葡萄糖轉運蛋白4 在魚類糖代謝中的作用研究進展
2023-11-16 06:31:16吳杉杉鄧瑤霍歡歡彭墨
水產學雜志 2023年5期
吳杉杉,鄧瑤,霍歡歡,彭墨
(江西農業大學動物科學技術學院,江西 南昌 330045)
硬骨魚比哺乳動物不耐受葡萄糖,尤其是肉食性魚類[1]。在哺乳動物中,葡萄糖從血液運輸到細胞內由葡萄糖轉運蛋白家族(GLUTs)完成[2],目前已報道了14 種不同的葡萄糖轉運蛋白,即GLUT1~12、HMIT 和GLUT14[3]。GLUT1~4 主要介導葡萄糖的轉運,其他GLUTs 主要介導果糖轉運等功能[3]。GLUT1、GLUT2 和GLUT3 分別廣泛分布于紅細胞、胰腺[4],肝、胰[5]和神經組織中[3],GLUT4 則在骨骼肌和脂肪組織中高度表達[6],介導組織細胞攝取葡萄糖。
GLUT4 協助葡萄糖通過細胞膜脂質雙層進入細胞,形成組織葡萄糖動態供給平衡,維持機體正常的糖代謝[7]。自發現GLUT4 以來,深入研究了其結構、儲存部位、功能以及參與葡萄糖轉運的相關信號途徑等[8-11],在哺乳動物中發現GLUT4 參與了PI3K/Akt、AMPK、PKC 等信號通路調節葡萄糖轉運[12-14]。
目前在魚類中多研究GLUT4 的基因序列分析、組織表達。Li 等[15]分析了卵形鯧鲹(Trachinotus ovatus)、軍曹魚(Rachycenton canadum)和尼羅羅非魚(Oreochromis niloticus)GLUT4 的表征;劉含亮[16]研究了羅非魚GLUT4 的組織表達,但缺乏長期以來涉及葡萄糖轉運機制的研究。本文綜述了GLUT4在魚類中的發現歷程、結構、功能和轉運方式以及相關信號通路的最新研究成果,為進一步了解魚類中GLUT4 的作用和機制提供理論基礎。
1 魚類中GLUT4 的發現歷程
最初,廣泛研究了魚類不耐受葡萄糖的機理。Wright 等[17]研究指出,羅非魚外周組織中缺乏GLUT4,是一種自然發生的GLUT4 基因缺失。Legate 等[18]在虹鱒(Oncorhynchus mykiss)、美洲鰻鱺(Anguilla rostrata)、黑(Ameiurus melas)的白肌和紅肌以及心臟中都沒有檢測到哺乳動物類型的GLUT4,因此,猜測硬骨魚體內缺乏GLUT4 是魚類不耐受葡萄糖的一大原因。隨后Josep 實驗室[19]從褐鱒(Salmo trutta)肌肉中克隆了一種葡萄糖轉運蛋白(btGLUT)。它與哺乳動物的GLUT4 具有很高的序列同源性,主要在紅肌和白肌中表達,這是褐鱒攝取葡萄糖的兩個主要部位,在鰓、腎臟和脂肪組織中也有較高表達。進一步研究發現,btGLUT 在紅肌中的表達與胰島素水平直接相關,相比之下,白肌中btGLUT 的表達似乎不受胰島素水平變化的影響[20]。隨后的研究結果顯示,btGLUT4 蛋白水平的變化與轉錄水平呈一致的相關性,表明褐鱒魚體內btGLUT4 的調節主要發生在紅肌中[21]。該實驗室后期的研究確定了褐鱒魚紅肌中的轉運蛋白為GLUT4,即在褐鱒原代骨骼肌細胞中,胰島素促進內源性GLUT4 移位到質膜,促進葡萄糖轉運[22]。Díaz 等[23]對虹鱒的研究表明,虹鱒骨骼肌中的GLUT4 在轉錄和蛋白水平上受胰島素的調節,同時胰島素和胰島素樣生長因子1(insulin-like growth factors-1,IGF-1)可能直接作用于虹鱒的肌肉細胞,調節GLUT4 的表達。因此,魚類GLUT4 和哺乳動物的GLUT4 功能一致,是葡萄糖進入骨骼肌的重要介質,對膳食葡萄糖的利用非常重要。
在魚類骨骼肌中發現GLUT4 的同時,其他組織也有GLUT4 的分布。Capilla 等[24]從銀大麻哈魚(Oncorhynchus kisutch)的脂肪組織克隆出okGLUT4,發現其與哺乳動物GLUT4 具有高度的序列同源性。盡管這兩者之間的蛋白質基序存在差異,當okGLUT4 在銀大麻哈魚脂肪細胞中表達時,卻能響應胰島素的刺激從細胞內轉運至質膜,這表明okGLUT4 是哺乳動物GLUT4 的結構性和功能性同源物,但對葡萄糖的親和力較低。Capilla 等[24]的研究在一定程度上可以解釋魚類對葡萄糖不耐受的原因。