紅樹林來源細(xì)菌抑菌活性及其增肥潛力評(píng)估
2024-01-01 00:00:00楊道茂周鈺燁李嘉欣易家樂楊勤偉黃康隆




摘要: 通過平板對(duì)峙實(shí)驗(yàn)、菌絲生長(zhǎng)速率抑制實(shí)驗(yàn),測(cè)試細(xì)菌對(duì)禾谷鐮刀菌、假禾谷鐮刀菌、香蕉枯萎病菌、油茶炭疽病菌、巴西曲霉的抑菌活性,并通過固氮解磷實(shí)驗(yàn)、產(chǎn)γ-聚谷氨酸實(shí)驗(yàn)探究增肥潛力。結(jié)果表明:細(xì)菌YY9,暹羅芽孢桿菌T1,T6,貝萊斯芽孢桿菌T3對(duì)菌絲生長(zhǎng)速率抑制率均達(dá)到80%以上;細(xì)菌YY6~YY12表現(xiàn)出較強(qiáng)的固氮能力,細(xì)菌YY9產(chǎn)γ-聚谷氨酸質(zhì)量濃度達(dá)到(6.57±0.34) g·L-1。
關(guān)鍵詞: 貝萊斯芽孢桿菌; 暹羅芽孢桿菌; 抑菌活性; 固氮能力; γ-聚谷氨酸
中圖分類號(hào): S 476文獻(xiàn)標(biāo)志碼: A"" 文章編號(hào): 1000-5013(2024)05-0654-07
Evaluation of Antibacterial Activity and Fertilization Potential of Bacteria Derived From Mangrove Forests
Abstract: The antibacterial activity of bacteria against Fusarium graminearum, Fusarium pesudograminearum, banana wilt fungus, Camellia oleifera, and Aspergillus brasiliensis were tested through plate confrontation experiments and mycelial growth rate inhibition experiments. The fertilization potential was explored through nitrogen fixation and phosphorus removal experiment, as well as γ-polyglutamic acid production experiment. The results show that bacteria YY9, Bacillus siamensis T1 and T6, Bacillus velezensis T3 have inhibition rate" over 80% on mycelial growth rate. Bacteria YY6-YY12 show stronger nitrogen fixation ability, and the mass concentration of γ-polyglutamic acid produced by bacteria YY9 can reach (6.57±0.34) g·L-1.
