褪黑素調控自噬抑制肝星狀細胞增殖的作用及機制研究
2023-12-12 12:57:16陳滌非黃啟鳴徐德祥任曉非洪汝濤
安徽醫科大學學報 2023年11期
陳滌非,揭 磊,黃啟鳴,徐德祥,任曉非,洪汝濤
肝纖維化( hepatic fibrosis,HF)是各類致病因素引起肝臟損傷-修復反應的結果,其主要病理改變為肝內彌漫性細胞外基質(extracellular matrix,ECM)的過度沉積,是多種慢性肝病向肝硬化進展的共同基礎[1]。研究[2]表明,活化的肝星狀細胞(hepatic stellate cells,HSCs)在HF發展的進程中起到舉足輕重的作用。研究[3]表明,血小板源性生長因子-BB(platelet-derived growth factor-BB,PDGF-BB)是較強致炎因子之一,在HSCs激活中起重要作用。在這一過程中,溶酶體發揮積極作用[4]。有文獻[5]提示,自噬在HF進展中發揮重要作用,但其具體機制還不清楚。HF作為肝病發展的共同通路,逆轉HF是至關重要的,如果通過調節細胞自噬,減少HSCs活化,增強對肝細胞的保護和再生,就可以減緩病程甚至逆轉HF。該課題組前期研究表明,在肝組織中,褪黑素(melatonin,MEL)可以抑制自噬從而減輕HF[6]。目前自噬與HSCs增殖關系及MEL對其抑制作用除該課題組外尚未見報道,該研究將進一步研究MEL抑制HSCs增殖與抑制自噬水平的相關性。
1 材料與方法
1.1 細胞與試劑HSC T-6(永生系細胞,武漢普諾賽公司);DMEM高糖培養基、胎牛血清(杭州四季青生物有限公司);PDGF-BB(美國PeproTech公司);抗微管相關蛋白1輕鏈3b(microtubule-associated protein 1 light chain-3b,LC3b)抗體、α-平滑肌肌動蛋白(alpha-smooth muscle actin,α-SMA)兔抗鼠單克隆抗體、抗β-actin抗體(美國Abcam公司);二甲基亞砜(dimethyl sulfoxide,DMSO)、苯甲基磺酰氟(phenylmethylsulfonyl fluoride,PMSF)、青霉素、鏈霉素、胰酶消化酶、雙重縮氨酸法(bicinchoninic acid assay,BCA)試劑盒、聚偏氟乙烯(polyvinylidene fluoride,PVDF)膜(上海碧云天生物技術有限公司);MEL(美國Sigma公司);細胞計數試劑盒-8(cell counting Kit-8,CCK-8)、山羊抗小鼠辣根酶標記IgG(H+L)、4%多聚甲醛、1%四氧化鋨(合肥Biosharp生物科技公司)。
1.2 細胞培養細胞培養HSC-T6細胞株復蘇后培養于含10%胎牛血清、1%青-鏈霉素的DMEM培養基中,置37 ℃、5% CO2的培養箱中培養至70%~80%融合后傳代,取對數生長期細胞用于實驗。
1.3 CCK-8實驗用3~4代細胞,共設置5組:對照組、模型組、MEL(低、中、高劑量)處理組。HSCs接種于96孔培養板(濃度5×104/L),每孔設6個復孔,培養24 h后細胞貼壁,約70%~80%細胞融合,換用無血清的DMEM培養基,以使細胞同步化于G0期,再培養24 h后除對照組外各組加入PDGF-BB(終濃度10 ng/ml)、MEL處理組加入MEL(低劑量組為 1 nmol/L,中劑量組為1 μmol/L,高劑量組為0.1 mmol/L),孵育48 h后加入CCK-8(10 g/L)20 μl,繼續37 ℃培養2 h,小心吸棄培養液。酶標儀測波長為570 nm處的吸光度(A)值。
