瘤胃保護型胍基乙酸對肉牛育肥后期生產性能、血清生化指標及胴體品質的影響
2023-11-20 12:58:28許遲吳仙花趙莉岳家琪胡波趙強
當代畜禽養殖業 2023年5期
關鍵詞:血清
許遲,吳仙花,趙莉,岳家琪,胡波,趙強
(寧夏大北農科技實業有限公司,寧夏銀川 750200)
近年來, 隨著生活水平的提高和膳食結構的調整,人們對高品質牛肉產品的需求日益增加[1-2]。因此,如何在提高肉牛生長性能的同時,提升肉牛瘦肉沉積率和胴體品質,將越來越受重視。 研究發現,肌酸(Cr)作為內源物質主要儲存在肌肉組織中,可促進蛋白質合成,畜禽育肥后期可改善畜禽胴體品質,起到增肌塑形的作用[3]。 此外,Cr 促進能量代謝作用顯著,可優化家畜的體脂分配,降低料重比,提高養殖綜合經濟收益[4]。 Cr 主要存在于動物源性飼料原料中,包括魚粉、肉骨粉和血粉。 植物性飼糧成分中不含有Cr[5]。 在飼糧中直接補充Cr 或者Cr 水合物,對畜禽機體血漿中的Cr、肌酐濃度無顯著影響[6]。補充特異性營養物質可促使牛羊機體合成內源性Cr、改善其生產性能與胴體品質。 歐美地區通過營養調控有效提高了育肥牛、羊內源性Cr 含量[7-9],使育肥牛、羊的胴體品質、肉品質及生產性能得以提高。
胍基乙酸(guanidinoacetic acid,GAA)是動物體內Cr 合成的前體物質, 參與動物的肌肉合成與能量代謝,因其具備制備方法簡單、生產成本低且穩定等優點而備受青睞[10]。 GAA 通過血液循環被運送到肝臟,甲基化生成Cr[11-14]。 研究表明,直接補充氨基酸在一定程度上可以提高育肥羊的生產性能,但大部分氨基酸會在瘤胃中被瘤胃微生物降解,采用過瘤胃的方法可以降低氨基酸在瘤胃中的降解率,更大程度上發揮氨基酸的生物學功能[15]。 