Keywords: Bacillus velezensis; Bacillus siamensis; antibacterial activity; nitrogen fixation ability; γ-polyglutamic acid
農(nóng)業(yè)生產(chǎn)對(duì)維護(hù)人類糧食安全和社會(huì)穩(wěn)定起到了重要的作用。然而,植物病蟲害的不斷蔓延對(duì)農(nóng)作物產(chǎn)量和質(zhì)量造成了嚴(yán)重威脅,限制了農(nóng)業(yè)的可持續(xù)發(fā)展,如假禾谷鐮刀菌(Fusarium pesudograminearum)、禾谷鐮刀菌(Fusarium graminearum)等病原真菌綜合侵染引起的小麥莖基腐病[1],鐮刀菌或腐霉菌單獨(dú)侵染、鐮刀菌和腐霉菌復(fù)合侵染引起的玉米莖腐病[2],古巴尖鐮孢菌侵染引起的香蕉枯萎病[3], 油茶炭疽病菌(Camellia oleifera)則是危害油茶植株生長(zhǎng)和產(chǎn)量的主要病害[4-5]。
防控植物病害的方法一般采用化學(xué)防控手段,但殺菌劑的長(zhǎng)期使用會(huì)導(dǎo)致環(huán)境污染、生態(tài)失衡及“3R”問題的產(chǎn)生等,而生物防控對(duì)環(huán)境友好,病原菌不易產(chǎn)生抗藥性,符合可持續(xù)發(fā)展需求。因此,開發(fā)綠色微生物農(nóng)藥有現(xiàn)實(shí)意義和經(jīng)濟(jì)意義。
微生物農(nóng)藥因具備選擇性高、對(duì)人畜無害、對(duì)自然環(huán)境污染小、不易產(chǎn)生抗藥性等優(yōu)點(diǎn)而倍受青睞,是目前發(fā)展最迅速、推廣應(yīng)用最成功的一類生物農(nóng)藥產(chǎn)品。與真菌和病毒等其他微生物農(nóng)藥相比,細(xì)菌易于培養(yǎng),農(nóng)藥活性評(píng)價(jià)也相對(duì)容易。更重要的是細(xì)菌發(fā)酵生產(chǎn)工藝成熟,工業(yè)化生產(chǎn)成本可控,更加便于產(chǎn)業(yè)化利用。所以,細(xì)菌類微生物農(nóng)藥一直受到科研單位和企業(yè)的青睞,成為微生物農(nóng)藥研發(fā)創(chuàng)制和商業(yè)化開發(fā)的熱點(diǎn)領(lǐng)域。在2010-2020年全球新登記的微生物農(nóng)藥中,細(xì)菌種類占比超過40%[6]。目前,我國(guó)主要生物農(nóng)藥品種多樣[7],包括細(xì)菌類微生物農(nóng)藥、真菌類微生物農(nóng)藥、病毒類微生物農(nóng)藥、基因工程菌類微生物農(nóng)藥等,在植物保護(hù)、蟲害防治方面發(fā)揮著重要作用。
