維素含量均高于同株系的葉片。與野生型相比,轉基因株系葉中纖維素含量除株系CslE2和CslE10外,其余株系半纖維素含量是野生型半纖維素含量的1.18~2.22倍,其中含量最高的株系為CslE4;莖中半纖維素含量是野生型半纖維素含量的1.47~5.74倍,含量最低的株系是CslE6,最高的株系是CslE2。
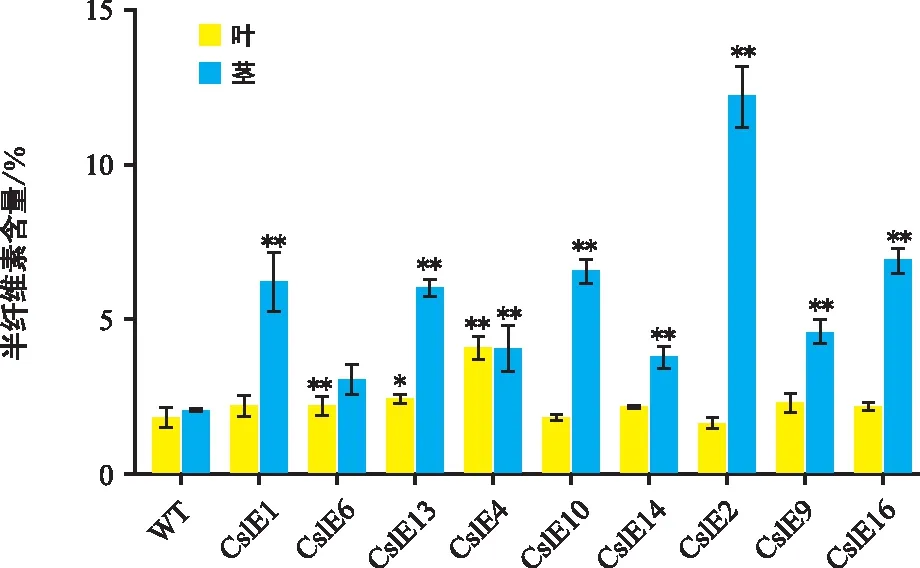
圖5 轉基因煙草半纖維素含量
2.5 果膠含量
野生型和轉基因煙草的果膠含量測定結果(圖6)表明,莖段中的果膠含量比葉片低,轉基因煙草果膠含量在葉與莖中均比野生型煙草果膠含量高。轉基因株系葉中果膠含量是野生型的1.08~1.84倍,其中含量最低的株系是CslE9,最高的是CslE4。莖中轉基因株系果膠含量是野生型的1.29~3.67倍,含量最低的株系是CslE2,最高的是CslE14。同時發現,同一株系的纖維素和半纖維素含量增高時,果膠含量存在補償性的降低,即果膠含量變化趨勢與纖維素和半纖維素含量變化趨勢相反。
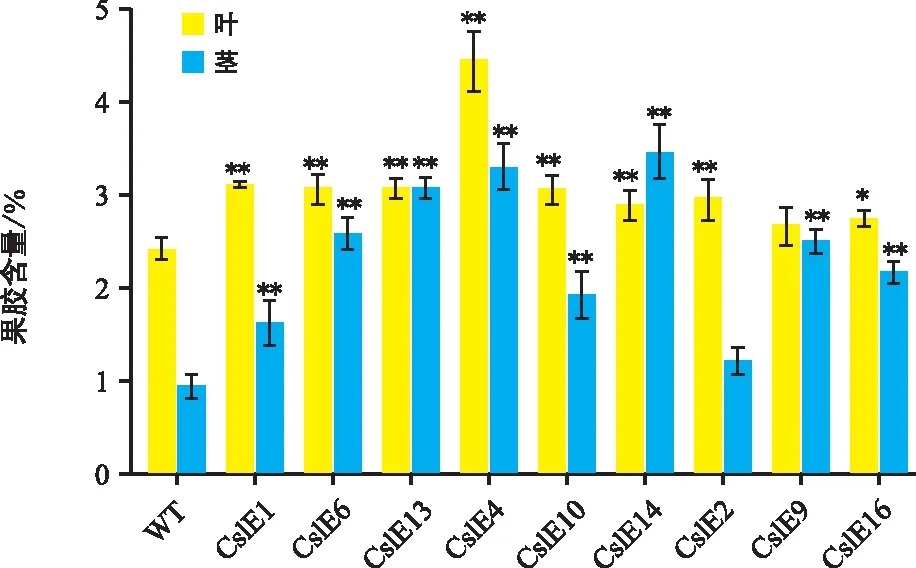
圖6 轉基因煙草果膠含量
2.6 木質素含量
采用乙酰溴測定木質素含量法,測定過表達轉CslE1_4基因煙草葉片中的木質素(圖7)。以野生型煙草為對照,在葉片中CslE1的木質素含量最高,是野生型煙草的1.09倍。莖段中CslE10的木質素含量最高,是野生型的1.05倍。其余轉基因煙草木質素含量在葉片中均有一定程度的提高,而在莖段中呈降低的趨勢。結果表明,轉CslE1_4基因對煙草的木質素含量沒有太大影響。

圖7 轉基因煙草的木質素含量
3 討 論
細胞壁是植物適應和生存的關鍵,能夠調節植物的生長發育,其主要成分包括纖維素和半纖維素[12]。本研究通過對野生型和轉基因煙草進行基因表達以及細胞壁成分測定,發現與野生型煙草相比,過量表達后的轉基因煙草的細胞壁多糖成分含量均有提高,如纖維素、半纖維素和果膠。轉基因煙草的莖段比葉片的纖維素和半纖維素提高更加明顯,果膠含量較葉片降低,木質素含量沒有明顯的變化。
