磷酸鈣納米顆粒介導干擾LMO4對皮膚鱗癌細胞增殖的影響
2023-12-12 12:57:20項明華郭立鈺涂珍珍周海勝
安徽醫科大學學報 2023年11期
關鍵詞:檢測
項明華,郭立鈺,涂珍珍,王 月,周海勝
納米顆粒作為運送基因或藥物的載體,在生物醫學研究中得到廣泛應用。磷酸鈣納米顆粒(calcium phosphate nanoparticles, NP)作為一種新型的非病毒載體,可用于包裹目的基因轉染真核細胞[1]。LIM結構域蛋白4(lim-domain protein 4,LMO4)是有2個串聯重復排列的、富含有半胱氨酸-組氨酸的LIM結構域,故屬于LMO蛋白家族成員。LMO4作為核內轉錄因子,不與DNA分子直接發生作用,而是協助其他轉錄因子以調節靶基因的表達[2]。研究[3-6]表明在胚胎發育過程中,LMO4調控生殖細胞、表皮細胞和上皮細胞的增殖、分化和遷移。多項研究[7-9]表明上皮來源的多種腫瘤如乳腺癌、胰腺癌、肺癌和食管癌等腫瘤細胞中LMO4的表達顯著增加,與腫瘤細胞的增殖和凋亡存在密切關系。為了探討LMO4在皮膚鱗癌中作用和機制,分析皮膚鱗癌組織和細胞系中LMO4的表達水平,構建表達靶向干擾LMO4的shRNA,利用NP包裹這種表達載體,轉染皮膚鱗癌細胞,觀察其增殖能力并探討其機制。
1 材料與方法
1.1 試劑十二烷基硫酸納(sodium dodecyl sulfate, SDS)、瓊脂糖、枸櫞酸鈉、磷酸氫二鈉、氯化鈣和泡沫消除劑(聚醚F68)均為國產化學試劑(化學分析純),購自北京索來寶科技有限公司;高糖DMEM培養基和青-鏈霉素雙抗試劑購自美國Hyclone公司;胎牛血清(FBS)購自美國Gibco公司;細胞周期素蛋白(Cyclin)D1和E1抗體均購自美國Cell Signaling Technologies公司;GAPDH抗體、LMO4抗體、細胞周期素蛋白激酶(cyclin-dependent kinase, CDK)2和4均購自美國Abcam公司;檢測細胞增殖的噻唑藍試劑盒(thiazolyl blue, MTT)、細胞周期檢測試劑盒購自上海碧云天生物科技有限公司;引物(表1)均由上海生工生物工程有限公司合成;TRIzol試劑盒和轉染試劑Lipofectamine 3000購自美國Invitrogen公司;逆轉錄試劑盒、PCR檢測試劑盒均購自大連寶生生物公司。

表1 檢測目的基因引物
1.2 組織、細胞系與載體人皮膚鱗癌組織來自安徽省立醫院皮膚科,在完成臨床組織病理分析后剩余的標本;人正常皮膚組織來自安徽省立醫院泌尿外科門診包皮環切手術后遺棄的組織標本。人皮膚鱗癌細胞系A431和永生化的人角質形成細胞系(human immortalized epidermal cells, HaCat)均為所在研究室保存。干擾LMO4的干擾序列shRNA-L1和L2為研究室設計,載體GV248、干擾LMO4質粒shRNA-L1和shRNAc-L2均為課題組保存。
1.3 方法
1.3.1NP制備 根據文獻[1]提供方法制備NP。制備過程描述如下:將50 ml的0.05 mol/L氯化鈣、10 ml 1% SDS和10 ml 1% F68混合,持續攪拌24 h以制備均勻的微乳液A;同時將50 ml的0.025 mol/L磷酸氫二鈉、50 ml的0.025 mol/L枸櫞酸鈉、10 ml 1% SDS和10 ml 1% F68混合,攪拌24 h以制備均勻的微乳液B。再將微乳液B以10 ml/h 的速度緩慢加入到微乳液A中,并在35 ℃攪拌72 h,以促進兩種微乳液充分混合。將混合液15 000 r/min離心15 min獲得納米顆粒,75%乙醇洗滌4次,加入15 ml去離子水進行超聲勻化(頻率22 kHz,功率120 W)1 h。獲得的納米顆粒溶液置于-70 ℃冰箱中快速冷凍2 h,再轉入冷凍干燥箱放置48 h,從而獲得NP粉末備用。