細(xì)菌不僅可以開發(fā)為生物農(nóng)藥,而且還有增肥效果。如某些細(xì)菌具有合成γ-聚谷氨酸(γ-PGA)的能力。γ-PGA是由多種桿菌產(chǎn)生的一種胞外多肽,具有優(yōu)良生物相容性、生物降解性及無毒無污染性,廣泛用作藥物緩釋材料、食品的水凝劑及高強(qiáng)度纖維[8]。在農(nóng)業(yè)應(yīng)用方面,γ-PGA可以用于改良酸化植煙土壤[9],調(diào)節(jié)土壤微生物群落變化和促進(jìn)作物生長(zhǎng),富集有益微生物,在作物采摘后依舊維持較高的土壤生物活性[10],與化學(xué)肥料合用,起到明顯提高肥效的作用[11]。目前,暹羅芽孢桿菌[12-13]、地衣芽孢桿菌[14]、枯草芽孢桿菌[11,15]等細(xì)菌均具有產(chǎn)γ-PGA的能力。基于此,本文對(duì)紅樹林來源細(xì)菌抑菌活性及增肥潛力進(jìn)行評(píng)估。
1 材料與方法
1.1 藥品與儀器
γ-PGA(上海市麥克林生化科技股份有限公司);葡萄糖,磷酸氫二鈉,硫酸鎂等藥品(國(guó)產(chǎn)分析純);TGL-20M型臺(tái)式高速冷凍離心機(jī)(湖南省長(zhǎng)沙市湘儀實(shí)驗(yàn)室儀器開發(fā)有限公司);UV-1800PC型紫外可見分光光度計(jì)(上海市美譜達(dá)儀器有限公司)。
1.2 供試試驗(yàn)菌株
油茶炭疽病菌由華僑大學(xué)化工學(xué)院王奇志副教授饋贈(zèng)。香蕉枯萎病菌(banana wilt fungus)由華僑大學(xué)化工學(xué)院王明元教授饋贈(zèng)。禾谷鐮刀菌,假禾谷鐮刀菌由江蘇省南京市中旗科技股份有限公司饋贈(zèng)。巴西曲霉(Aspergillus brasiliensis)購自廣東省廣州市微生物菌種保藏中心。
1.3 培養(yǎng)基配方
培養(yǎng)基配方,如表1所示。
2 實(shí)驗(yàn)部分
2.1 細(xì)菌的分離
用無菌藥勺取約5 g福建省廈門市集美大橋紅樹林根部土壤,并將其裝于無菌培養(yǎng)皿中。將約1 g土壤倒入9 mL無菌水試管中,搖勻后,稀釋10倍。取0.5 mL稀釋液涂布于馬鈴薯葡萄糖瓊脂培養(yǎng)基(PDA)上,30 ℃培養(yǎng)2~5 d。觀察微生物生長(zhǎng)情況,依據(jù)菌落特征的差異挑取菌株。純化菌株,直到獲得單一菌落為止。挑選一部分菌株送往北京市擎科生物科技股份有限公司做16S rDNA鑒定。
2.2 固氮解磷效果