因此,本試驗旨在探究日糧中添加瘤胃保護型胍基乙酸 (rumen protective guanidinoacetic acid,RPG)對育肥后期西門塔爾牛生產性能、血清生化、屠宰性能及胴體品質的影響。 以期為RPG 在育肥后期肉牛養殖中的應用提供一定的理論依據。
1 材料與方法
1.1 試驗設計
選擇遺傳背景一致、平均體重為(585.13±5.65) kg及膘情相近的西門塔爾肉牛120 頭, 隨機分為2 組,每組3 個重復(欄),每個重復(欄)20 頭。 對照組飼喂基礎日糧, 試驗組飼喂添加了含0.34% RPG 濃縮飼料配制的飼糧。 各組的粗飼料原料及用量保持一致。日糧組成和營養水平見表1。 預試期為10 d,正試期為90 d。
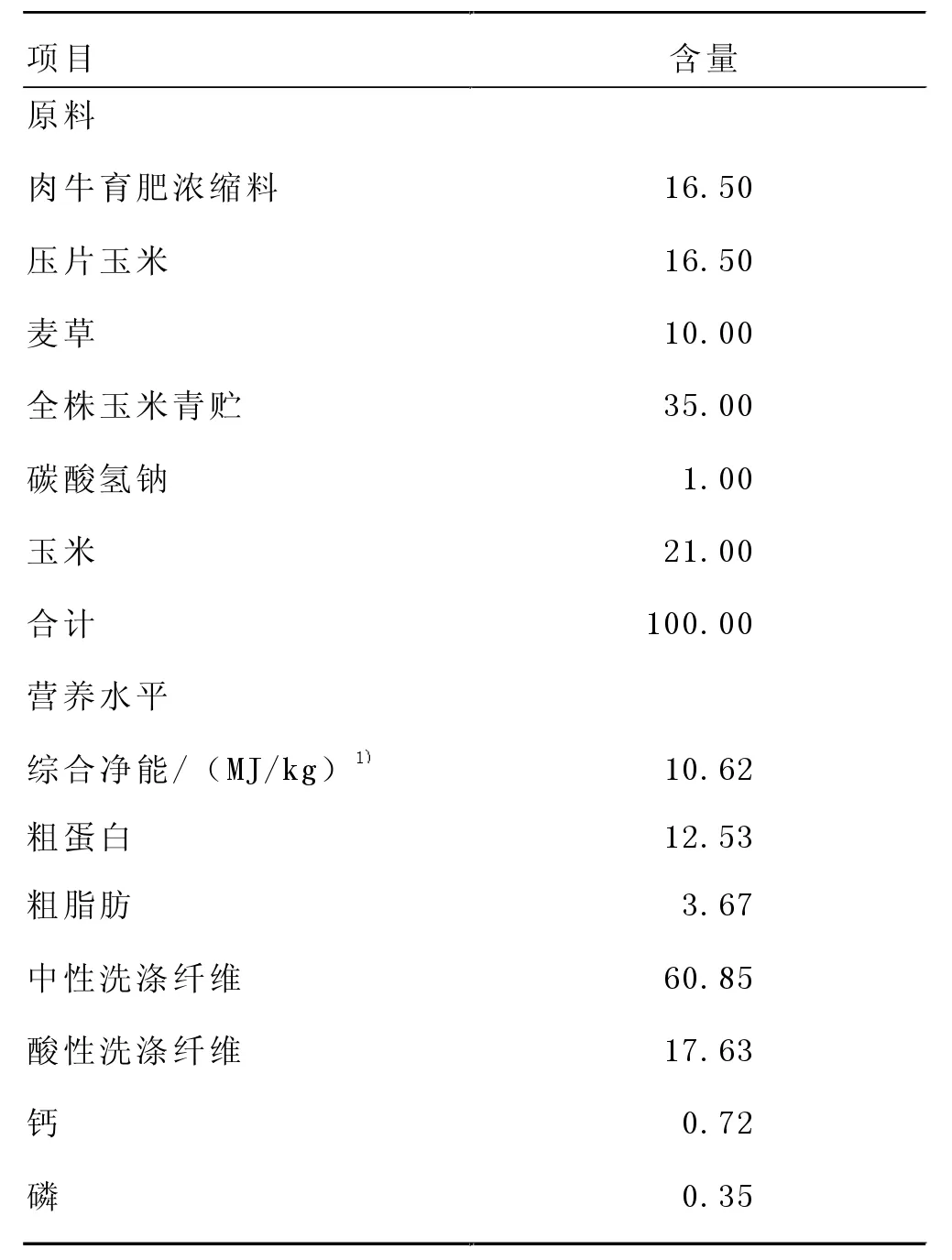
表1 試驗飼糧組成和營養水平(風干基礎)單位:%
1.2 飼養管理
試驗在吳忠市規模化牧場進行,對試驗牛進行編號,飼養在半開放式牛舍內,保持良好通風。 試驗開始前,對育肥牛進行驅蟲和耳標記。 飼喂相同的TMR 基礎飼糧,每天06:00 和17:00 分2 次飼喂,自由飲水。每天記錄每頭試驗牛的采食量和剩余飼料量,牛舍每周消毒1 次。
1.3 檢測指標
1.3.1 生產性能指標。 所有西門塔爾肉牛在試驗開始和結束時,用地磅稱量試驗牛早上空腹體重,記錄初始體重和最終體重,計算平均日增重(ADG);每天飼喂前清理料槽, 記錄采食量, 計算平均日采食量(ADFI)和料重比(F/G)。
ADG/(kg/d)=(終末體重-初始體重)/試驗天數;
ADFI/kg=總耗料量/(頭數×試驗天數);
F/G=總耗料量/總增重。