1.3.2NP包被DNA及包裹效率分析 將制備的NP分別與shRNA-L1、shRNA-L2按照質量比10 ∶1、5 ∶1混合后,加入0.05 mol/L的氯化鈣重懸,室溫放置30 min,10 000 r/min離心10 min,收集沉淀備用。將上清液進行瓊脂糖凝膠電泳分析納米顆粒包裹DNA的效果。確定包裹方案,包裹載體GV248,作為納米顆粒介導的載體DNA(nanoparticles mediated vectors,NP/sh-V);包裹的NP/shRNA-L1和NP/shRNA-L2混合,制備納米顆粒介導的干涉LMO4混合液(nanoparticles mediated interfering LMO4, NP/sh-L)。制備好的納米包裹DNA液于4 ℃保存、備用。
1.3.3細胞培養與轉染 人皮膚鱗癌細胞系A431細胞和人皮膚角質形成細胞HaCat細胞均使用含10%熱失活的FBS和1%青-鏈霉素的高糖DMEM培養基,放置于37 ℃、5% CO2的培養箱中培養。按照脂質體Lipofectamine 3000說明書操作步驟,包裹shRNA-L1、shRNA-L2,并將二者混合形成脂質體介導的干涉LMO4(liposome mediated interfering LMO4, Lip/sh-L);同時使用脂質體包裹載體GV248(Lip/sh-V),分別轉染A431細胞。將納米顆粒包裹的NP/sh-L或NP/sh-V加入A431細胞,進行瞬時轉染,轉染的細胞數量和DNA量與脂質體轉染保持一致。轉染后培養24 h,更換新鮮培養基繼續培養,用于后續實驗。
1.3.4逆轉錄-定量PCR(RT-qPCR) 根據TRIzol試劑說明書提取細胞總RNA。參照試劑盒說明進行逆轉錄。PCR 反應條件為94 ℃變性4 min,按照94 ℃變性30 s,55 ℃退火30 s,72 ℃延伸30 s,共32個循環,最后72 ℃延伸8 min。每個樣品均設置β-actin作為內參基因。
1.3.5免疫組織化學 脫蠟處理組織切片,枸櫞酸鹽修復抗原,內源性過氧化物酶進行封閉。兔源抗體LMO4在4 ℃孵育過夜;次日依次滴加反應增強液和增強型酶標記二抗,室溫孵育30 min,DAB顯色,蘇木精染細胞核,自來水沖洗染料,烘干后使用中性樹脂封閉玻片;顯微鏡觀察和拍片。
1.3.6蛋白質免疫印跡(Western blot)實驗 收集細胞,加入細胞裂解液,冰上裂解30 min;細胞裂解液置于100 ℃水浴 10 min,使蛋白發生變性;進行12%聚丙烯酰胺-SDS電泳分離蛋白質;再以170 mA穩定電流轉膜1.5 h;含5%脫脂牛奶的封閉液封閉40 min;分別加入對應的抗體4 ℃孵育過夜;次日漂洗后加入酶標記二抗室溫孵育6 h,增強化學發光曝光顯影。
1.3.7細胞增殖實驗 用DMEM制備2×105/ml的細胞懸液,在96孔板中每孔接種500個細胞,接種后12 h(細胞貼壁),在12、24、36、48 h等4個時間點,根據MTT時試劑盒說明書操作。每孔加入10 μl新配制好的MTT溶液,在培養箱內孵育4 h,每孔加入100 μl的甲臜溶液,在培養箱內繼續孵育,直至甲臜完全溶解,在490 nm測定吸光度值(A490)。
1.3.8細胞周期分析 按照細胞周期檢測試劑盒說明書操作,簡要描述如下:收集細胞,制備單細胞懸液,離心后去除上清液,在細胞中加入70%預冷乙醇 500 μl 固定4 ℃過夜;去除固定液、洗滌細胞,細胞沉淀中加入100 μl RNase A溶液,重懸細胞,37 ℃水浴 30 min;再加入400 μl的碘化丙啶染色液混勻,4 ℃避光孵育30 min。流式細胞儀檢測分析細胞周期。
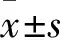
2 結果
2.1 LMO4在皮膚組織和細胞中表達利用收集的皮膚鱗癌組織和正常人皮膚組織,進行免疫組織化學分析。結果顯示(圖1A):LMO4表達定位在細胞核。在正常人皮膚組織中的表達水平較低,主要分布在表皮的基底層細胞,棘層、顆粒層和角化層等位置均沒有檢測到LMO4表達;而在皮膚鱗癌組織中,LMO4的表達水平顯著增加。
