奧密克戎的感染、免疫及其免疫逃逸機制的研究進展
2023-12-24 18:16:00詹廣耀冼柏俊黃姝賢黎劍橋梁君如廖釗宏
廣東藥科大學學報 2023年6期
詹廣耀,冼柏俊,黃姝賢,黎劍橋,梁君如,廖釗宏
(佛山科學技術學院醫(yī)學院檢驗系教研室,廣東 佛山 528000)
新冠病毒感染容易致使人患上新型冠狀病毒性肺炎,對人體肺部功能造成巨大損傷,甚至致死,即使是治愈也可能留下肺纖維化等后遺癥。2020年1 月,我國疾控中心成功分離出首株新冠病毒毒種;2020 年2 月11 日,世界衛(wèi)生組織(WHO)正式將新型冠狀病毒肺炎命名為COVID-19,引起COVID-19 的病毒被稱為2019-nCoV,后被修訂為SARSCOV-2。
2021 年,我國成功開發(fā)和使用多種新冠病毒疫苗來抵抗病毒的感染,通過接種新冠疫苗激發(fā)主動免疫獲得了針對新冠病毒特異性的免疫力。但是新冠病毒也在利用自身單鏈RNA病毒的特性,不斷突變出新的變異株,不斷考驗著人體的免疫系統(tǒng)。據(jù)統(tǒng)計,2019年到2023年新型冠狀病毒已經(jīng)迭代變異出了多種變異株,其中常見的分別有阿爾法變異毒株、貝塔變異毒株、伽馬變異毒株、德爾塔變異毒株和奧密克戎變異毒株。
目前,國內普遍感染新冠病毒的種類為Omicron,Omicron 相較原始病毒具有隱匿性強、傳播速度快、傳播能力強、致病力相對較弱等特點。同時Omicron也是目前已知的具有最強逃逸能力的新冠病毒變異株,許多科研工作者投入大量資金和精力研究Omicron,了解其逃逸的機制和特性。本文對Omicron 的結構、入侵人體細胞的機制與危害性、人體T、B、NK 細胞抗病毒免疫、Omicron 對人體免疫細胞(T、B、NK細胞)的逃逸機制的相關研究等進行了回顧分析,為后續(xù)對Omicron 感染預防和相關藥物研發(fā)提供參考。
1 Omicron的特征與危害性
Omicron是于2021年11月9日在南非地區(qū)被檢測出來的新型冠狀病毒的一種變異毒株,屬于β-冠狀病毒。Omicron 由于其基因突變導致其胞膜蛋白中部分表位發(fā)生變化,其在宿主間傳播能力更強,在宿主體內隱匿能力也得到加強,可引起呼吸、消化、泌尿、神經(jīng)、生殖等系統(tǒng)的器官損傷。
1.1 Omicron病毒結構特征
Omicron 作為新型冠狀病毒的一種變異株,其結構和特征與其原始毒株具有一定的相似性。Omicron 是一種具有包膜的單股正鏈RNA 病毒,其病毒直徑為60~140 nm,形態(tài)呈圓形或橢圓形。其結構蛋白成分有4種,由外到內分別是:位于病毒顆粒包膜最外圍的刺突糖蛋白(spike protein,S),組成病毒顆粒包膜的小包膜糖蛋白(envelope protein,E)和膜糖蛋白(membrane protein,M),以及位于病毒顆粒中心、與病毒RNA 基因鏈結合的核衣殼蛋白(nucleocapsid protein,N)[1]。