1.3.2 血清生化指標。 試驗結束當天,晨飼前采用普通10 mL 采血管于頸靜脈采集3 管血液, 靜置分層后,3 000 r/min 離心15 min, 然后分裝至1.5 mL 離心管中,于-80 ℃保存待測。 血清生化指標采用自動生化分析儀(Hitachi-7020,Hitachi Co,日本)進行分析,包括血清總蛋白(TP)、白蛋白(ALB)、尿素氮(UN)、葡萄糖(GLU)、堿性磷酸酶(ALP)和甘油三酯(TG)含量。
1.3.3 屠宰性能指標。 試驗期第90 天進行屠宰,宰前試驗牛禁食16 h,自由飲水。頸靜脈采血,血液經離心后制得血清;宰殺放血后,除去毛皮、頭、內臟及前肢膝關節和后肢趾關節以下部分, 靜置30 min 后稱量胴體重,分別測定骨重、油重和瘦肉重,計算凈肉重、屠宰率。
凈肉重=胴體重-骨重;
屠宰率/%=胴體重/宰前活重×100;
凈油率/%=油重/胴體重×100;
凈骨率/%=骨重/胴體重×100;
凈肉率/%=凈肉重/胴體重×100;
瘦肉率/%=瘦肉重/胴體重×100。
1.4 統計方法
試驗數據采用Excel 2010 進行整理, 用SPSS 17.0 統計學軟件中的ANOVA 過程進行單因素方差分析后, 采用Duncan’s 法進行多重比較檢驗,P<0.05表示差異顯著,P<0.01 表示差異極顯著,P>0.05 表示差異不顯著。
2 結果與分析
2.1 飼糧中添加RPG 對西門塔爾牛生長性能的影響
由表2 可知,與對照組相比,在育肥后期飼糧中添加RPG 顯著提升了西門塔爾牛終末體重和日增重(P<0.05),降低了料重比,但差異不顯著(P>0.05)。 西門塔爾牛育肥后期飼糧中添加RPG 可顯著改善肉牛的生產性能。
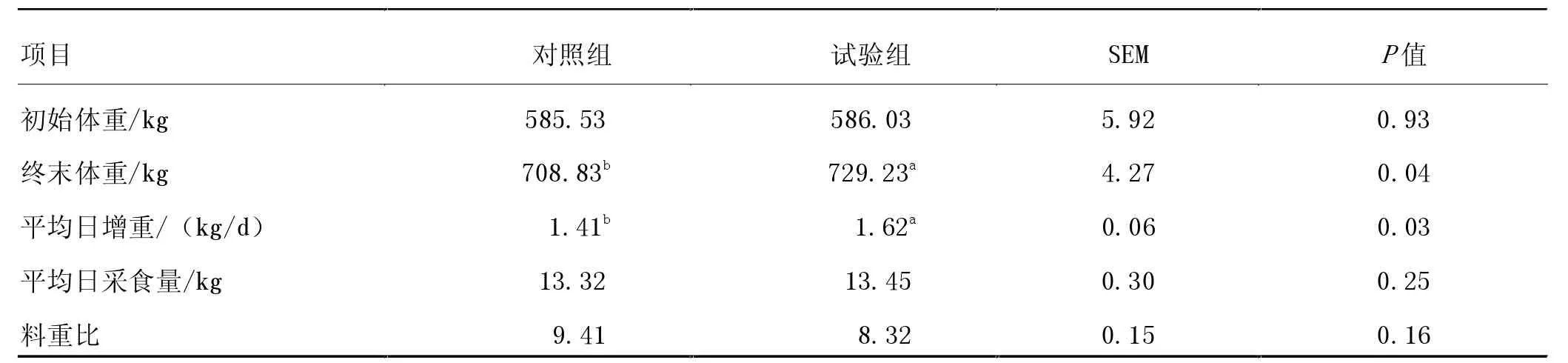
表2 飼糧中添加RPG 對西門塔爾牛生長性能的影響
2.2 飼糧中添加RPG 對西門塔爾牛血清生化的影響
