水培條件下火龍果根系形態和生理過程對供磷水平的響應
2024-01-01 00:00:00高翔楊永志李娟陳思如阮云澤高偉
熱帶作物學報 2024年5期






關鍵詞:磷;火龍果;根系形態;根系生理;養分吸收
中圖分類號:S667.9 文獻標志碼:A
磷是植物必不可少的大量營養元素,其在植物體內參與光合作用、呼吸作用、能量儲存和傳遞、細胞分裂等重要生理過程[1]。在以高產為目標的現代種植系統中,農民為保證作物充足的磷供應,往往通過大量施用化學磷肥而保證作物磷營養。然而,植物營養元素之間存在一定的相互作用。眾多研究表明,過量的磷吸收可能抑制作物對鋅、鐵等微量元素的吸收[2-3]。同時,大量吸收的磷會以植酸態磷的形式儲存在植物的果實和種子中,難以被動物和人體吸收利用,一定程度上是對磷資源的浪費。此外,磷是不可再生資源,全球磷資源儲量有限。因此,合理施用磷肥,不僅能夠最大化磷資源的利用效率,也能減少植酸等無效磷的累積并增加鋅和鐵含量,提高作物的品質和營養價值[4]。
土壤中全磷含量一般較為豐富,介于0.2~5.0 g/kg 之間,然而絕大部分磷都是有機態,難以被植物吸收利用。同時一些無機磷容易被鐵鋁氧化物、碳酸鈣和粘土礦物等吸附,造成土壤中能被植物吸收的磷含量很低[5]。針對上述情況,植物通過改變根系形態,有效地探索和活化土壤磷資源,提高自身磷營養[6-7]。PEREIRA 等[8]在對辣椒進行磷利用效率等方面的試驗中發現,磷脅迫情況下,根系總長度增加、根冠比提高、細根占根系長度的比例增加,同時降低了根系平均直徑。DING 等[9]研究發現細根和磷效率之間呈正相關關系,表明許多植物根系通過調節直徑的大小表現出對環境的適應[10]。除了根系的粗細之外,一、二級根系的生長也是根系在磷脅迫時會主動調整的方向之一。同時,植物為了對磷素的“覓食”達到高效,也會改變根際土壤的化學性質[11-12]。例如,小麥野生型和大豆低磷條件下分泌大量的質子,降低根際的酸堿度,同時分泌檸檬酸根和蘋果酸根等有機酸陰離子,從而促進磷的活化,最終提高植物的磷吸收[13-14]。因此,根系分泌物被認為是提高作物磷資源利用效率的重要途徑[15]。
火龍果是仙人掌科量天尺屬或蛇鞭柱屬植物,其外形奇特、風味獨特、營養豐富,富含維生素和水溶性膳食纖維,其特有的植物性白蛋白與甜菜紅素具有良好的保健功效。已有研究表明,火龍果果實中磷濃度顯著高于其他常見水果,可達350 g/kg[16-17]。同時,火龍果生產中,鮮果667 m2平均產量能達到5000 kg,高額的產量和果實的高磷含量要求根系從土壤中獲取大量磷資源。然而,國內外關于火龍果根系磷吸收利用的相關研究依然匱乏,對火龍果響應供磷水平的根系形態結構特性和生理過程缺乏系統性的研究。另一方面,生產中高磷條件下,是否會因為磷過量吸收,抑制火龍果對其他中、微量元素的吸收尚不清楚。因此,探究火龍果根系形態和生理過程對磷響應的作用機制,對實現火龍果高效利用磷以及平衡作物營養吸收具有重要的理論和實踐意義。
本研究采用營養液水培的方式,設定不同供磷水平,通過測定火龍果根系形態及根系分泌物、根系陽離子交換量等指標,探究火龍果根系形態和生理過程等根系生物學特性對不同梯度磷水平的響應特征。同時分析供磷水平與其他中、微量元素吸收之間的相互關系,為生產中提出合理的磷資源管理途徑提供理論依據。
1 材料與方法
1.1 材料