纖維素是植物細胞壁重要的組成成分之一,在本研究中,與野生型相比,轉基因株系纖維素含量高于野生型煙草。在油菜(Brassicacampestris)[13]、大豆(Glycinemax)[14-15]研究中也有類似結果;高纖維素含量可提高植株的機械強度,增強抗倒伏能力[16-17]。一般認為,半纖維素與纖維素形成的交聯結構可提高細胞壁強度,從而提高植株的抗倒伏能力[18-19]。本研究發現,野生型和轉基因株系莖和葉片中的半纖維素含量均高于野生型煙草,其中莖半纖維素含量高于同株系葉片半纖維素含量。參與木聚糖合成的IRX14H基因[20]與維持木聚糖含量和穩定細胞壁的IRX15-L基因均在直立性好的品種中表達量較高[21]。因此,結合基因表達量以及轉基因煙草的半纖維素含量結果分析,推測在煙草莖發育中,半纖維素合成相關基因表達量增加,導致半纖維素含量增加。果膠作為初生細胞壁的主要組成物質,對植物細胞的形態建成具有重要作用[22]。隨著纖維的發育,各果膠前體核苷糖含量均有下降趨勢。纖維品質極好的陸地棉品系的果膠含量明顯低于品質較差的品系[23]。在細胞壁多糖成分測定結果中,轉基因煙草的纖維素、半纖維素含量的變化趨勢基本一致,果膠含量則相反。并且,轉基因株系在葉片中的纖維素和半纖維素含量均降低,果膠含量補償性增加;而莖段纖維素含量和半纖維素含量均升高,果膠含量反而降低。本研究認為,低含量的果膠可以增加細胞壁的硬度,從而提高煙草的硬度。
4 結 論
CslE1_4基因可能與植物中纖維素合成有關,調控植物中纖維素的形成,影響植物中細胞壁組成成分含量。本研究結果可為煙草及其他作物品種在選育和栽培上提供理論依據。
猜你喜歡
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
中國煙草學報(2019年5期)2019-11-14 07:54:12
首都公共衛生(2019年5期)2019-05-21 01:08:34
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
新聞傳播(2016年3期)2016-07-12 12:55:34
當代化工研究(2016年9期)2016-03-20 16:22:08
自動化博覽(2014年6期)2014-02-28 22:32:15
聲屏世界(2014年6期)2014-02-28 15:18:09
西南學林(2013年2期)2013-11-12 12:58:54
中國煙草學報(2012年5期)2012-04-12 06:21:18