Omicron 的遺傳物質由RNA 基因鏈和核衣殼蛋白組成。其RNA 基因組長度約為30 kb,具有2個重疊的開放閱讀框ORF1a 和ORF1b,可以編碼16 個非結構蛋白(Nsp1~16)和4 種結構蛋白及9 種輔助蛋白,即ORF3a、ORF3b、ORF6、ORF7a、ORF7b、ORF8、ORF9b、ORF9c、ORF10。與原始毒株比較,Omicron具有多達60個突變,其中37個是S蛋白的突變,且有15個突變點位于受體結合結構域(receptor binding domain,RBD)上[2]。除了S 蛋白,在Omicron 其他結構蛋白也發(fā)現(xiàn)了突變,包括E 蛋白上T9I 突變,M 蛋白上D3G、Q19E、A63T 突變和N蛋白上P13L、Δ31 33、R203K、G204R突變[3]。
S 蛋白是新冠病毒主要的膜表面蛋白,負責介導病毒顆粒與宿主細胞上的血管緊張素轉化酶2(angiotensin converting enzyme 2,ACE2)受體結合,引導病毒顆粒進入宿主細胞[4],同時目前大部分疫苗都是基于S 蛋白的多表位肽疫苗。Omicron 的S蛋白RBD 中存在的G446S、S477N、T478K、E484A、Q493R、G496S、Q498R、N501Y、Y505H、K417N 和N440K 氨基酸突變,這使得Omicron S 蛋白的RBD 和S 蛋白都擁有更多的疏水氨基酸和α-螺旋結構,這些突變最終導致Omicron 的S 蛋白具有更加穩(wěn)定的結構[5]。
M 蛋白在新型冠狀病毒中以二聚體形式存在,是含量最多的結構蛋白。新冠病毒的M 蛋白含有222 個氨基酸,由1 個短的氨基末端域、中間的多次跨膜結構域和1個長的羧基末端結構域組成。且新冠病毒M 蛋白上有aa21-36、aa46-6 和aa76-6,3 個α-螺旋疏水性跨膜域和2 個固有無序區(qū)域,分別為aal-7 和aa205-222,這2 個固有無序區(qū)域具有轉錄調節(jié)、細胞信號轉導等重要功能。新冠病毒顆粒上的M 蛋白決定著病毒包膜的形態(tài),同時引導著病毒顆粒在宿主細胞體內組裝和釋放[6]。
Omicron的N蛋白以二聚體形式存在,包括1個N 端結構域(N-terminal domain,NTD)和1 個羧基端結構域(carboxyl-terminal domain,CTD)。其NTD與病毒的RNA 非特異性結合,能識別病毒基因組的3′端核苷酸序列[7]。同時N 蛋白的突變導致了其磷酸化位點的變化,形成了可以和14-3-3 蛋白結合的磷酸化依賴結構域。這將導致宿主細胞內負責控制細胞周期、細胞存活和細胞死亡的14-3-3 蛋白被結合進而使宿主細胞在被感染后其細胞周期變長,更利于Omicron在宿主細胞內部的復制[8]。
1.2 Omicron侵入人體細胞的機制