陳強[25]和劉含亮[16]的研究發現,羅非魚、卵形鯧鲹和軍曹魚的GLUT4 與其他物種相比較具有很高的同源性。羅非魚骨骼肌中克隆出了GLUT4,在肌肉和心臟中的表達量很高[16];將卵形鯧鲹和軍曹魚的GLUT4 進行多序列比較分析發現,FQQI、TELEY 等基因序列的突變可能導致GLUT4 功能、轉運方式的改變,影響魚類對葡萄糖的吸收和利用[25]。這與先前Wright 等[17]提出羅非魚體內缺失GLUT4的結論相沖突。除此之外,大西洋鱈(Gadus morhua)、龍膽石斑魚(Epinephelus lanceolatus)的GLUT4 相應在心臟和肌肉[26]、肌肉和眼睛[27]中有較高的表達。綜上研究,魚類的GLUT4 與哺乳動物相比存在較高的保守性,推測魚類GLUT4 具備轉運葡萄糖的功能,對研究魚類攝取葡萄糖的機制提供理論依據。
2 GLUT4 的結構、功能與轉運模式
GLUT4 由12 個跨膜結構域蛋白組成[8],在基礎條件下,當GLUT4 從質膜移動到早期內吞體(early endosome,EE),在返回質膜之前從EE 移動到循環內吞體(recycling endosome,RE)時,GLUT4通過內吞和胞吐進行結構性循環。還對RE 和反面高爾基網(trans-golgi network,TGN)中的GLUT4 進行了動態分選,以生成GLUT4 儲存囊泡(GSVs)[12]。正常胰島素敏感狀態下通過將胰島素響應的GSVs易位到細胞表面來促進葡萄糖攝取,這種囊泡群體既不同于RE,也不同于TGN,它的形成可能支持GLUT4 的獨特特性,這些特性只有在胰島素敏感細胞中表達時才明顯[9,28]。
胰島素刺激的GSVs 轉位較為復雜,可溶性NFS 附著蛋白受體(soluble N-ethylmaleimide-sensitive factor attachment protein receptor,SNARE)是介導GSVs 與細胞膜融合的核心分子,Sec1p/Munc 18類蛋白(Sec1/Munc18-like proteins,SM)和SNARE協同作用調控膜融合過程,存在靶膜上的SNAP 受體(target-SNARE,t-SNARE)以及囊泡膜上的SNAP受體(vesicle-SNARE,v-SNARE)[29],t-SNARE 包含突觸融合蛋白4(syntaxin-4,Stx4)和23 kDa 突觸關聯蛋白(synaptosomal associated protein 23 kDa,SNAP23)[12],囊泡相關膜蛋白2(vesicle associated membrane protein 2,VAMP2)是胰島素敏感組織中主要的v-SNARE[30]。基礎狀態下Munc18c 與Stx4的結合使其處于“關閉”狀態,從而抑制GSVs 與細胞膜的融合過程,經胰島素刺激Munc18c 同Stx4 解離,Stx4 轉變為“開放”狀態[31],通過雙C2 域蛋白-β(double C2-like domains beta,DOC2B)和擴展突觸結合蛋白1(extended synaptotagmin-like protein 1,ESYT1)等一些Ca2+敏感蛋白調節Stx4、SNAP23 和VAMP2 形成三元復合物[12],Stx4 和SNAP23 足以將囊泡拴在細胞膜上,而VAMP2 促進兩者的融合[32],釋放GSVs 中的GLUT4,轉運葡萄糖。
在不同魚類中GLUT4 蛋白相對保守,不同魚種類間GLUT4 的外顯子數目和基因長度不同:河鲀(Tetraodontidae)、綠河鲀(Tetraodon nigroviridis)、羅非魚、刺魚(Gasterosteus)、青鳉(Oryzias latipes)和花斑劍尾魚(Xiphophorus maculatus)分別有12、13、13、11、12 和15 個外顯子,基因長度分別為4.5 kb、4.5kb、10.2kb、4.9kb、12.7kb 和14.1kb[33]。人類GLUT4基因外顯子VI~X 在魚類中高度保守(圖1)。

圖1 魚類GLUT4 的基因組結構[33]Fig.1 Genomic structure of GLUT4 in fish