細(xì)菌固氮解磷的實(shí)驗(yàn)流程參考文獻(xiàn)[16],并做略微調(diào)整。挑取一接種環(huán)培養(yǎng)物分別接種到阿須貝無氮培養(yǎng)基、蒙金娜無機(jī)磷培養(yǎng)基和蒙金娜有機(jī)磷培養(yǎng)基上,每個(gè)培養(yǎng)基接種3~4株菌,培養(yǎng)5 d后觀察透明圈產(chǎn)生情況。
2.3 對(duì)病原真菌的拮抗活性
采用對(duì)峙培養(yǎng)法考察各細(xì)菌對(duì)病原真菌的拮抗活性[18]。在PDA中央位置接種病原菌菌絲,并在周邊等距接3~4株細(xì)菌,在室溫下培養(yǎng)5 d后觀察其拮抗效果。
2.4 對(duì)病原真菌菌絲體生長(zhǎng)速率的抑制效果
實(shí)驗(yàn)過程參考文獻(xiàn)[16],并做一些調(diào)整。取節(jié)2.3具有拮抗效果的菌株接種于裝有30 mL的LB液體培養(yǎng)基中,于30 ℃培養(yǎng)2 d后,取2 mL菌液加入到無菌培養(yǎng)皿中,隨后倒入約50 ℃,20 mL的PDA中,搖勻。待培養(yǎng)基凝固后,往培養(yǎng)基中央接種一環(huán)病原真菌菌絲,室溫下培養(yǎng)7 d,以不接細(xì)菌的PDA為對(duì)照。生長(zhǎng)結(jié)束后分別測(cè)量對(duì)照組菌絲直徑、實(shí)驗(yàn)組菌絲直徑及菌絲生長(zhǎng)速率抑菌率。菌絲生長(zhǎng)速率抑制率計(jì)算式為
2.5 γ-PGA產(chǎn)量
采用比濁法建立γ-PGA標(biāo)準(zhǔn)曲線。精確配制γ-PGA母液,γ-PGA質(zhì)量濃度為0.500 g·L-1,然后逐步稀釋成0.278,0.227,0.182,0.167,0.154,0.125,0.100 g·L-1溶液,備用。分別取3 mL各質(zhì)量濃度標(biāo)準(zhǔn)品溶液與等體積十六烷基三甲基溴化銨(CTAB,0.07 mol·L-1)溶液混勻,室溫下靜置3 min后,測(cè)量其在400 nm波長(zhǎng)下的吸光度(D(400))[19]。將各細(xì)菌接種于γ-PGA發(fā)酵培養(yǎng)基中,30 ℃培養(yǎng)3 d后,發(fā)酵液在轉(zhuǎn)速為10 000 r·min-1離心機(jī)中離心20 min,將1 mL上清液稀釋10,20倍,最終按照標(biāo)準(zhǔn)曲線計(jì)算γ-PGA質(zhì)量濃度。每株菌重復(fù)3次,最后計(jì)算平均值。
2.6 復(fù)合菌劑可能性