Omicron 作為新冠病毒的一個變體,其入侵人體細胞的主要途徑為兩種(圖1):一種是由跨膜絲氨酸蛋白酶2(transmembrane protease serine2,TMPRSS2)切割S 蛋白后的細胞膜融合進入途徑;另一種是由內體蛋白酶組織蛋白酶L或B介導的內體膜融合進入途徑[9]。在TMPRSS2 切割S 蛋白后的細胞膜融合進入途徑中,Omicron 包膜上的S 蛋白是在入侵宿主細胞過程中發(fā)揮主要作用的功能蛋白。當Omicron 病毒顆粒接近宿主細胞時候,S蛋白的S1 結構域上的受體結合結構域RBD 就會與宿主細胞膜上的ACE2結合,隨后TMPRSS2就會與新冠病毒的S 蛋白作用,將S 蛋白進行切割,進一步促進病毒包膜和宿主細胞膜的融合,使得Omicron的遺傳物質進入宿主細胞內部[9]。TMPRSS2 是一種II型跨膜蛋白酶,在呼吸道和胃腸道、前列腺和其他器官的上皮細胞中廣泛表達,在人體其他器官呈低水平表達[10]。另外,在內體蛋白酶組織蛋白酶L或B 介導的內體膜融合進入途徑中,Omicron 包膜上的S 蛋白與宿主細胞膜上的ACE2 受體接觸結合后,不會被TMPRSS2 切割,而是直接由ACE2 受體介導病毒顆粒通過胞吞作用進入宿主細胞內部,再由宿主細胞內部的組織蛋白酶L 進行切割,釋放出膜融合肽使病毒包膜與囊泡融合,最后使得Omicron的遺傳物質進入宿主細胞內部[11]。
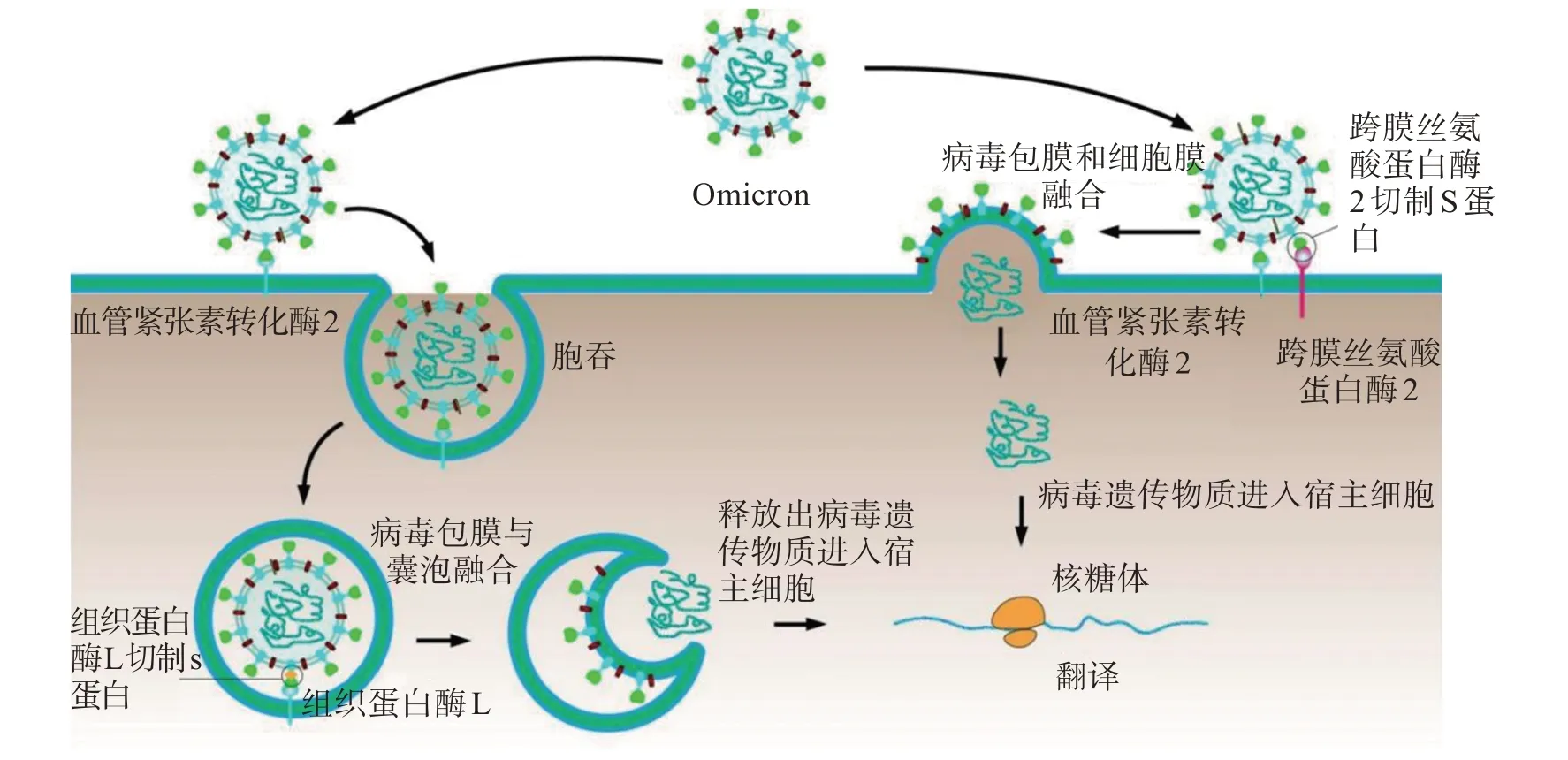
圖1 Omicron侵入人體細胞的機制Figure 1 Mechanism of Omicron invasion into human cells
Omicron 相比于其他變異株具有較低的TMPRSS2 依賴性,也就是說即使在細胞低表達或不表達TMPRSS2 時候,Omicron 也可以入侵宿主細胞。且Omicron 的感染不會被TMPRSS2 抑制劑(Camstat 或Nafamostat)阻斷,這說明Omicron 主要通過內體蛋白酶組織蛋白酶L 或B 介導的內體膜融合進入途徑侵入宿主細胞[12]。還有研究表明,新冠病毒還可以通過與脫唾液酸糖蛋白受體1(asialoglycoprotein receptor 1,ASGR1)和含環(huán)狀結構跨膜蛋白1(kringle containing transmembrane protein 1,KREMEN1)相互作用來侵入宿主細胞[13],也就是說Omicron 也可以利用ASGR1 和KREMEN1 來感染人體細胞,但是目前對于該途徑感染機制尚不明確。
1.3 Omicron對人體的危害
自Omicron 在南非被發(fā)現(xiàn)后不久,就有研究團隊對感染Omicron 患者病癥嚴重程度進行了統(tǒng)計,結果發(fā)現(xiàn)感染Omicron患者的住院率和死亡率均低于新冠病毒原始毒株和其他變異株[14]。還有研究表明,TMPRSS2 在人肺細胞上廣泛表達,但是由于Omicron 對TMPRSS2 依賴性降低,且主要通過內膜融合方式侵入人體細胞,這使得Omicron 對人肺部損害有所降低[15]。這說明Omicron 在突變后,引起嚴重肺炎致人死亡的致病力有所下降。但是由于Omicron 通過ACE2 介導來入侵宿主細胞[11],并且ACE2 在人的多種器官組織細胞上均有表達,所以Omicron 可以同時入侵人體多個器官組織,同時損害人體多個器官組織的功能,表明Omicron 對于人仍具有不可忽視危害性。