由表3 可知, 與對照組相比,RPG 處理組顯著提高了肉牛育肥后期血清總蛋白含量(P<0.05),提高了血清ALB 含量,但差異不顯著(P>0.05);顯著降低了血清UN 含量(P<0.05)。
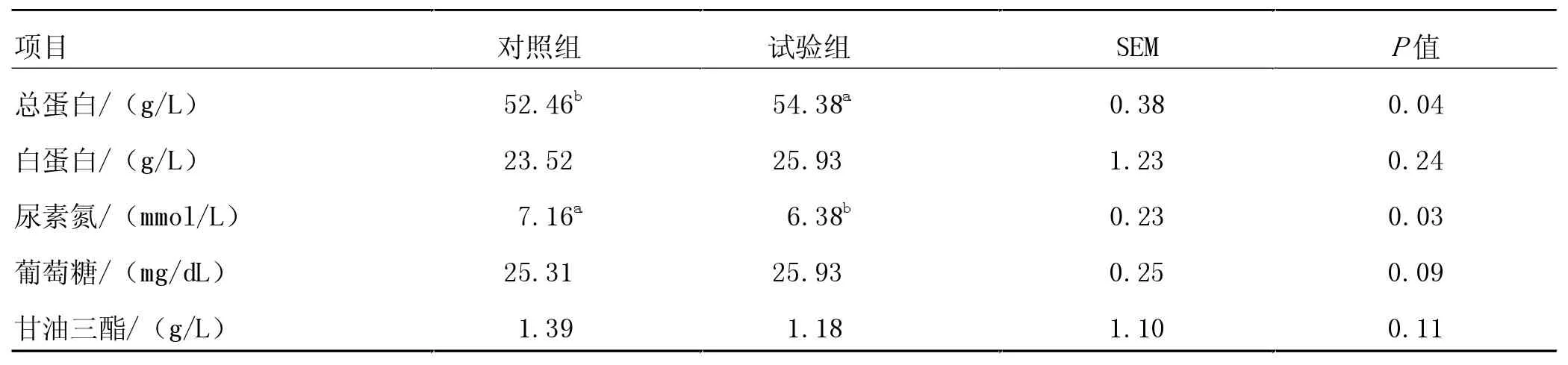
表3 飼糧中添加RPG 對西門塔爾牛血清生化的影響
2.3 飼糧中添加RPG 對西門塔爾牛屠宰性能和胴體品質的影響
由表4 可知,與對照組相比,西門塔爾牛育肥后期飼糧中添加RPG 顯著提高了宰前活重、胴體重、平均瘦肉重和屠宰率(P<0.05)。
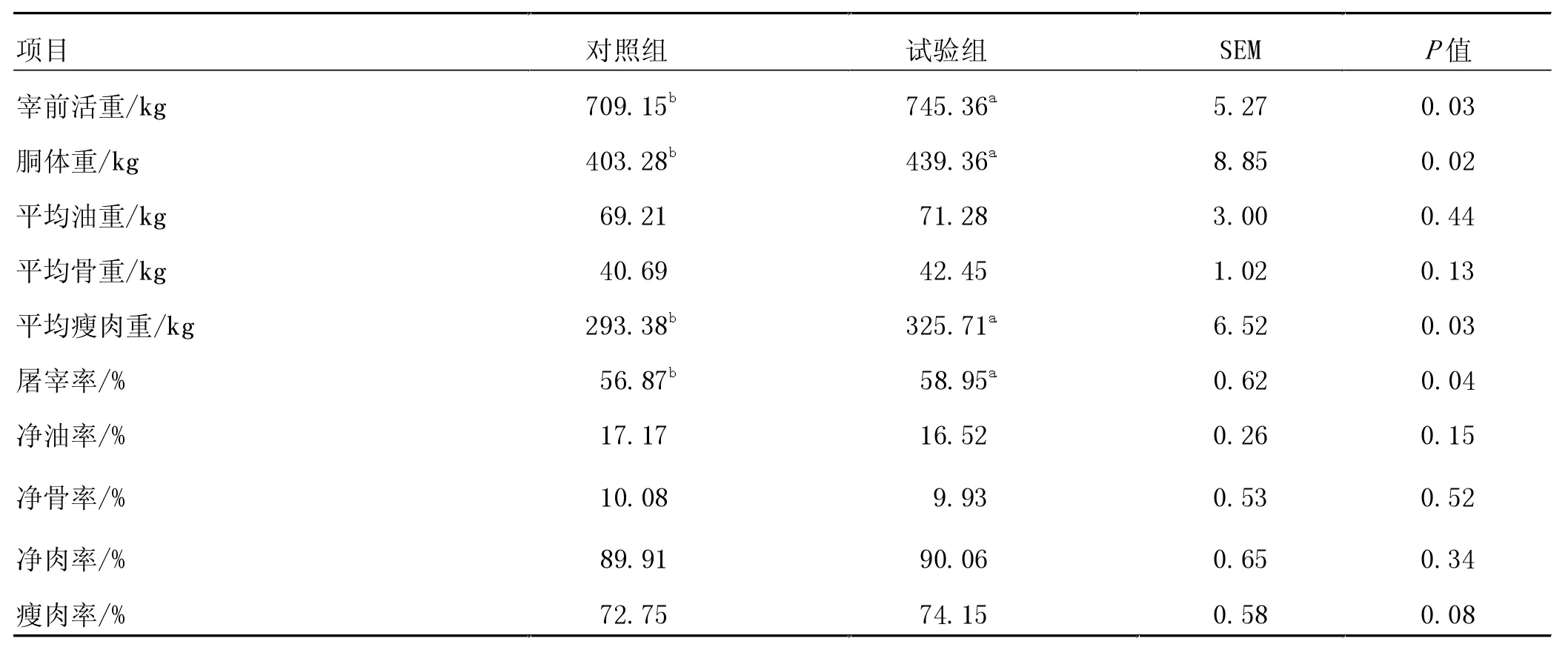
表4 飼糧中添加RPG 對西門塔爾牛屠宰性能和胴體品質的影響
3 討論
3.1 飼糧中添加RPG 對西門塔爾牛生長性能的影響
Cr 是一類自然存在于脊椎動物體內、能夠快速為肌肉和神經細胞提供能量的物質。GAA 作為機體合成Cr 的重要內源物,以甲基化的形式最終形成Cr,通過與高能磷酸基團結合以儲備能量[16]。 在日糧中添加GAA 可提高磷酸肌酸(PCr)和ATP 含量,為肌肉組織收縮、細胞活動及合成代謝提供更多的ATP,從而促進肌纖維的發育,改善動物的生長性能[17]。 本研究中,RPG 添加組與對照組相比, 可顯著提高肉牛的日增重,降低料重比,但差異不顯著。ADG 是肉牛生產中的一項重要指標,其高低決定了肉牛的生長速度,并影響著牛場的經濟效益。 與前人研究結果相似,晁雅琳等[18]研究表明,飼糧中添加800 mg/kg 和1 200 mg/kg GAA 可顯著提高舍飼灘羊ADG。 王子苑等[19]的研究表明, 飼糧中添加800 mg/kg GAA 可顯著提高肉牛ADG。 