本試驗于2021年11月在海南大學實驗站進行,試驗地位于海南省海口市北部,典型的三角洲島嶼,屬于熱帶島嶼季風性氣候,年平均氣溫24.3 ℃,全年平均日照時數在2000 h 以上。試驗期間,晝夜溫度約為28 ℃/22 ℃,光照采用自然光。試驗為水培試驗,營養液為改良型1/2 強度的Hoagland 溶液[18],具體配方如下:1.75 mmol/L(NH4)2SO4,3.50 mmol/L KNO3,1.25 mmol/L K2SO4,2.50 mmol/L CaCl2 , 1.00 mmol/L MgSO4·7H2O ,0.035 mmol/L Fe-EDTA , 0.05 mmol/L H3BO3 ,0.01 mmol/L MnSO4,0.0008 mmol/L ZnSO4·7H2O,0.0002 mmol/L CuSO4·5H2O , 0.0021 mmol/LNaMoO3·2H2O,0.0048 mmol/L KI。火龍果生產中使用無性繁殖枝條扦插的方式種植,試驗使用的火龍果母枝為海南諾普信東河火龍果基地提供的大紅二號枝條。
1.2 方法
1.2.1 試驗設計 供試植株用雙氧水進行殺菌后,移植于1/2 強度改良型Hoagland 營養液中進行前期培養,待植株生長狀況穩定,挑選相似長度和相似重量的枝條,每個水培罐移植1 株。試驗設5 個磷水平濃度處理:(1)CK(0 mmol/L);(2)T1(0.01 mmol/L);(3)T2(0.10 mmol/L);(4)T3(0.50 mmol/L);(5)T4(1.00 mmol/L)[19]。試驗為隨機區組設計,每個處理重復5 次。試驗采用的磷源為磷酸二氫鉀,溶液中的其他基本元素在不同磷處理下保持完全相同,鉀離子用KCl補齊,酸堿度調節至pH=6.5,水培罐中溶液每5 d更新1 次,并24 h 不間斷通氣。
1.2.2 指標測定
(1)地上部生物量測定。火龍果幼苗定植180 d 后,于2022 年4 月收獲。收獲時沿莖基部剪下地上部,在105 ℃下殺青30 min后80 ℃烘干至恒重,隨后稱量地上部干重。
(2) 營養元素測定。烘干樣品粉碎后經H2SO4-H2O2 消煮,氮、磷、鉀含量分別用奈氏試劑比色法、鉬銻抗比色法、火焰光度法。其他營養元素測定方法采用原子吸收分光光度法,具體操作步驟參照鮑士旦《土壤農化分析》[20]。
(3)根系形態指標測定。根系樣品用去離子水沖洗干凈,置于50%的酒精溶液中,以防止根系變形。隨后置于4 ℃冰箱保存用于根系形態的分析。根系用平板掃描儀進行掃描(Epson ExpressionV800),分辨率設為400 dpi。所有圖片用WinRHIZO 軟件( Regent Instruments Inc.,Quebec, QC, Canada)進行分析,獲取根系長度、根表面積、根系直徑等參數[21]。
(4)根系有機酸測定。根系分泌物收集步驟:上午10:00(光期開始后2 h)開始收集;即將植株從培養溶液中取出,根系在無菌水中洗滌3次。控制環境條件與培養試驗相同的情況下,將根系轉至通氣、盛有500 mL 無菌去離子水的非透明容器中,使整個火龍果根部完全淹沒于無菌去離子水中。容器上覆蓋著鋁箔,為根系創造黑暗的條件。當收集1 h 后輕輕轉動根系1 次,收集2 h 后,將根放回培養容器中,將分泌物溶液搖勻后過濾。迅速放入?20 ℃冰箱冷凍,保存至測定時解凍[22] 。有機酸用高效液相色譜儀(HPLC)進行測定。溶液在0.45 μm 下進行過濾滅菌。采用高效液相色譜系統(Waters)(安捷倫1100,美國)。色譜條件如下:250 mm×4.6 mm(5 μm 粒徑)C18 反相柱,流動相:0.1%磷酸去離子水和乙腈98∶2(V/V),檢測器波長:214 nm,流速:1 mL/min,注入量:20 μL,柱溫度:35 ℃[23]。