為了研究LMO4在皮膚鱗癌發生中的作用和機制,選擇了正常人皮膚來源的角質形成細胞HaCat和皮膚鱗癌細胞A431作為研究對象。首先在mRNA水平利用RT-qPCR方法對LMO4進行檢測。結果顯示(圖1B):皮膚鱗癌A431細胞中LMO4的表達水平顯著增加,是正常HaCat細胞的4倍。進一步在蛋白質水平利用Western blot進行驗證,結果顯示(圖1C):LMO4在A431的表達水平較HaCat明顯增加。因此,LMO4在皮膚鱗癌組織和鱗癌細胞中均具有較高的表達水平,由此推測LMO4可能參與皮膚鱗癌的發生、發展過程。
2.2 納米顆粒包裹DNA效率和干擾效率根據人LMO4的mRNA序列和表達載體GV248結構,設計并構建了2個可以表達干擾RNA的表達載體shRNA-L1和shRNA-L2(圖2A),用于干擾LMO4的表達。目的基因插入位置是在GV248載體的AgeI和EcoRI位點(圖2B)。將納米顆粒與DNA(shRNA-L1和shRNA-L2)按照質量比(μg)為5 ∶1和10 ∶1的比例進行混合,以包裹質粒DNA。將這些包裹的DNA溶液進行離心后,收集的上清液進行瓊脂糖凝膠電泳,以檢測上清液中殘留DNA,從而判斷包裹效率。如圖2C顯示:在5 ∶1的上清液中殘留較多DNA,包裹效率較低;而在10 ∶1組的上清液中未見殘留DNA,顯示較好的包裹效果。在后續實驗中,將納米顆粒按照10 ∶1包裹DNA。

圖2 shRNA表達載體構建及NP包裹DNA檢測
為了檢測轉染效率和干涉效率,分別將納米顆粒包裹的DNA(NP/sh-L和NP/sh-V)和脂質體包裹DNA (Lip/sh-L和Lip/sh-V)瞬時轉染A431細胞,提取細胞總RNA進行RT-qPCR檢測LMO4的表達水平。結果顯示(圖3A):在Lip/sh-L組細胞和NP/sh-L組細胞中LMO4的表達水平,分別是A431細胞組的64%±12.5%和55.0%±14.3%,分析顯示差異有統計學意義(P=0.002 1,P=0.03)。但是Lip/sh-L組細胞與NP/sh-L組細胞相比,LMO4的表達差異無統計學意義(P=0.21)。Western blot檢測結果脂質體和NP轉染的干擾載體,都能降低LMO4的表達(圖3B)。灰度分析顯示(圖3C):Lip/sh-L組中LMO4的表達水平是A431組細胞的74.0%±4.5%,差異有統計學意義(P=0.024)。NP/sh-L轉染組的細胞中LMO4的表達水平是A431細胞的51.0%±3.0%,差異有統計學意義(P=0.002)。可見Lip/sh-L和NP/sh-L轉染的細胞,LMO4的表達均得到有效抑制。

圖3 A431細胞LMO4干擾效率分析
2.3 干擾LMO4對A431細胞增殖和細胞周期的影響為了研究干擾LMO4的表達對A431細胞增殖的影響,使用MTT法檢測細胞增殖能力,結果顯示(圖4):在轉染后24、36和48 h,NP/sh-L組吸光度值(A490)分別為0.28±0.01、0.31±0.11、0.38±0.18;NP/sh-V組的A490分別為0.39±0.02、0.54±0.02、0.73±0.11。與A431組相比,NP/sh-L組A490在24 h(P=0.011)、36 h(P=0.002)和48 h(P=0.001)等時間點的表達差異均有統計學意義。因此,A431細胞在干擾LMO4時,其細胞的增殖能力顯著降低。

圖4 干擾LMO4對細胞增殖能力的影響