目前,已通過外源性(哺乳動物L6 肌肉細胞或3T3-L1 脂肪細胞)和內源性(鱒原代肌肉細胞)成功研究了鮭體內兩種葡萄糖轉運蛋白(btGLUT4 和okGLUT4)的細胞內轉運特性,并與哺乳動物的GLUT4 進行了比較。當btGLUT4 在L6 肌肉細胞中穩定表達時,btGLUT4 在PM 中的表達水平(20%~25%)高于大鼠GLUT4(10%~15%)[22]。這不僅在肌肉細胞中觀察到,因為在穩態條件下分別瞬時表達btGLUT4 或okGLUT4 的3T3-L1 脂肪細胞中,btGLUT4 在PM的蛋白水平(30%~40%)顯著高于大鼠GLUT4(10%~15%),也顯著高于okGLUT4(15%~20%)[24,34]。重要的是,鱒原代肌肉細胞中GLUT4在質膜上的基礎定位也相對較高[22]。因此,在基礎狀態下,魚類和哺乳動物在細胞內保留GLUT4 的機制不同。如前所述,哺乳動物細胞中的GLUT4 主要分布于GSVs,而研究表明,btGLUT4 在兩種細胞類型中均可以通過胰島素刺激轉位至PM。除此之外,當btGLUT4 和大鼠GLUT4 在3T3-L1 脂肪細胞[34]或L6 肌肉細胞[22]中共表達時,btGLUT4 僅顯示出部分共同定位。表明btGLUT4 可能分布在獨特的儲存室中,在儲存室與PM之間進行結構性循環。根據魚類與哺乳動物GLUT4 在基礎狀態下PM 定位的差異,okGLUT4 比btGLUT4 更類似于哺乳動物對應蛋白的轉運方式。有人認為這兩種GLUT4 的不同轉運方式可能與GLUT4 蛋白序列(即N-端和C-端蛋白質基序)的不同調控特征有關[34]。因此,證明了魚類的GLUT4 與哺乳動物同源物一樣具有胰島素反應性,即使它們在穩態時具有更高的PM定位。
3 與GLUT4 有關的信號通路
GLUT4 介導的葡萄糖轉運涉及PI3K/Akt、AMPK、PKC 等信號通路,目前有關哺乳動物的各種信號通路調控GLUT4 轉運葡萄糖的機制已有廣泛研究,而在魚類中,GLUT4 轉運葡萄糖的具體機制還不明確。
3.1 PI3K/Akt 信號通路在葡萄糖轉運中的作用
PI3K/Akt 信號通路參與胰島素刺激GLUT4 轉位[35]。PI3K 是一種二聚體,由一個催化亞基P110 和一個調節亞基P85 組成。根據序列的相似度和底物的特異性哺乳動物的PI3K 分為3 種亞型,I 型以磷脂酰肌醇(phosphatidylinositol,PI)、磷脂酰肌醇磷酸(phosphatidylinositol phosphate,PIP)、磷脂酰肌醇-4,5-二磷酸(phosphatidylinositol-4,5 -bisphosphate,PIP2)為底物;II 型以PI 及PIP 為底物,III 型以PI 為底物[36]。Akt 包括Akt1、Akt2 和Akt3 三個亞型,每個亞型都有一個PH 區域能夠被磷脂酰肌醇-3,4,5-三磷酸(phosphatidylinositol-3,4,5-trisphosphate,PIP3)識別。
胰島素刺激后,磷酸化的胰島素受體(insulin receptor,IR)與胰島素受體底物(insulin receptor substrate,IRS)結合,激活IRS,讓活化的PI3K 促使PIP2 磷酸化,產生PIP3[12],隨后3-磷酸肌醇依賴性蛋白激酶1(3-phosphoinositide-dependent protein kinase 1,PDK1)磷酸化Akt[37],導致TBC1(tre-2/USP6、BUB2、cdc16)結構域家族成員1(TBC1D1)和TBC1(tre-2/USP6、BUB2、cdc16)結構域家族成員160 kDa 蛋白激酶B 底物(TBC1D4/AS160)失活,抑制了其GTP 酶激活蛋白(GTPase-activating protein,GAP)活性,從而允許Rab 家族的GTP 酶負載GTP[38,39],最終由Rab10 的激活刺激3T3-L1 脂肪細胞中的GLUT4 轉位[40],骨骼肌細胞中的GLUT4 轉位通過Rab8A、Rab10、Rab13 和Rab14 的激活[41-43]。胰島素單獨刺激脂肪細胞后,經Cbl-CAP-APS-TC10 信號通路介導GLUT4 轉運葡萄糖(圖2)[44]。