有研究表明,當Omicron 入侵人體神經(jīng)細胞時候,會損害人體正常神經(jīng)系統(tǒng)的功能,引起一系列神經(jīng)功能障礙。免疫力較強的成年人被感染時,會出現(xiàn)乏力、頭痛、味覺和嗅覺減退或消失等神經(jīng)功能障礙。免疫不完全或者較弱的兒童被感染時,有可能會出現(xiàn)更為嚴重的后果。有研究表明,兒童感染Omicron后,病毒侵犯兒童神經(jīng)細胞,可能導致患病兒童出現(xiàn)癲癇和驚厥的癥狀[16]。
當Omicron 侵害人體消化系統(tǒng)時,感染者會出現(xiàn)嘔吐、腹瀉等腸胃炎癥狀,嚴重情況可導致脫水甚至休克。還有研究表明,Omicron 感染會加重患者患有其他疾病的癥狀,如高血壓、糖尿病、腦血管疾病、心血管疾病、呼吸道疾病、惡性腫瘤、慢性腎病和慢性肝病[17]。還有研究發(fā)現(xiàn),Omicron 感染會損傷男性的生殖系統(tǒng),由于Omicron 與ACE2 的結合[18]增強和TMPRSS2 依賴性降低,使得Omicron 可以更輕易感染一些表達ACE2 而低表達TMPRSS2的組織器官,例如男性的睪丸[19]。
Omicron 在突變后具有了更強的逃逸能力。曾有研究團隊測試,對Omicron 使用了11 種抗新型冠狀病毒的單克隆抗體進行中和實驗,結果發(fā)現(xiàn)這11種抗體中有6 種抗體對Omicron 失去了中和能力,4種抗體對Omicron 的中和效果降低,只有1 種仍具有穩(wěn)定的中和效力[20]。這提示,即使是目前注射過三針新冠病毒疫苗的接種者也可能被Omicron感染。
此外,Omicron 的S 蛋白發(fā)生了37 個突變,這使得Omicron 可能具有更強的感染能力和隱匿能力。通過小鼠感染模型實驗研究,發(fā)現(xiàn)這些突變使得S蛋白與宿主細胞ACE2 的親和性增強,使Omicron具有更強的感染能力[21]。同時還有研究發(fā)現(xiàn),Omicron 的S 蛋白活性略低于其他變異株。當采用表達最低水平內源性ACE2 的HEK293 細胞作為靶細胞時,所有其他變體(特別是Delta)都表現(xiàn)出顯著的融合活性,而Omicron 的S 蛋白卻依舊不活躍,Omicron 的S 蛋白在10 倍高的ACE2 水平時才表現(xiàn)出類似的融合活性[22]。這說明,Omicron 在感染初期時檢測容易出現(xiàn)假陰性,使得Omicron 在感染宿主后隱匿性獲得加強。
2 人體T、B、NK細胞的免疫反應
2.1 人體T細胞免疫反應
T 淋巴細胞是人體對抗病毒入侵中發(fā)揮細胞免疫的關鍵細胞(見表1)。當Omicron 入侵到人體內部時候,初始T細胞就會在樹突狀細胞(DC細胞)的抗原提呈作用或細胞因子作用下激活、分化為CD4+T 細胞和CD8+T 細胞。其中CD4+T 細胞在Omicron 的S 蛋白抗原刺激下會釋放出大量的細胞因子,如白細胞介素2(InterleuKin-2,IL-2),γ干擾素(Interferon-γ,IFN-γ)和白細胞介素4(InterleuKin-4,IL-4),進一步促進人體對新冠病毒的抗病毒免疫反應[23],過量的細胞因子會導致細胞因子風暴發(fā)生。