1.4 Western blot實驗同上述CCK-8分組,將細胞接種于培養瓶中,于恒溫培養箱中培養,待細胞長至單層80%左右時,同步化處理,各分組加入處理因素(PDGF-BB和MEL,濃度同CCK-8),培養48 h,小心吸棄培養液,加入PBS反復吹打細胞至細胞基本脫落,離心(650 r/min,5 min),棄去上清液,加入細胞裂解液并反復吹打細胞使細胞與裂解液充分混勻,靜置30 min離心,提取總蛋白。蛋白定量后每組上樣10 μg,進行12.5%聚丙烯酰胺凝膠電泳,轉膜封閉。將一抗稀釋至所需濃度(兔抗鼠LC3b 1 ∶2 000、兔抗鼠α-SMA 1 ∶1 000、 β-actin 1 ∶3 000),4 ℃孵育過夜。洗膜3次后,加入二抗,將其濃度稀釋至1 ∶8 000,洗脫二抗,ECL發光顯色。同時檢測β-actin表達作為內參。拍照掃描后Image J圖像分析軟件進行灰度值分析,計量以平均吸光度值×面積來表示,以LC3 b/內參β-actin的灰度值比值作為相對表達量。
1.5 qRT-PCR實驗同上述CCK-8分組,將細胞接種于培養瓶中,融合度為80%左右時同步化處理24 h后分別加入各處理因素(PDGF-BB和MEL,濃度同CCK-8)培養48 h后使用TRIzol試劑提取HSCs總RNA,用紫外分光光度法測定RNA水平,A260/A280鑒定RNA純度。分別提取1 μg總RNA,經RNA反轉錄酶催化合成cDNA第一鏈反轉錄體系:MgCl225 mmol/L 2.2 μl,10×PCR Buffer II 1.0 μl,dNTP 10 mmol/L 2.0 μl,Oligo(dt)18 50 μmol/L 0.5 μl, RNase Inhibiter 20 U/μl 0.25 μl,RNase-free H2O 1.85 μl,50 ng/μl RNA 2.0 μl,總體積 10 μl。反轉錄條件:25 ℃ 10 min, 48 ℃ 60 min, 95 ℃ 5 min,以此為模板進行實時熒光定量PCR擴增,PCR反應系統采用SYBRG,按操作程序進行,反應總體系為20 μl,反應條件為50 ℃ 120 s,1個循環;95 ℃ 600 s,1個循環;55 ℃ 15 s,40個循環。60 ℃ 60 s,標記熒光,得出各樣本的初始拷貝數。所用引物序列為:LC3b,上游 CCCAGACATCAGGGAGTAATGG,下游 TCTATCGGATACTTCAGCGTCA;GAPDH,上游CGGAGTCAACGGATTTGGTCGTAT,下游 AGCCTTC-TCCATGGTGGTGAAGAC;α-SMA,上游 CCCAGACA-TCAGGGAGTAATGG,下游 TCTATCGGATACTTCA-GCGTCA。實驗過程中以GAPDH內參基因作為實時熒光RT-PCR的質控。每組樣本一式2份,在指數期內設定參數值,得到每個PCR反應的循環域(cyclethreshold, Ct)值,即模板cDNA經PCR擴增后達到某一熒光強度的循環次數。以每例細胞的目的片段與含內參抗體片段的Ct值作為該標本基因表達的相對數值,以對照組的表達量為100%計算其余各組的表達量。
1.6 透射電鏡觀察HSCs的超微結構同上述 CCK-8分組,將生長狀態良好的HSCs均勻接種于6孔板中。融合度為80%左右時同步化處理24 h后分別加入各處理因素(PDGF-BB和MEL,濃度同CCK-8)培養48 h。干預完成后,各組細胞分別按細胞傳代培養的方法將細胞制成細胞懸液。1 000 r/min離心5 min,使細胞沉淀到離心管底部。棄掉上清液PBS,洗滌后加入4%多聚甲醛固定液固定 2 h。1 000 r/min離心5 min沉淀細胞,棄掉多聚甲醛固定液,留取細胞沉淀。沿著離心管的壁用勾針輕輕的把離心沉淀的細胞團勾離管壁,加入1%四氧化鋨進行處理。勾針輕輕取出細胞團,小心切成約1 mm3的小塊。脫水包埋浸透4 h。對固化的細胞小塊進行超薄切片,厚度70 nm。枸櫞酸鉛染色。通過透射電鏡(Talos L120C G2美國賽默飛公司)觀察HSCs超微結構,每組內每個樣本挑選3個視野進行計數,以圖片下方2 μm標尺為標準,統計每張圖片內自噬溶酶體個數。
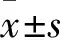
2 結果
2.1 MEL對PDGF-BB激活的HSCs細胞增殖的影響結果顯示:與對照組比較,PDGF-BB促進模型組HSCs活化、增殖(P<0.01);與模型組比較,MEL可以抑制HSCs增殖(P<0.01)。見表1。