為了進一步研究LMO4調節細胞增殖的機制,采用碘化丙啶染色DNA方法,通過流式細胞分析技術觀察細胞周期的變化。結果顯示(圖5A):在A431組細胞中,處于G0/G1期的細胞比例為30.76%±1.96%;NP/sh-L組細胞中G0/G1期細胞比例為39.82%±1.86%,二者相比,差異有統計學意義(P=0.003)。A431組細胞中S期的細胞的比例為50.65%±0.62%,NP/sh-L組細胞中S期細胞比例為39.56%±0.65%,二者相比,差異有統計學意義(P=0.002)。而G2/M期細胞在NP/sh-L組與A431組之間比較,二者的差異無統計學意義(P=0.517)。可見干擾LMO4可以阻止細胞從G1期進入S期。

圖5 干擾LMO4對A431細胞周期的影響
為探討LMO4調控細胞周期的機制,利用RT-qPCR方法對Cyclin D1、Cyclin E、CDK2和CDK4進行了檢測。結果顯示NP/sh-L組細胞中Cyclin E和 CDK2較對照組明顯降低。其中,Cyclin E是NP/sh-V組細胞的0.36±0.03倍,差異有統計學意義(P=0.016);NP/sh-L組細胞中CDK2是NP/sh-V組細胞的0.49±0.02倍,差異有統計學意義(P=0.001)。干擾LMO4的表達對Cyclin D1和CDK4的表達沒有明顯影響(圖5B)。Western blot方法檢測Cyclin D1、Cyclin E及CDK2和CDK4等蛋白的表達變化,與RT-qPCR的結果一致(圖5C)。因此,LMO4可以通過調控Cyclin E和 CDK2表達,影響細胞周期的變化。
3 討論
將目的基因轉入真核細胞,常見的有脂質體轉染、磷酸鈣轉染、電轉染或病毒介導等方法,因操作過程較為繁瑣,要求具有較高的實驗條件,且需要較高的實驗成本,在應用中受到一定限制。利用磷酸鹽和氯化鈣聚合形成直徑為23.5~34.5 nm的納米顆粒,具有易制備、實驗成本低和較低的細胞毒性,且不具有免疫原性等優點[1],具有一定應用價值。在氯化鈣修飾制備的納米顆粒時,改變了納米顆粒表面帶電性,包裹帶負電的DNA時能提高包裹效率[10]。利用此方法制備的納米顆粒可以有效包裹干擾載體DNA,與傳統的脂質體介導基因轉移相比,其轉染效率和干擾效率具有較好的一致性。
LMO4作為核內轉錄因子,協助其他轉錄因子以調節靶基因的表達[2]。由于胚胎發育時LMO4參與調控表皮細胞的增殖、分化和遷移[3-6];而在正常皮膚組織中,LMO4僅僅在基底層細胞中表達,其功能是參與維持角質形成細胞的增殖、分化和遷移,保持表皮層細胞的更新[11]。有研究[12-14]表明在上皮組織來源的腫瘤,如乳腺癌、非小細胞型肺癌、鱗狀細胞癌、胰腺癌等腫瘤中,LMO4的表達水平均顯著增加,且LMO4參與調控這些腫瘤細胞的增殖、遷移和轉移。因此,LMO4與腫瘤細胞惡性生物學行為密切相關。免疫組織化學檢測證實LMO4在皮膚鱗癌組織顯著增加;細胞水平檢測也表明LMO4在皮膚鱗癌細胞中表達水平較正常皮膚角質形成細胞顯著增加。提示LMO4可能參與調控皮膚鱗癌的發生。
在皮膚鱗癌細胞系A431中靶向干擾LMO4的表達,結果顯示敲低LMO4的表達,能顯著抑制A431細胞的增殖。在干擾LMO4表達的細胞中,G2/M期細胞沒有受到影響,而主要變化是G0/G1期細胞數量顯著增加,S期細胞較對照組細胞明顯減少。因此,干擾LMO4表達,有利于細胞周期停滯在G0/G1期。已知調節細胞周期的主要是周期素蛋白包括有 A、B、D和E等5種類別。其中,Cyclin D1和E主要調控細胞G1期到S期,Cyclin D1能激活CDK4調控G1期細胞生長;Cyclin E激活CDK2調控S期的染色體復制[12]。在A431細胞中干擾LMO4時,Cyclin E表達顯著降低,同時伴隨CDK2表達減少;而Cyclin D1和CDK4的表達變化不明顯。這些結果與細胞周期的變化具有一致性。有研究[15]表明在乳腺癌細胞中敲低LMO4后,Cyclin D1和E的表達顯著降低,導致細胞停留在G2/M期,這可能是LMO4在不同類型腫瘤細胞中作用機制存在差異性。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