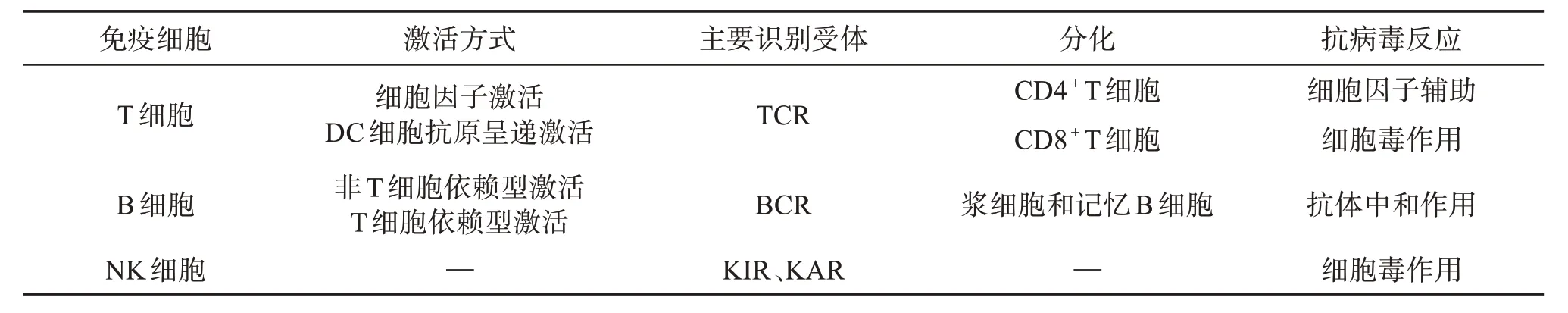
表1 T、B、NK細胞抗病毒免疫的對比Table 1 Comparison of antiviral immunity among T cells,B cells and NK cells
CD8+T 細胞在Omicron 入侵后在CD4+T 細胞產(chǎn)生的IL-2 的激活下,通過自身的T 細胞抗原受體(T cell receptor,TCR)交叉識別Omicron 感染宿主細胞后殘留在宿主細胞膜上的表面抗原,如S蛋白、E 蛋白和M 蛋白抗原[24],釋放穿孔素和顆粒酶或通過CD40/CD40 L途徑發(fā)揮細胞毒性作用。
2.2 人體B細胞免疫反應
B淋巴細胞是人體適應性抗病毒免疫中發(fā)揮體液免疫功能的重要角色(見表1)。當Omicron 入侵人體后或注射新冠疫苗后,B 淋巴細胞通過自身的B 細胞抗原受體(B-cell receptor,BCR)對Omicron相關抗原,如Omicron 核酸抗原或S 蛋白抗原進行識別[25],前者會使B 細胞發(fā)生非T 細胞依賴型激活使B 細胞產(chǎn)生IgM 抗體;后者使B 細胞發(fā)生T 細胞依賴型激活,分化成漿細胞和記憶B細胞,漿細胞會產(chǎn)生針對S蛋白抗原特異性的IgG抗體[26]。
這些Omicron 特異性IgM 抗體和IgG 抗體通過和新冠病毒上的S蛋白抗原或被感染細胞膜表面殘留的S 蛋白抗原結合后,介導后續(xù)其他免疫細胞或免疫分子參與抗病毒免疫反應,最終清除病毒。其主要作用機理如下:IgM、IgG 抗體與補體C1 的C1 q結合,啟動補體依賴性細胞毒作用(complement dependent cytotoxicity,CDC)[27],IgG 抗體可以通過Fc-FcγR 相互作用引發(fā)抗體依賴性的細胞毒作用(antibody-dependent cytotoxicity,ADCC)或者抗體依賴性細胞介導的吞噬作用(antibody-dependent cellular phagocytosis,ADCP)[28]。 CDC、ADCC、ADCP可以有效清理人體被感染的細胞和游離的新冠病毒顆粒。
2.3 人體NK細胞免疫反應
NK 細胞是人體固有免疫系統(tǒng)的重要免疫細胞,屬于固有樣淋巴細胞(innate lymphoid cells,ILC);NK 細胞特點是具有非特異性識別異常細胞的能力(見表1)。