圖2 PI3K/Akt 信號通路和胰島素刺激產生的Cbl-CAPAPS 復合物調控GLUT4 轉運葡萄糖[54]Fig.2 The PI3K/Akt pathway and the Cbl-CAP-APS complex produced by insulin stimulation enhances GLUT4 translocation,eventually resulting in an increase of glucose uptake
目前的研究發現,PI3K/Akt 基因廣泛分布在斑馬魚(Brachydanio rerio var)、黃顙魚(Pelteobagrus fulvidraco)和草魚(Ctenopharyngodon idella)等魚類組織中[45-47]。有研究指出,鋅可以通過上調矛尾復虎魚(Synechogobius hasta)肝細胞中IR、IRS 和PI3K 的mRNA 表達,激活胰島素信號通路[48]。Gu等[49]的研究顯示,胰島素介導的PI3K/Akt 信號途徑能夠影響大黃魚(Larimichthys crocea)的葡萄糖利用,其是否通過GLUT4 轉運來影響葡萄糖吸收還未知。Yang 等[50]的研究表明,注射胰島素能夠上調鯉(Cyprinus carpio L.)肌肉中GLUT4 的mRNA 表達水平。除此之外,有研究探討了通過PI3K/Akt 信號通路調控團頭魴(Megalobrama amblycephala)的抗氧化機制以及胰島素激活IRS-PI3K/Akt 信號通路調控黃顙魚和團頭魴脂肪代謝的分子機制[51-53]。
3.2 AMPK 信號通路在葡萄糖轉運中的作用
在哺乳動物中已證實,AMPK 為一種能量傳感器,能影響GLUT4 細胞內轉位和葡萄糖吸收[14]。AMPK 是一種異源三聚體復合物,由一個催化亞基(α-亞基)和兩個調節亞基(β-亞基和γ-亞基)組成,又分為兩個α 亞基異構體(α1 和α2),兩個β 亞基異構體(β1 和β2)和三個γ 亞基異構體(γ1,γ2 和γ3)。α-亞基包含N 端的絲氨酸/蘇氨酸蛋白激酶的殘基Thr172,AMPK 的激活離不開Thr172 的磷酸化[55]。AMPKα1 在腎、肝、肺、心和腦中大量表達,AMPKα2 在骨骼肌、心臟和肝臟中表達最為顯著[56]。
在哺乳動物中,除二甲雙胍、白藜蘆醇、AICAR等AMPK 激活劑可以激活AMPK 之外[57],運動也能夠激活AMPK[58]。在能量改變即AMP/ATP 比率變化的情況下,通過腫瘤抑制因子肝激酶B1(liver kinase B1,LKB1)的響應磷酸化AMPKα 的Thr172位點[59]。鈣離子/鈣調蛋白依賴性蛋白激酶激酶β(Ca2+/calmodulin-dependent protein kinase kinase β,CaMKKβ)也是AMPK 的替代上游激酶,增加細胞內Ca2+濃度后,促使CaMKKβ 磷酸化并激活AMPK[60]。研究顯示,二甲雙胍以AMPK 依賴的方式增加Src 的磷酸化,Src 抑制可阻斷二甲雙胍介導的Cbl 磷酸化。二甲雙胍可以刺激c-jun 氨基末端激酶(c-jun N-terminal kinases,JNK)及其下游分子c-jun 的磷酸化。c-jun 是Cbl 相關蛋白(Cbl-associated protein,CAP)轉錄的關鍵分子,然后通過Cbl/CAP 相關復合體的形成來調節GLUT4 的轉位[61]。除此之外,還可以通過AMPK/TBC1D1 信號通路刺激GSVs 轉運上膜(圖3)。

圖3 AMPK 信號通路調控GLUT4 轉運葡萄糖[58,60]Fig.3 The AMPK pathway enhances GLUT4 translocation,eventually resulting in an increase of glucose uptake
TBC1D1 作為TBC1D4/AS160 的類似物,TBC1D1 的GAP 結構域具有功能性,并且具有與TBC1D4/AS160 高度相似的GAP 結構域幾乎相同的Rab 特異性[41]。AMPK 是TBC1D1 磷酸化的觸發器,通過磷酸化TBC1D1 Ser231 位點抑制其Rab-GAP 活性,然后相關的Rabs 被解除抑制,促進GSVs 轉運至細胞膜并嵌入脂質雙分子層釋放GLUT4,介導非胰島素依賴的骨骼肌攝取葡萄糖[62]。