考察增肥潛力細(xì)菌與抑菌活性的芽孢桿菌進(jìn)行復(fù)配的可能性。將具有增肥潛力的細(xì)菌YY6,YY11接種于50 mL的LB培養(yǎng)基中,30 ℃培養(yǎng)24 h后,各取0.5 mL溶液涂布于PDA培養(yǎng)基上,隨后在培養(yǎng)基表面接種暹羅芽孢桿菌T1,T6(細(xì)菌T1,T6),貝萊斯芽孢桿菌T3(細(xì)菌T3),細(xì)菌YY9,30 ℃培養(yǎng)2 d后觀察是否有透明圈,不產(chǎn)生透明圈,則表明二者可共培養(yǎng),有制備復(fù)合菌劑的可能。
3 實(shí)驗(yàn)結(jié)果與討論
3.1 菌株分離
從紅樹林根部土壤中共分離到130多株細(xì)菌。取其中3株細(xì)菌T1,T3,T6進(jìn)行16S rDNA鑒定。結(jié)果表明,細(xì)菌T1,T6為暹羅芽孢桿菌 (相似性分別為99.93%和100.00%),細(xì)菌T3為貝萊斯芽孢桿菌(相似性為100%)。
暹羅芽孢桿菌于2010年被刊物International Journal of Systematic and Evolutionary Microbiology收錄為有效種名,屬芽孢桿菌科芽孢桿菌屬。研究表明,暹羅芽孢桿菌可用于治療煙草赤星病[20-21]、花生白絹病[22],分解原油,降解土壤中化合物等,有著廣泛的應(yīng)用前景[23]。
貝萊斯芽孢桿菌對(duì)辣椒褐腐病[24]、油菜根腫病和稻瘟病[25]、禾谷鐮刀菌[26]均表現(xiàn)出良好的抑菌效果。目前僅有四川省成都市百事東旺生物科技有限公司登記的貝萊斯芽孢桿菌為生物農(nóng)藥(http:∥www.icama.org.cn/),貝萊斯芽孢桿菌還有較大的開發(fā)空間。
3.2 菌株固氮解磷效果
細(xì)菌YY5~YY12固氮解磷效果,如圖1所示。
由圖1可知:細(xì)菌YY6~YY12的透明圈直徑與菌落直徑的比值為2.0~5.2,具有較強(qiáng)的固氮效果;細(xì)菌YY7,YY11具有其他菌株不具備的解無機(jī)磷和有機(jī)磷效果,具有較強(qiáng)的增肥潛力研究?jī)r(jià)值。
3.3 對(duì)病原真菌拮抗活性的考察
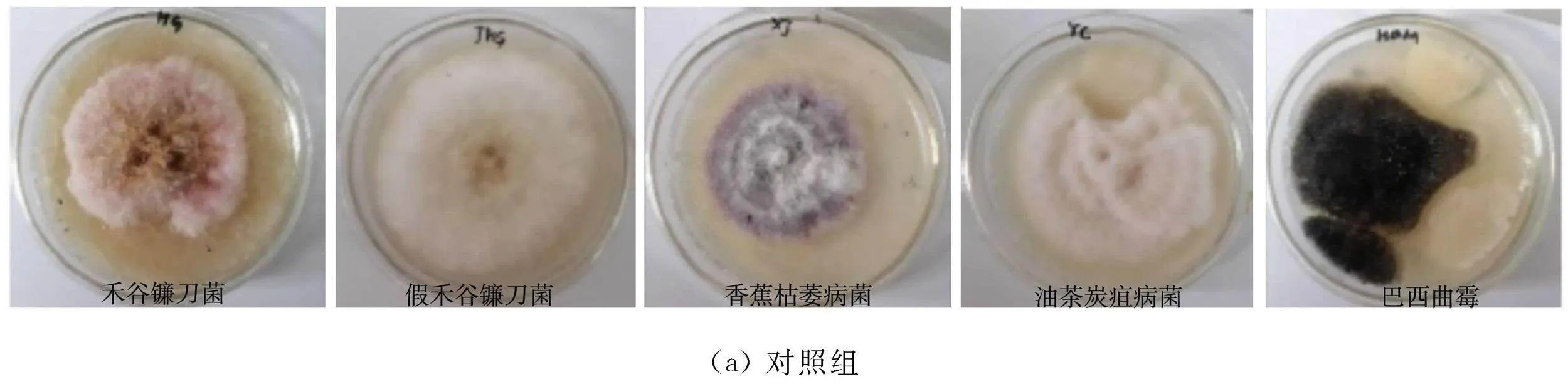
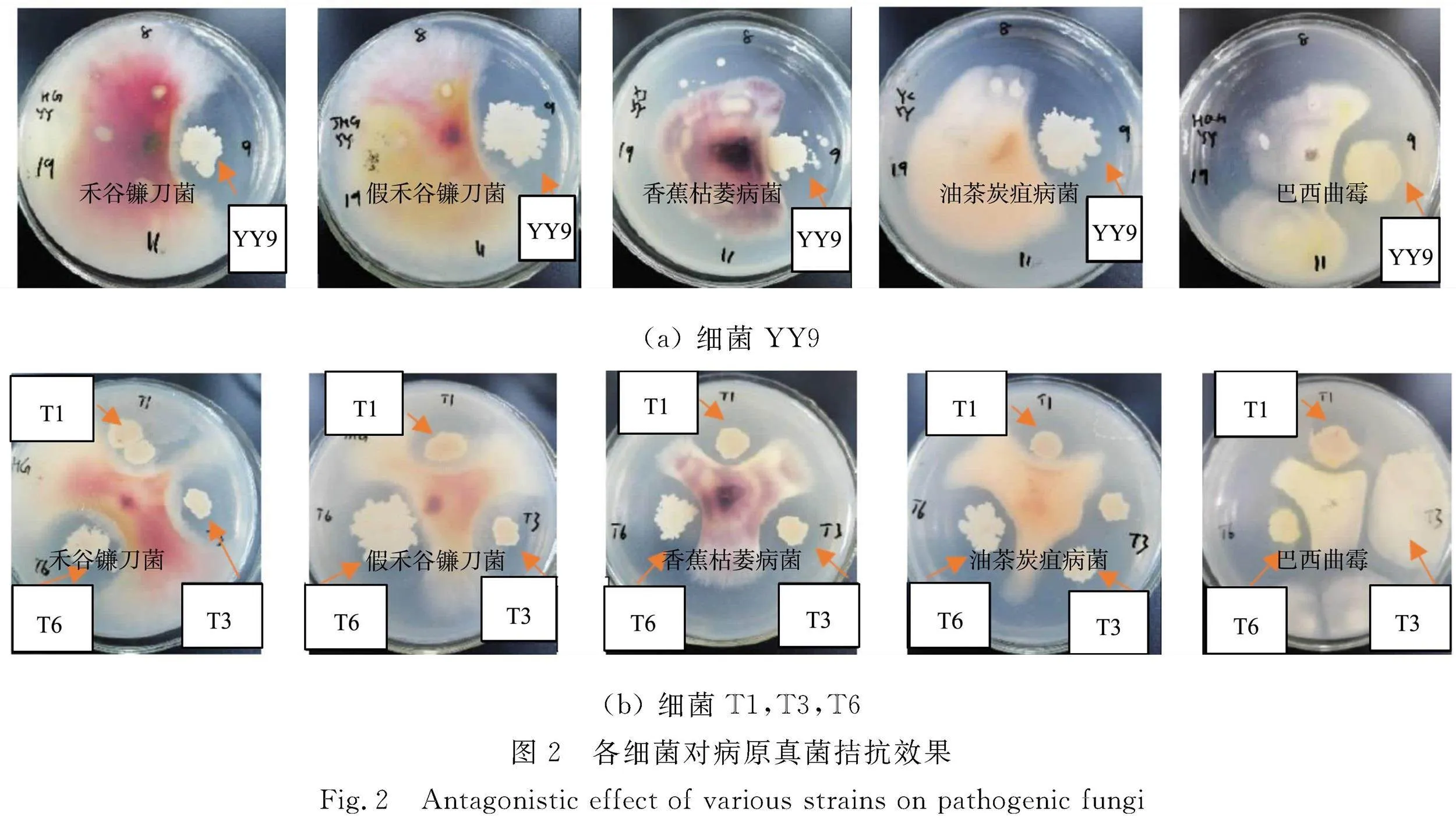
各細(xì)菌對(duì)病原真菌拮抗效果,如圖2所示。由圖2可知:各菌株對(duì)5種植物病原菌表現(xiàn)出較明顯的抑菌效果,有進(jìn)一步研究開發(fā)的價(jià)值。
3.4 植物病原真菌菌絲生長(zhǎng)速率抑制實(shí)驗(yàn)
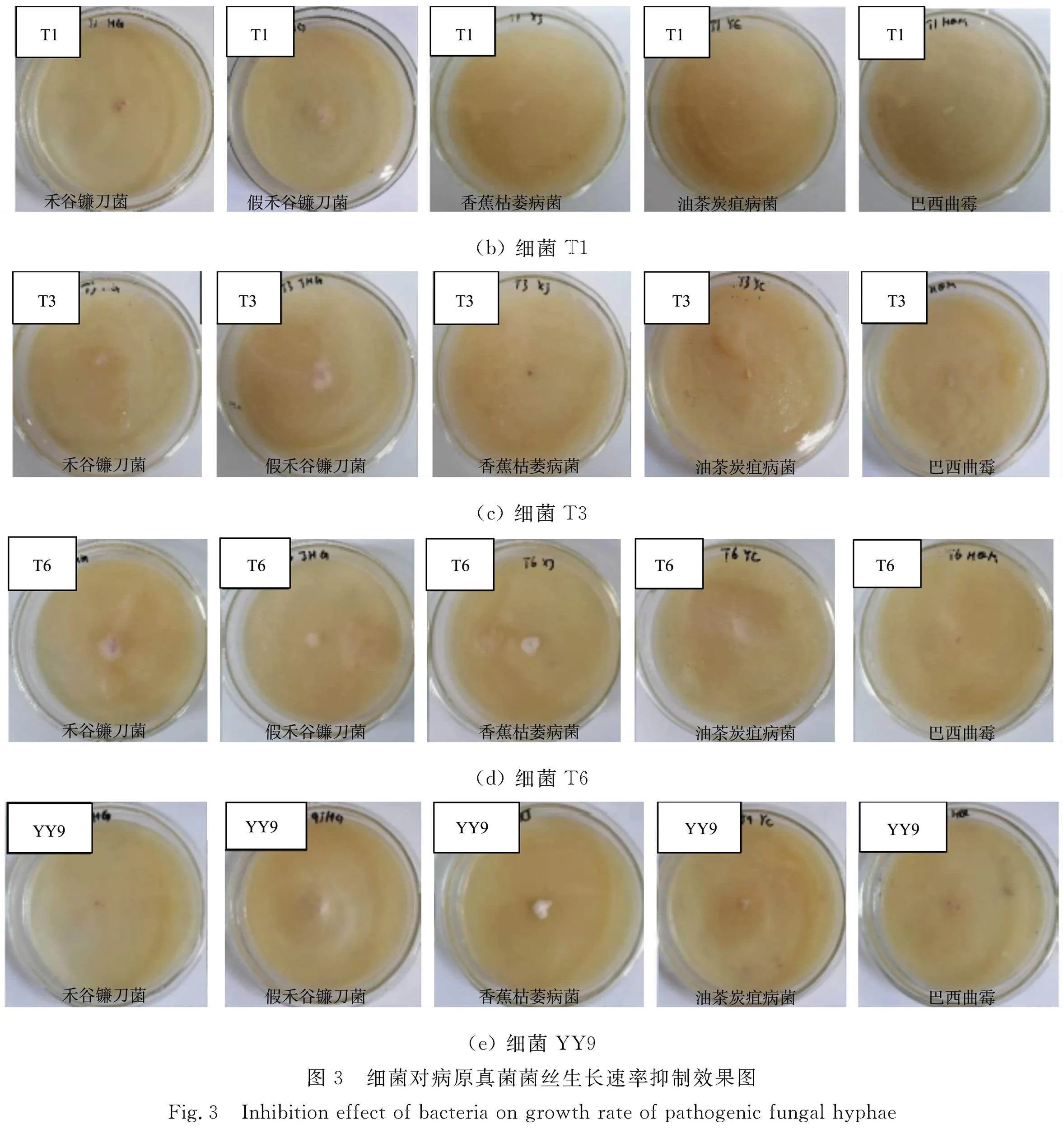
細(xì)菌對(duì)病原真菌菌絲生長(zhǎng)速率抑制效果圖,如圖3所示。
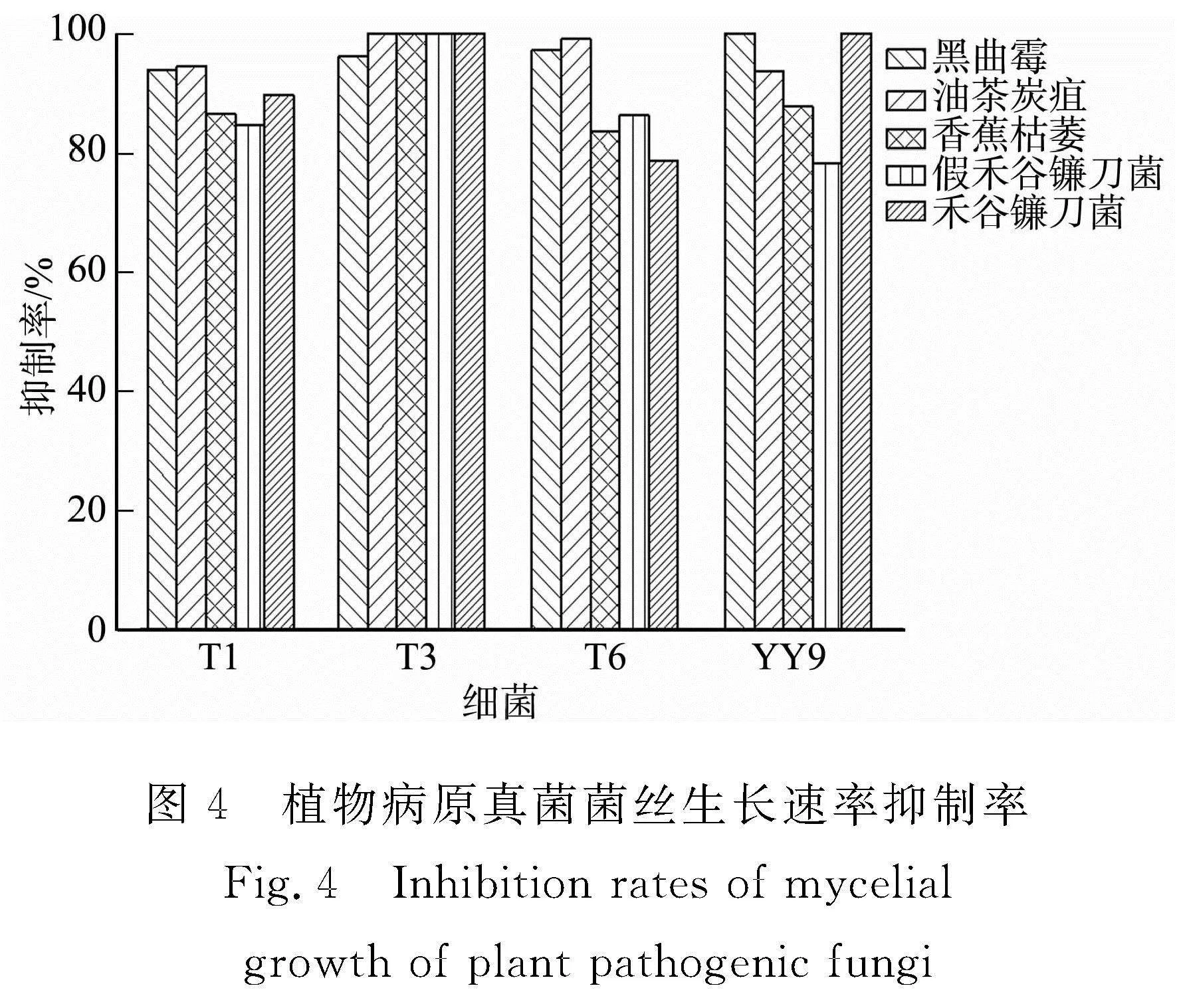
植物病原真菌菌絲生長(zhǎng)速率抑制率,如圖4所示。