在NK 細胞的細胞膜表面存在殺傷細胞活化受體(killer activation receptor,KAR)和殺傷細胞免疫球蛋白樣受體(killer immunoglobulin-like receptors,KIR),正常情況下,它們分別與人體細胞上的主要組織相容性復合體Ⅰ類分子(major histocompatibility complex-Ⅰ,MHC-Ⅰ)和多糖類抗原結合抑制NK細胞發(fā)揮細胞毒作用[29-30]。當Omicron 感染宿主細胞后,宿主細胞膜上的MHC-Ⅰ類分子結構發(fā)生變化或者表達降低,導致KIR 無法正常識別,進而引發(fā)NK 細胞對被感染細胞發(fā)揮細胞毒作用,使被感染細胞凋亡,最終達到消滅入侵人體Omicron 的目的。
NK細胞的細胞毒作用與CD8+T細胞具有一定的相似性,也會分泌穿孔素和顆粒酶,不過NK 細胞還會分泌瘤壞死因子(tumor necrosis factor,TNF)。
3 Omicron免疫逃逸的機制
3.1 Omicron逃逸T細胞攻擊的機制
目前,為了預防新冠病毒的感染,國內很多人都響應國家政府的號召,接種了三針新冠病毒疫苗。通過疫苗刺激自身免疫系統(tǒng),使得機體獲得對新冠病毒特異性免疫能力。Omicron 的S 蛋白基因上有30 個突變[2],這使得其中RBD 段發(fā)生數(shù)種氨基酸的變化。
有研究表明,發(fā)生在Omicron S 蛋白上RBD 的突變,導致大約20%的新冠疫苗接種者體內新冠病毒S 蛋白特異性的CD4+T 細胞和記憶性CD8+T 細胞的免疫反應功能下降約50%[31]。但是有其他研究表明,由于人體T 細胞可以將新型冠狀病毒其他成分作為識別抗原,Omicron 僅通過S 蛋白上的突變無法完全逃脫T 細胞的免疫作用[32]。面對這種情況,Omicron仍有其他逃逸T細胞免疫的方法。
新冠病毒Omicron可以通過引發(fā)人體細胞因子風暴來導致宿主出現(xiàn)免疫失調現(xiàn)象[33]。Omicron 通過嚴重炎癥反應來導致人體產(chǎn)生大量細胞因子,過量細胞因子(如IL-6、IL-10、IL-1β、CXCL8、CXCLS和TNF)引起一部分初始T細胞出現(xiàn)錯誤表達,使后者向著低效能、低活性的方向分化[34]。在這些因子影響下,初始T 細胞會分化為低活化T 細胞和過度活化T 細胞,前者最終會變成晚期衰竭T 細胞無法發(fā)揮正常的抗病毒免疫反應,而后者則會引起患者出現(xiàn)全身抗病毒反應,加重患者病狀[35],最終導致T細胞過度激活,會使人體的CD4+T、CD8+T 細胞數(shù)量顯著減少(圖2)。
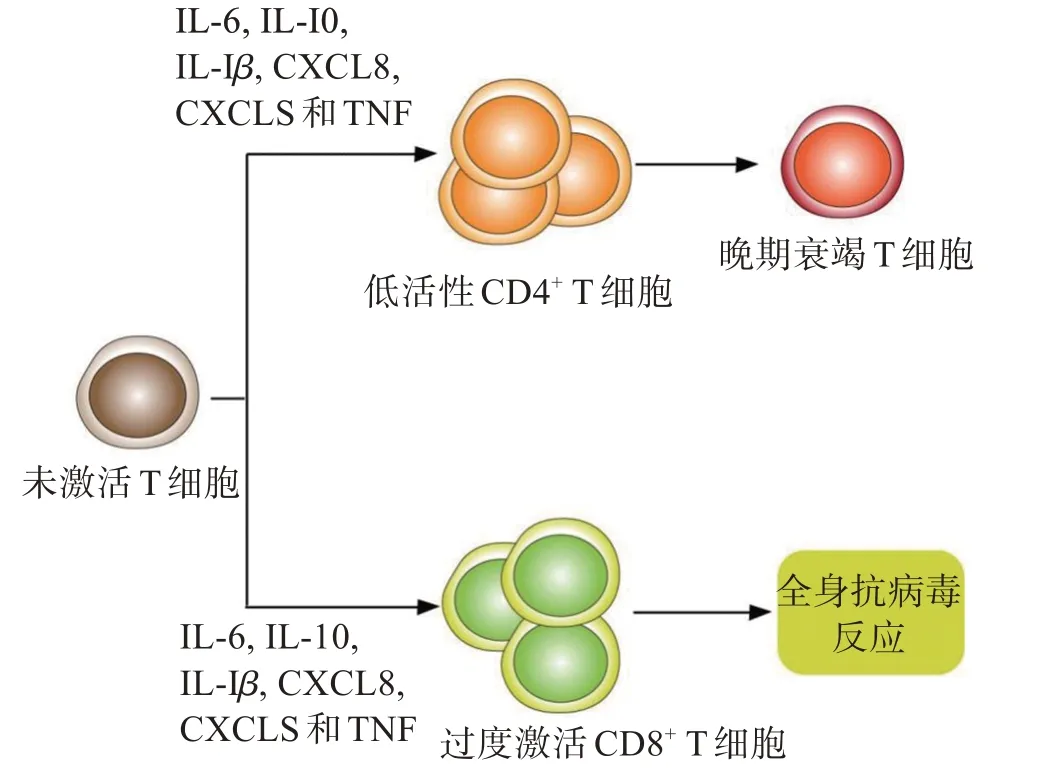
圖2 Omicron引發(fā)T細胞錯誤激活Figure 2 Incorrect activation of T cells triggered by Omicron
3.2 Omicron逃逸B細胞攻擊的機制