表1 MEL對激活的HSCs增殖的影響
2.2 MEL對PDGF-BB激活的HSCs LC3b蛋白的影響Western blot結果顯示模型組HSCs中LC3b蛋白表達水平相對于對照組明顯增加(P<0.05);相對于模型組,在MEL干預的處理組中,LC3b蛋白表達水平顯著降低(P<0.05)。見圖1。

圖1 MEL對LC3b蛋白表達的影響
2.3 MEL對PDGF-BB激活的HSCs α-SMA蛋白的影響Western blot結果顯示模型組HSCs中α-SMA蛋白表達水平相對于對照組明顯增加(P<0.05);相對于模型組,在MEL干預的處理組中,α-SMA蛋白表達水平顯著降低(P<0.05;P<0.01)。見圖2。

圖2 MEL對α-SMA蛋白表達的影響
2.4 MEL對PDGF-BB激活的HSCs LC3b mRNA表達水平的影響qRT-PCR結果顯示模型組HSCs中LC3b mRNA表達水平相對于對照組顯著增加(P<0.01);相對于模型組,在MEL干預的處理組中,LC3b mRNA表達水平顯著降低(P<0.01)。見圖3。

圖3 MEL對LC3b mRNA表達的影響
2.5 MEL對PDGF-BB激活的HSCs α-SMA mRNA
表達水平的影響qRT-PCR結果顯示模型組HSCs中α-SMA mRNA表達水平相對于對照組明顯增加(P<0.05);相對于模型組,在MEL干預的處理組中α-SMA mRNA表達水平顯著降低(P<0.01)。見圖4。

圖4 MEL對α-SMA mRNA表達的影響
2.6 MEL對PDGF-BB激活的HSCs自噬溶酶體的影響圖5A雙核膜結構清晰,核周隙正常,核內染色質未見明顯凝集。胞質內可見少量自噬溶酶體。圖5B雙核膜結構清晰,核周隙正常,核內染色質未見凝集,胞質內可見大量自噬溶酶體。圖5C雙核膜結構清晰,核周隙正常,核內染色質可見輕度邊集化。胞質內可見少量自噬溶酶體。圖5D雙核膜結構清晰,核周隙正常,核內染色質未見明顯凝集,胞質內自噬溶酶體數量顯著降低。

圖5 各組HSCs超微結構 ×50 000
透射電鏡結果顯示模型組HSCs中自噬溶酶體相對于對照組明顯增加(P<0.05);相對于模型組,在MEL(1 μmol/L; 0.1 mmol/L)干預的處理組中自噬溶酶體顯著減少(P<0.01),與模型組比較,MEL(1 nmol/L)差異無統計學意義。見圖6。