由圖3,4可知:4株細(xì)菌對(duì)5種植物病原真菌菌絲生長(zhǎng)速率抑制率均在80%以上,其中細(xì)菌T3的抑制效果最佳,對(duì)5株植物病原菌菌絲生長(zhǎng)速率抑制率均在90%以上。
菌株zk1的菌液對(duì)膠孢炭疽菌的抑制率達(dá)到91.63%,對(duì)哈茨木霉的抑制率也接近80%[27]。細(xì)菌Vel-HNGD-F2抑菌物質(zhì)粗提物對(duì)禾谷鐮刀菌抑菌率達(dá)到(63.21±0.94)%[26]。細(xì)菌Pm9對(duì)禾谷鐮刀菌、小麥全蝕病菌、小麥紋枯病菌、君子蘭莖基腐病菌、番茄灰霉病菌和南天竹炭疽病菌6 種供試植物病原菌抑制率在55.5%~87.5%之間[28]。細(xì)菌T3對(duì)植物病原菌的抑制率與文獻(xiàn)報(bào)道的一致,在農(nóng)業(yè)生產(chǎn)上具有較好的應(yīng)用價(jià)值。
3.5 產(chǎn)γ-PGA的能力
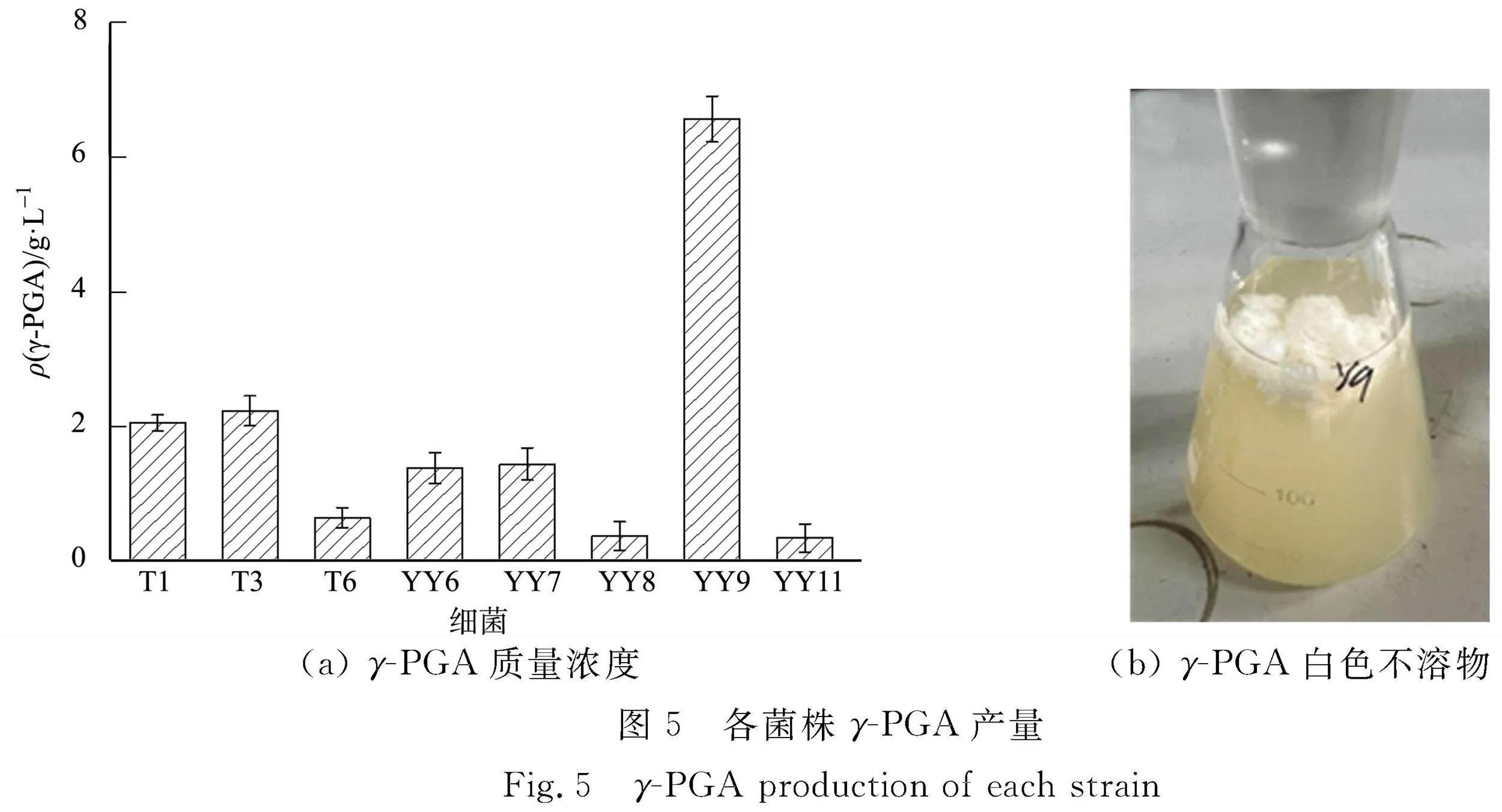
γ-PGA標(biāo)準(zhǔn)曲線為Y=3.355 74X-0.020 95,R2=0.995 3,其中Y為γ-PGA質(zhì)量濃度,X為D(400)。各菌株γ-PGA產(chǎn)量,如圖5所示。圖5中:ρ(γ-PGA)為γ-PGA質(zhì)量濃度。
由圖5可知:8株菌具有合成γ-PGA的產(chǎn)量,其中,細(xì)菌YY9合成的γ-PGA質(zhì)量濃度達(dá)到(6.57±0.34) g·L-1。 細(xì)菌CAU83的γ-PGA最高產(chǎn)量為30.10 g·L-1[29],細(xì)菌LBY-7的γ-PGA最高產(chǎn)量為23.15 g·L-1[13],細(xì)菌LW-1的γ-PGA最高產(chǎn)量為44.78 g·L-1[12]。因此,細(xì)菌YY9的γ-GPA產(chǎn)量還有很大的提升空間。
3.6 復(fù)合菌肥配制可能性評(píng)估
細(xì)菌與芽孢桿菌拮抗關(guān)系,如圖6所示。
由圖6(a)可知:細(xì)菌YY6與4株生防細(xì)菌有一定的拮抗關(guān)系,不能用于復(fù)配復(fù)合菌劑用,而細(xì)菌YY11可與細(xì)菌YY9進(jìn)行復(fù)配。
4 結(jié)論
1) 分離到的細(xì)菌具有固氮功能,其透明圈直徑與菌落直徑的比值在2.0~5.2,具有較強(qiáng)的固氮效果,對(duì)無機(jī)磷和有機(jī)磷的溶解效果則較弱。
2) 通過對(duì)峙培養(yǎng)實(shí)驗(yàn)表明,分離到的細(xì)菌T1,T6,T3,細(xì)菌YY9對(duì)5株植物病原真菌(禾谷鐮刀菌、假禾谷鐮刀菌、香蕉枯萎病菌,油茶炭疽病菌、巴西曲霉)有抑制作用。通過菌絲體生長(zhǎng)速率抑制實(shí)驗(yàn),結(jié)果表明,4株細(xì)菌對(duì)5株植物病原真菌菌絲生長(zhǎng)速率抑制率達(dá)到80%以上,尤其細(xì)菌T3的抑制率達(dá)到90%以上,有進(jìn)一步開發(fā)的價(jià)值。