Omicron 應對B 細胞策略的主要表現(xiàn)就是抗體中和作用的減弱甚至消失。Omicron 在S 蛋白上的突變可能導致疫苗接種者和新冠肺炎康復者體內產(chǎn)生的抗體對Omicron 的中和作用下降甚至消失。有研究表明,疫苗接種者和有既往感染康復者體內抗體對Omicron 抵抗效力僅為56%[36]。這是由于Omicron S 蛋白上的多個氨基酸發(fā)生突變,在N 端結構域(NTD)和受體結合結構域(RBD)上出現(xiàn)了約20 個氨基酸的替換,使該變異株S 蛋白RBD 和S 蛋白本身的疏水氨基酸增加和局部構象改變,這些變化導致了大部分既有的RBD 特異性抗體和NTD 特異性抗體無法識別Omicron[37](圖3),例如,Omicron S蛋白上存在的K444Q、V445A和P499R/S突變會降低病毒與bebtelovimab 單克隆抗體的結合[38]。雖然Omicron S 蛋白突變會影響大部分特異性抗體與S 蛋白的結合,但是仍有小部分抗體通過結合S 蛋白中保守片段來發(fā)揮對Omicron 的中和作用,例如35B5抗體[39]。
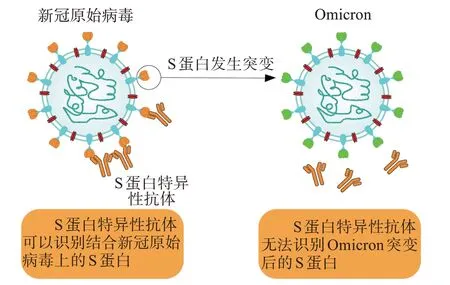
圖3 Omicron S蛋白突變導致抗體無法識別Figure 3 Omicron S protein mutation results in unrecognizable antibodies
3.3 Omicron逃逸NK細胞攻擊的機制
Omicron 逃逸NK 細胞攻擊的機制與T 細胞的有相同之處,也是通過細胞因子風暴影響NK 細胞的歸巢。NK 細胞會在炎癥趨化因子的影響下出現(xiàn)歸巢現(xiàn)象,這原本是人體免疫系統(tǒng)清除異常細胞正常現(xiàn)象(圖4)。
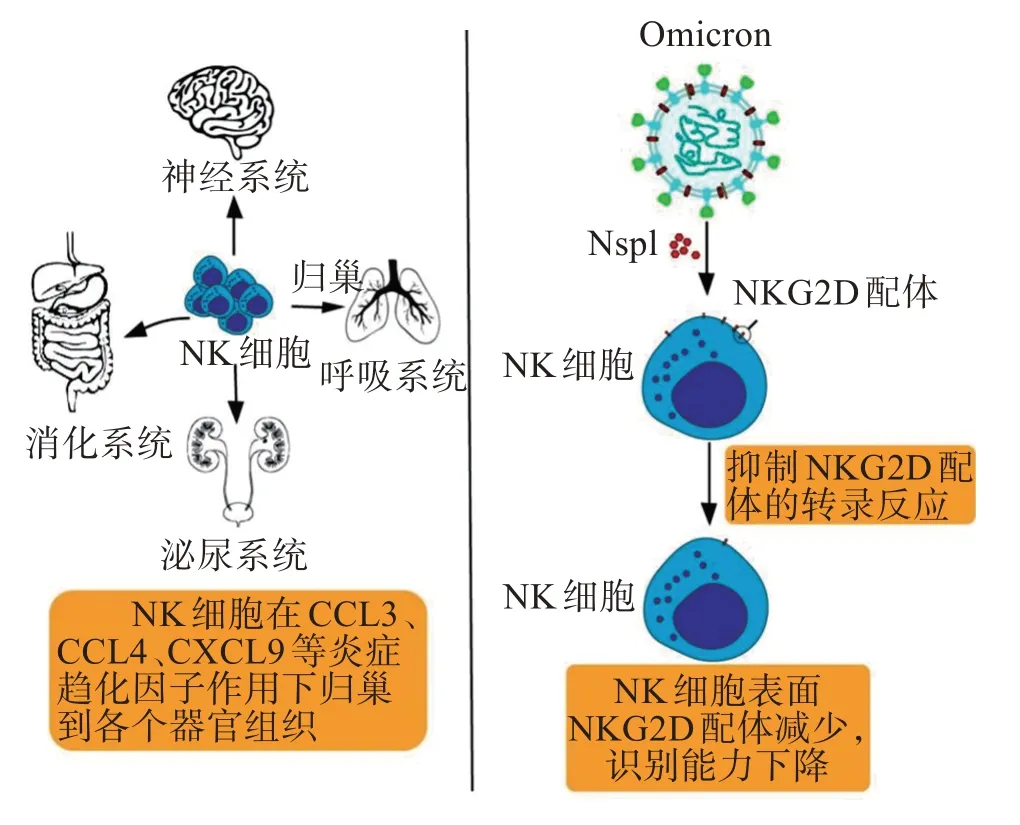
圖4 Omicron對NK細胞的策略Figure 4 Strategy of Omicron for NK cells
在過往研究中,新冠病毒可以引起細胞因子風暴,誘導分泌CCL3、CCL4 和CXCL9 等趨化因子干擾和誤導NK 細胞的歸巢。過度激活的NK 細胞通過CXCR3、CCR5 等趨化因子受體介導,歸巢到人體的肺部,引起人肺部發(fā)生嚴重炎癥反應,損傷肺部細胞[40]。
由于Omicron進入人體細胞主要方式和原始病毒株不同,Omicron 會同時感染人體多個組織器官的細胞。換言之,Omicron對NK細胞歸巢的影響不再主要局限在肺部,而是會出現(xiàn)多個器官組織NK細胞歸巢現(xiàn)象,同時損傷人體多個組織器官。
此外,新冠病毒還能通過非結構蛋白1(non-structural protein 1,Nsp1)來逃脫NK 細胞的殺傷。Nsp1是新冠病毒產(chǎn)生的一種非結構蛋白,研究發(fā)現(xiàn)Nsp1 能通過翻譯抑制和mRNA 降解的方式抑制宿主基因表達[41]。有研究發(fā)現(xiàn),新冠病毒可以通過Nsp1 介導的NKG2 D 配體(NKG2 DL)下調,逃脫了NK 細胞的直接殺傷。NKG2 D 配體是NKG2 D/NKG2 DL 通路的重要功能部分,NKG2 D 配體的表達減少會導致NK 細胞識別功能降低甚至無法識別異常細胞,從而使得新冠病毒感染宿主細胞能夠逃脫NK細胞的細胞毒作用[42]。