原因可能是動物機體在合成GAA 時需要精氨酸的參與, 而外源添加GAA 可有效減少體內精氨酸消耗量,這就為精氨酸參與機體內氨基酸與蛋白質的合成提供了有利條件,從而促進動物生長[20]。 牛血清TP 含量在一定程度上可反映動物機體的營養狀況和蛋白質代謝水平。 辛均平[21]在錦江黃牛日糧中添加0.4%的GAA,其血清中TP 含量顯著提高,本研究與其結果相似。 血清中TG 是脂肪代謝中的一種重要中間代謝產物, 其含量可作為評價脂肪代謝的指標[22]。當脂類消化吸收不良時,TG 含量明顯下降。 本試驗中, 在育肥后期西門塔爾牛飼料中添加0.34%的RPG,提高了血清ALB 含量,但差異不顯著。 血清UN的含量與蛋白質采食量及蛋白質利用率相關,當蛋白質采食量高或蛋白質利用率低時,血清UN 含量均會增加[23]。 本研究發現,在育肥后期西門塔爾牛飼料中添加0.34% RPG 組的牛血清UN 含量顯著低于對照組,說明飼糧中添加0.34% RPG 時,肉牛體內的氨基酸含量以及氮含量處于相對平衡狀態,可明顯提高蛋白質的利用率,這與劉永青等[24]在灘羊飼糧中添加包被蛋氨酸、李書杰等[25]在全混合顆粒料中添加不同類型蛋氨酸的研究結果相似。
3.2 飼糧中添加RPG 對西門塔爾牛屠宰性能和胴體品質的影響
胴體重、屠宰率是衡量產肉性能的重要指標。 本研究發現,在育肥后期西門塔爾牛飼料中添加0.34%RPG 可以顯著提高胴體重和屠宰率,這與劉笑梅等[26]的研究結果一致;同時還能夠明顯提高瘦肉率,減少體內脂肪沉積,這與張含等[27]的研究結果一致。 有研究表明,飼糧中添加GAA 能顯著提高肌肉中Cr 含量[28],同時血漿肌酸激酶(CK)活性也相應升高[29],在CK 催化下促使Cr 吸收多余ATP 的磷酸根基團形成PCr,從而提升磷酸原系統的能量儲備,為機體蛋白質沉積提供保障。 