3) 多株細(xì)菌具有合成γ-PGA的能力,細(xì)菌YY9合成γ-PGA的質(zhì)量濃度為(6.57±0.34) g·L-1,還有很大的提升空間。參考文獻(xiàn):
[1] 吳振鋒,王陸軍,任淑芳,等.苯醚甲環(huán)唑及其復(fù)配劑對(duì)小麥莖基腐病的防效[J].中國(guó)植保導(dǎo)刊,2023,43(9):88-90.
[2] 楊冰娟,陶睿澤,林麗,等.芽孢桿菌抑制玉米莖腐病菌禾谷鐮孢菌和擬輪枝鐮孢菌的研究進(jìn)展[J].江蘇農(nóng)業(yè)科學(xué),2023,51(12):42-49.DOI:10.15889 /j.issn.1002-1302.2023.12.005.
[3] 韋錫昌,林藍(lán)慧,蒙姣榮,等.一株香蕉枯萎病菌拮抗菌的篩選鑒定及盆栽防治效果[J].西南農(nóng)業(yè)學(xué)報(bào),2024,37(7):1522-1530.DOI:10.16213/j.cnki.scjas.2024.7.014.
[4] 李勇,劉飛,葉鍇,等.油茶炭疽病病原菌的分離與鑒定[J].分子植物育種,2024,22(12):3947-3953.DOI:10.13271/j.mpb.022.003947.
[5] 陳潔宇,姜毅,龔涵,等.油茶炭疽病病原鑒定及生物學(xué)特性[J].生物災(zāi)害科學(xué).2024,47(2):224-237.DOI:10.3969/j.issn.2095-3704.2024.02.29.
[6] 張正煒,成瑋,何壯,等.我國(guó)細(xì)菌類微生物農(nóng)藥的登記情況與應(yīng)用策略淺析[J].世界農(nóng)藥,2024,46(4):11-18.DOI:10.16201/j.cnki.cn10-1660/tq.2024.04.02.
[7] 朱偉冉,劉媛,趙晨,等.我國(guó)微生物農(nóng)藥的概述與進(jìn)展[J].工業(yè)微生物,2024,54(3):153-155.DOI:10.3969/j.issn.1001-6678.2024.03.042
[8] 凡朱,呂忠良,楊葉東,等.高黏發(fā)酵液中γ-聚谷氨酸的分離純化工藝[J].化學(xué)工程,2013,41(12):9-11,47.DOI:10.3969 /j.issn.1005-9954.2013.12.003.
[9] 施河麗,彭五星,向修志,等.γ-聚谷氨酸高產(chǎn)菌株的篩選及改良酸化植煙土壤效果研究[J].中國(guó)煙草科學(xué),2022,43(4):15-21.DOI:10.13496/j.issn.1007-5119.2022.04.003.