(5)根系陽離子交換量(CEC)。準確稱取0.1000 g(雙子葉植物)樣品放入250 mL 的燒杯中并加幾滴蒸餾水使其濕潤,防止加入HCl 后樣品飄浮,影響測定結果。待樣品完全濕潤后,加入20 mL 0.1 mol/L HCl 攪拌5 min 至沉降,將上清液通過鋪有慢速定量濾紙的漏斗除去,將樣品留在燒杯中,用蒸餾水多次洗滌樣品,并移于漏斗上,繼續用蒸餾水洗至無Cl?為止(用AgNO3檢驗,一般需用蒸餾水約300 mL)即為H+根,同時做空白。將洗好的樣品連同漏斗移至250 mL 燒杯中將濾紙戳穿,用200 mL 1 mol/LKCl 將全部樣品洗入燒杯中。洗畢,把燒杯放至磁力攪拌器(或用玻璃棒)不斷攪拌,往裝有KCl 懸濁液的燒杯中滴入中性紅-溴百里酚藍混合指示劑10滴,后用0.01mol/L KOH 滴定溶液顏色至藍綠色,同時對空白進行滴定。根據以下公式進行計算[24]。
式中,CEC 為根系陽離子交換量(cmol/kg),C為KOH 滴定液的摩爾濃度;V 為KOH 滴定后的體積(mL);V0 為KOH 滴定前的初始體積(mL);m 為稱取根系樣品的質量(g);100 為換算系數。
1.3 數據處理
采用Excel 2021軟件進行數據處理,不同磷梯度水平處理間地上部生物量、營養元素吸收、根系形態和生理特性均使用SPSS 26.0軟件進行單因素方差分析,不同處理間的差異采用Turkey多重比較,利用Origin 9.0和R4.0軟件繪制圖表。
2 結果與分析
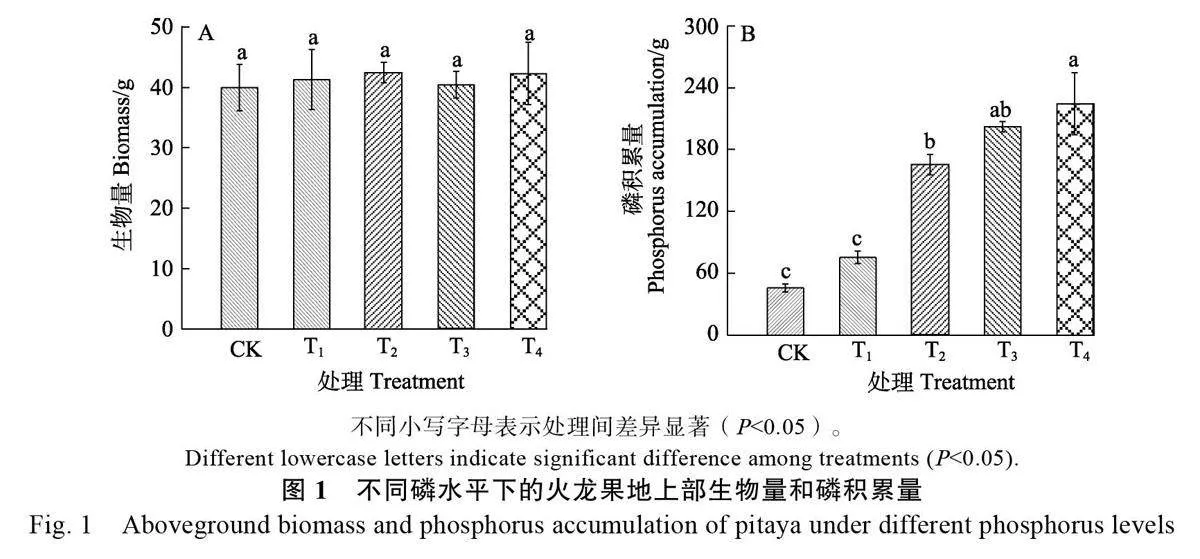
2.1 不同磷梯度水平處理對火龍果生物量和磷吸收的影響
整體趨勢上,相較于CK 處理,供磷后火龍果的地上部生物量并未增加,各處理之間地上部生物量差異不顯著(圖1A)。隨著供磷濃度的提升,火龍果磷的積累量呈現逐漸增加的趨勢,增幅在64.7%~390%之間;T4 處理累積量最高,為224 g/株,除T1 處理外的其他處理均與對照之間差異顯著(圖1B)。
2.2 不同磷水平處理對火龍果枝條磷濃度的影響
火龍果母枝磷濃度隨著磷水平濃度的增加而增加,T3、T4 處理相比CK 均差異顯著(Plt;0.05;圖2A)。隨著火龍果側枝生長,一級側枝較母枝的磷濃度在T3 和T4 處理濃度下分別平均增加21.03%和9.89%(圖2B)。當供磷濃度在T2 處理時二級側枝磷吸收濃度升高,T3 和T4 處理的吸收濃度未出現顯著降低(圖2C)。
2.3 不同磷水平處理對火龍果地上部各級枝條營養吸收變化