同時,GAA 作為氨基酸衍生物能夠促進肌肉的生長。 目前,精氨酸被認為是畜禽營養中的條件性必需氨基酸[30],具有促進骨骼肌蛋白合成、刺激生長激素分泌[31]以及增強機體抗應激能力等功能,由于精氨酸是合成GAA 的前體物,外源添加GAA 可以有效節約飼糧中的精氨酸[32],促使更多精氨酸參與機體的蛋白質合成,從而提高畜禽的生長性能[33]。 因此,在育肥后期西門塔爾牛飼料中添加0.34% RPG 一方面提高了動物肌肉中的Cr 水平, 促進了動物蛋白質的合成;另一方面可以補充外源Cr,減少用于合成Cr 的精氨酸數量, 使更多的精氨酸參與蛋白質的合成,使肉牛的肌肉生長量顯著提升,也使屠宰率和瘦肉率顯著提高。
4 結論
綜上所述,在育肥后期西門塔爾肉牛濃縮飼料中添加0.34% RPG 能夠顯著提高西門塔爾牛后期日增重、屠宰率、瘦肉率和血清TP 含量,顯著降低血清UN含量,從而提高肉牛的生產性能和屠宰性能,改善胴體品質。
猜你喜歡
中老年保健(2021年3期)2021-08-22 06:50:04
天津醫科大學學報(2021年2期)2021-03-29 05:31:08
昆明醫科大學學報(2021年1期)2021-02-07 01:06:36
現代臨床醫學(2021年1期)2021-01-26 00:56:02
昆明醫科大學學報(2020年12期)2021-01-26 00:44:04
中華養生保健(2020年4期)2020-11-16 01:31:40
中西醫結合肝病雜志(2020年2期)2020-10-27 02:18:50
豬業科學(2018年8期)2018-09-28 01:27:38
中成藥(2017年8期)2017-11-22 03:18:47
川北醫學院學報(2015年5期)2015-12-05 08:22:29