3.3 PKC 信號通路在葡萄糖轉運中的作用
PKC 是一組依賴Ca2+和磷脂激活的絲氨酸/蘇氨酸蛋白激酶。哺乳動物的傳統PKC 信號途徑參與非胰島素調控的葡萄糖轉運,同時非典型PKC 與PI3K/Akt 信號途徑相關聯。傳統PKC 信號途徑中,胞外信號分子與其相應的G 蛋白偶聯受體結合后,激活的G 蛋白介導磷脂酶C(phospholipase C,PLC)的激活,導致帶有PIP2 的脂質水解。該反應產生了兩個第二信使二酰甘油(diacylglycerol,DAG)和1,4,5-三磷酸肌醇(inositol-1,4,5-triphosphosate,IP3),后者誘導內質網釋放Ca2+。由于DAG/Ca2+的作用,將PKC 募集至內膜可緩解酶的相互抑制,激活其激酶功能[69],之后Rab13 參與Ca2+介導的肌細胞GLUT4 胞吐過程(圖4)[13]。非典型PKC 信號途徑中,對3T3L1 脂肪細胞給予外源性PIP3,隨后會增加下游信號蛋白PKCζ/λ 的磷酸化,上調GLUT4 蛋白表達量,促進葡萄糖攝取(圖2)[70]。研究表明諸多PKC 亞型如PKCβ、PKCγ、PKCε 和PKCι/λ 的表達促進GLUT4 的轉位[71]。
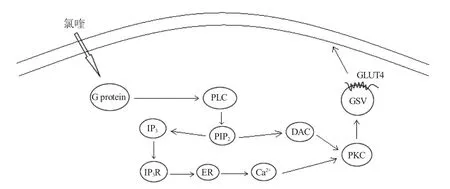
圖4 PKC 信號通路調控GLUT4 轉運葡萄糖[71]Fig.4 The PKC pathway enhances GLUT4 translocation,eventually resulting in an increase of glucose uptake
有關魚類PKC 的研究大多停留在基因序列分析、組織表達以及亞型層面上,是否通過PKC 信號通路影響魚類GLUT4 轉運及葡萄糖吸收目前尚不清楚。魚類組織中分布的PKC 亞型較為廣泛,如在虹鱒腦中發現的PKCα、PKCβ 和PKCγ[72],同時在美國紅魚(Sciaenops ocellatus)白細胞中鑒定出多種PKC 亞型[73]。傳統型的和非典型PKC 亞型存在于金魚(Carassius auratus)的垂體內[74]。PKCε 和PKCζ 廣泛分布于斑馬魚中樞神經系統,PKCθ 和PKCβII 在骨骼肌中表達[75]。PKC 信號通路在魚類中的研究涉及免疫和脂肪代謝等方面。PKCθ 在草魚宿主免疫防御機制中起重要作用[76],以及PKC 信號通路在虹鱒禁食期間被激活,介導脂質分解過程[77]。
4 小結
目前對魚類糖代謝主要專注于糖異生、糖酵解等方面,而魚類葡萄糖轉運體的機制研究十分有限。重要的是,葡萄糖轉運蛋白4(GLUT4)在魚類中的研究有較大的進展。已經證明魚類GLUT4 與哺乳動物具有較高的同源性,至少在進化上是保守的。其次,魚類GLUT4 的結構和轉運方式與哺乳動物存在一定的差異。GLUT4 的信號通路在哺乳動物中研究的廣泛且深入,魚類GLUT4 的信號通路研究還有許多空白。在胰島素依賴途徑影響魚類葡萄糖的吸收方面得到初步探究。已經證明,AMPK 作為非胰島素依賴性葡萄糖轉運途徑影響魚體吸收葡萄糖的進程,了解對葡萄糖不耐受的魚類的AMPK 刺激葡萄糖攝取的具體機制,為改善魚類糖耐受能力具有重要的意義。然而,目前PKC 信號通路未涉及到魚類GLUT4 介導的葡萄糖代謝。因此針對哺乳動物和魚類在不同信號通路上的作用差異以及不同信號通路中上游因子對魚類GLUT4 轉運的具體調控機制等的研究還需深入探討。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
科技傳播(2019年22期)2020-01-14 03:06:54
中國生殖健康(2019年2期)2019-08-23 08:12:10
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:25:42
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
中國衛生標準管理(2015年1期)2016-01-14 03:41:27