4 問題與展望
目前Omicron 仍在變異,WHO 又報道了一種Omicron 亞種XBB.1.16,并且該亞種更具傳染性。Omicron 的病毒結構和其原始毒株相似,但在許多重要蛋白上出現(xiàn)了突變。這使得Omicron可能具有更強的感染能力和逃逸能力。在之前面對新冠病毒時候,人們通過既往感染或接種新冠疫苗獲得了一定免疫能力,但是Omicron 的突變也給予了其突破和逃逸人體免疫的能力。雖然Omicron對肺的致病力下降了,但是由于Omicron 感染方式的變化使得更可能感染危害人體其他重要器官:感染心臟會引發(fā)心肌損傷,感染神經(jīng)細胞會引發(fā)癲癇,感染腸道會引起嚴重腸胃癥狀,感染睪丸甚至會導致生育能力下降。
Omicron 具有極強的突變能力和逃逸能力,這使得目前人們應對Omicron 的策略也需要不斷更新。目前國內上市了4 款新冠藥物:奈瑪特韋片/利托那韋片組合包裝(Paxlovid)、阿茲夫定、莫若拉韋、安巴韋單抗注射液及羅米司韋單抗注射液。阿茲夫定屬于人工合成的核苷類似物,可在細胞內磷酸化,成為有活性的5-三磷酸鹽代謝物,從而起到抑制病毒活性的效果。莫若拉韋可以抑制病毒RNA 復制所需的酶。雖然這些藥物都可以在一定程度上抑制或治療新冠病毒感染,但是它們均屬于廣譜抗病毒藥物,缺乏對Omicron的特異性。
Paxlovid 與安巴韋單抗注射液及羅米司韋單抗注射液是目前已研發(fā)的可針對Omicron 的特效藥。Paxlovid 中的奈瑪特韋成分是抗病毒“主力軍”,其作用是通過阻斷SARS-CoV-2 的主要多聚蛋白酶(SARS-CoV-2 Mpro)合成以抑制新冠病毒蛋白的復制;而Paxlovid 中利托那韋則是抗病毒的“強力助攻”,其最初是用于治療HIV-1 感染的蛋白酶抑制劑,但它同時也是細胞色素P450-3 A4(CYP3 A4)酶的有效抑制劑。在這個組合中,利托那韋的作用主要是抑制奈瑪特韋的代謝,降低奈瑪特韋在體內降解的速度,發(fā)揮更強、更持久的作用[43]。安巴韋單抗注射液及羅米司韋單抗注射液通過生物工程技術從新冠康復患者B淋巴細胞中分離篩選出的中和抗體,經(jīng)工業(yè)化大規(guī)模生產(chǎn),制備成標準化的藥品,這兩種單克隆抗體可精準靶向新冠病毒的刺突蛋白受體結合域(RBD),從而阻止刺突蛋白介導病毒入侵人體細胞,更特異、更高效[44]。換言之,安巴韋單抗注射液及羅米司韋單抗注射液是科研工作者從B細胞抗Omicron的角度被研發(fā)出來的。
新冠病毒目前仍處于不斷變異的過程中,針對新冠變異的情況,需要及時做出應對策略。目前的思路有二:一是特效藥物的聯(lián)用可以更有效地靶向性抗病毒,達到“1+1>2”的效果;二是研制更具特異性治療Omicron 感染和抑制Omicron 免疫逃逸的藥物,根據(jù)目前研究發(fā)現(xiàn),從避免Omicron逃逸NK細胞攻擊的角度來看,科研工作者可以通過抑制Omicron合成非結構蛋白所需酶或者以這些酶類作為靶點研發(fā)治療Omicron的特效藥。