圖6 MEL對自噬溶酶體數量表達的影響
3 討論
肝纖維化是多種慢性肝病引起的病理過程,肝硬化是HF的最終結局,并伴隨多種并發癥及肝功能紊亂,具有很高的病死率[6]。近年來,相關研究[7]表明,HF甚至是早期肝硬化是可以逆轉的。相關研究[8]表明,HSCs是ECM形成的主要來源,且HSCs激活轉化為肌成纖維細胞是HF發生發展的中心環節。HSCs的激活伴有NF-kb轉錄因子的激活及cymb基因表達增強,而cymb蛋白可結合α-SMA基因調控區,表達α-SMA。所以α-SMA作為HSCs活化的主要標志物,與HSCs活化的程度及數量呈正相關[9]。CCK-8實驗目前是一種方便、快速和準確的細胞代謝活性評估工具,可以用于定量評估細胞增殖和存活率。該實驗采用HSC-T6進行體外培養,用PDGF-BB激活HSCs,通過CCK-8和有關分子生物學實驗,從細胞和分子水平表明,HSCs數量和活性明顯增強,HSCs α-SMA蛋白質、mRNA表達水平明顯增高,說明PDGF-BB已活化HSCs,為后續實驗提供了基礎。
自噬是真核細胞中廣泛存在的一種代謝過程。正常細胞中,胞內衰老的細胞器及其他結構被自噬體吞噬并轉運至線粒體進行β氧化,提供能量。微管相關蛋白1輕鏈3(microtubule-associated protein 1 light chain 3,LC3)/Atg8是目前研究中被最廣泛使用的分子標志物。它參與了自噬體膜的形成,包括可相互轉化的兩種形式LC3 Ⅰ和LC3 Ⅱ。新合成的LC3經過加工形成可溶形式的LC3 I,后者經過泛素化加工修飾,與自噬體膜結合,成為膜結合形式的LC3 Ⅱ[10]。因而LC3 Ⅱ的含量或者LC3 Ⅱ/LC3 Ⅰ的比例與自噬體的數量呈正相關,在某種程度上反映了細胞的自噬活性。自噬的關鍵是自噬小體的形成,在透射電鏡觀察到自噬小體結構是檢測自噬的金標準。
研究[1]表明,自噬與HF發生發展有關,但其與HSCs關系不甚清楚。正常情況下,HSCs處于靜息或者未激活狀態,胞質中聚集著包含視黃醇和三酰甘油的脂滴[9]。當存在損傷因子的刺激時,HSCs中的脂滴會被自噬過程所識別,隨之被溶酶體降解,釋放游離脂肪酸[11],后者為HSCs轉化為肌成纖維細胞提供能量來源,進而釋放促纖維化因子,如α-SMA和膠原蛋白[4]。上述研究表明脂滴的降解與HSCs的激活有密切的關系。有體內研究[12]表明,抑制自噬可以引起脂滴在HSCs胞質內聚集,HSCs處于沉默狀態,并且能減弱HF。該實驗結果顯示,PDGF-BB處理后活化的HSCs LC3b蛋白質、mRNA表達水平明顯增高,初步說明HSCs自噬水平上調。進一步采用透射電鏡對PDGF-BB激活的HSCs進行超微結構分析,顯示其自噬溶酶體數量顯著增加,提示PDGF-BB激活的HSCs自噬水平明顯上調。
MEL是一種具有多種生理功能的內源性激素,特別是其抗氧化作用,已被證明能夠通過調節多條信號通路來抑制HF的發展,包括影響ECM的合成和降解[13]。該課題組前期體內實驗[6]表明MEL能抑制肝纖維化,同時下調自噬水平。在此基礎上,該研究進一步探討在PDGF-BB激活的HSCs中,MEL抑制HSCs增殖與抑制自噬水平是否相關。該研究結果顯示,PDGF-BB處理的HSCs中,細胞增殖和活性明顯增強, α-SMA、LC3b蛋白質、mRNA表達明顯增強,細胞內自噬溶酶體數量明顯增加,說明PDGF-BB能活化HSCs,同時上調自噬水平,而經MEL處理后,HSCs增殖被抑制,其內α-SMA、LC3b蛋白質、mRNA表達水平明顯下降,HSCs內自噬溶酶體數量顯著減少,說明MEL能抑制HSCs活化且下調其自噬水平。因此,MEL可以抑制HSCs的活化,其機制可能與MEL下調HSCs的自噬水平相關,這為治療HF提供了新的臨床思路。但該實驗僅為初步研究,有待于進一步深入研究。研究[14]表明自噬參與HF的通路主要有TGF-β1/Smads通路,P13K/AKT/mTOR信號通路,RAF/MEK/ERK信號通路,但其確切的作用和功能機制有待進一步研究。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
人大建設(2019年12期)2019-05-21 02:55:32
光學精密工程(2016年6期)2016-11-07 09:07:19
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