[10] 陶龍錦,張經(jīng)博,董正武,等.γ-聚谷氨酸對(duì)棉花生長(zhǎng)與根際微生物的影響[J].山西農(nóng)業(yè)大學(xué)學(xué)報(bào)(自然科學(xué)版),2023,43(4):33-43.DOI:10.13842/j.cnki.issn1671-8151.202305001.
[11] 禚優(yōu)優(yōu),邢芳芳,高明夫,等.γ-聚谷氨酸合成菌的鑒定及其對(duì)肥效的影響[J].湖北農(nóng)業(yè)科學(xué),2016,55(17):4569-4572.DOI:10.14088/j.cnki.issn0439-8114.2016.17.052.
[12] 蔡亞慧,王青,王文玉,等.暹羅芽孢桿菌LW-1產(chǎn)γ-聚谷氨酸發(fā)酵培養(yǎng)基的優(yōu)化[J].食品工業(yè)科技,2021,42(16):163-170.DOI:10.13386/j.issn1002-0306.2020110269.
[13] 張雷,張蕾,王玲莉,等.γ-聚谷氨酸生產(chǎn)菌株的鑒定及發(fā)酵培養(yǎng)基優(yōu)化[J].食品工業(yè)科技,2020,41(20):64-71.DOI:10.13386/j.issn1002-0306.2020.20.011.
[14] 李松.農(nóng)用聚谷氨酸低成本液體發(fā)酵工藝研發(fā)[D].濟(jì)南:齊魯工業(yè)大學(xué),2018.
[15] 賈艷萍,殷愛鳴,孫艷梅,等.產(chǎn)聚谷氨酸菌株的篩選及菌株發(fā)酵液對(duì)玉米幼苗抗旱性的作用[J].生物技術(shù)通報(bào),2017,33(10):135-142.DOI:10.13560/j.cnki.biotech.bull.1985.2017-0699.
[16] 梁倩文.禾谷鐮孢生防細(xì)菌的篩選、鑒定及生防潛能研究[D].合肥:安徽農(nóng)業(yè)大學(xué),2023.
[17] 張?chǎng)瑒⒗瑁瑥埰G,等.一株 γ-PGA 生產(chǎn)菌株的分離鑒定及產(chǎn)物性能研究[J].現(xiàn)代食品科技,2014,30(7):44-50.DOI:10.13982/j.mfst.1673-9078.2014.07.001.
[18] 李身.生防細(xì)菌 QTH8 的鑒定及對(duì)小麥莖基腐病、大豆胞囊線蟲病的防治研究[D].洛陽:河南科技大學(xué),2022.
[19] ASHIUCHI M.Analytical approaches to poly-γ-glutamate: Quantification,molecular size determination,and stereochemistry investigation[J].Journal of Chromatography B,2011,879(29):3096-3101.DOI:10.1016/j.jchromb.2011.03.029.
[20] 王東坤.暹羅芽孢桿菌LZ88抗煙草赤星病菌物質(zhì)鑒定與作用研究[D].北京:中國(guó)農(nóng)業(yè)科學(xué)院,2022.
[21] 謝中玉.暹羅芽孢桿菌LZ88對(duì)煙草赤星病的防治效果和作用機(jī)理研究[D].重慶:西南大學(xué),2020.
[22] 張霞,許曼琳,郭志青,等.暹羅芽孢桿菌ZHX-10的分離鑒定及其對(duì)花生白絹病的生防效果[J].中國(guó)油料作物學(xué)報(bào),2020,42(4):674-680.DOI:10.19802/j.issn.1007-9084.2019207.
[23] 林志楷,林文珍.暹羅芽孢桿菌研究進(jìn)展[J].亞熱帶植物科學(xué).2019,48(4):391-396.DOI:10.3969/j.issn.1009-7791.2019.04.016.
[24] 趙春燕,安良聰,趙秋玲,等.貝萊斯芽孢桿菌 JK-1可濕性粉劑研制及對(duì)辣椒褐腐病的防治效果[J].天津農(nóng)業(yè)科學(xué),2023,29(2):52-56,63.
[25] 周旭峰.貝萊斯芽孢桿菌F85劑型研制及防病效果評(píng)估[D].武漢:華中農(nóng)業(yè)大學(xué),2023.
[26] 王沖,李倩,肖紅英,等.貝萊斯芽孢桿菌 Vel-HNGD-F2 產(chǎn)抗菌物質(zhì)發(fā)酵條件優(yōu)化及抗菌特性研究[J].河南工業(yè)大學(xué)學(xué)報(bào)(自然科學(xué)版),2024,45(1):73-80.DOI:10.16433/ j.1673-2383.2024.01.010.
[27] 陳少先.貝萊斯芽孢桿菌zk1脂肽類次級(jí)代謝產(chǎn)物的抑菌機(jī)理研究[D].廣州:仲愷農(nóng)業(yè)工程學(xué)院,2022.
[28] 李永麗,周洲,曲良建,等.貝萊斯芽孢桿菌Pm9生物防治潛力及全基因組分析[J].河南農(nóng)業(yè)大學(xué)學(xué)報(bào),2021,55(6):1081-1088.DOI:10.16445/ j.cnki.1000-2340.20210816.002.
[29] 林格兒,劉宏,劉海杰,等.暹羅芽孢桿菌高產(chǎn)γ-聚谷氨酸的發(fā)酵條件優(yōu)化[J].微生物學(xué)通報(bào),2022,49(8):3335-3345.DOI:10.13344/j.microbiol.china.211134.