不同處理對母枝、一級側枝氮素吸收影響差異不顯著。二級側枝中,與對照和T1 相比,T3、T4 處理對枝條氮濃度影響差異顯著(Plt;0.05;表1),T3 處理下火龍果枝條氮濃度表現為最低,顯著低于CK、T1 和T2 處理。不同施磷濃度下,鉀吸收濃度在各部位枝條中差異均不顯著,整體來看一級側枝的鉀吸收濃度較高。母枝的鈣元素吸收濃度整體上大于一級側枝和二級側枝。母枝部位的T1 與T3 處理之間,鈣元素吸收濃度差異顯著(Plt;0.05),T3 處理相較于T1 處理鈣元素吸收濃度增長17.3%。不同施磷濃度下,鎂元素在母枝和一級側枝部位的吸收濃度差異不顯著。相較于CK,T3 和T4 處理的鎂吸收濃度在二級側枝處降低了26.2%,差異顯著(Plt;0.05)。不同施磷濃度下,鐵元素吸收濃度在各部位枝條中均差異顯著(Plt;0.05);在母枝及一級側枝中,鐵吸收濃度表現為隨著施磷濃度的升高呈逐漸下降的趨勢。相比CK 處理,T4 處理在母枝及一級側枝中鐵吸收濃度分別降低了39.7%、37.4%。在二級側枝部位,T3 處理的鐵濃度最大,為149 mg/kg;最低出現在T2 處理供磷水平,為109 mg/kg。鋅元素吸收濃度在母枝與一級側枝處均差異不顯著,一級側枝部位的鋅元素吸收濃度整體低于母枝,降幅為16.0%。不同處理對鋅元素吸收濃度的顯著差異出現在二級側枝部位(Plt;0.05)。CK處理的鋅元素吸收濃度最高,隨著磷濃度的升高,鋅元素吸收濃度均有不同程度的下降,在T3 處理達到最低,較CK 下降47.8 %。
通過分析火龍果地上部各營養元素之間的相關性并繪制熱圖,結果發現在母枝部位鈣元素吸收濃度與磷素吸收濃度呈現極強的正相關性。鐵元素在母枝和一級側枝部位與磷素呈現出極強的負相關性;二級側枝中,包括氮、鈣、鎂、錳、鋅在內的5種營養元素均與供磷水平呈現極強的負相關性(圖3)。
2.4 不同磷水平處理對火龍果根系形態特性的影響
根系生長隨著磷濃度的提高,整體呈現先增后減的變化趨勢。在T1 處理下,火龍果總根長為3196 cm,對比CK 處理增長了46.6%,T4 處理總根長最短,為1732 cm,各處理間存在不同程度的差異(Plt;0.05;圖4A)。總根表面積中,同樣出現先增后減的趨勢(圖4B)。T3 處理與T4 處理相比CK 處理分別降低了28.3%和29.6%。T1處理對比其他各處理明顯增加了總根系表面積,平均增長幅度為37.5%。通過將根系總長度按根系直徑大小進一步分級發現:≤0.5、0.5~≤1.0 mm根系直徑分級處,T1 處理的根長最大,相較于未施磷處理分別增長了47.1%、61.7%,T4 處理中≤0.5 mm 的細根為919 cm,對比T1 處理顯著降低59.2%(Plt;0.05;圖5)。
2.5 不同磷水平處理對火龍果根系生理特性的影響
如圖6 所示,T1 處理顯著增加了總有機酸含量,比未施磷處理增幅25.3%,與各處理之間對比差異顯著(Plt;0.05;圖6A)。根系CEC 隨著磷濃度的梯度性升高而提高,最高根系陽離子交換量出現在T1 處理。隨后,隨著供磷水平的提高,根系陽離子交換量隨之下降,到T4 處理時,表現為最低(圖6B)。
3 討論
3.1 不同磷水平處理對火龍果根系形態、生理以及磷吸收的影響
根系生長發育的好壞,一方面來源于作物自身的生物遺傳特性,另一方面來自外部環境變化[25]。大量研究結果表明,植物對磷吸收的多少與根系形態有密切關系,同時受環境磷水平高低的影響根系生長也會做出相應調控[26-27]。PEDRO 等[28]通過對小麥生物量、根系形態、酸性磷酸酶活性研究發現,低磷情況下小麥根系表現出更長、更大、更多分枝和直徑更細等特征。DING 等[29]在研究缺磷條件下水稻根系生長特性研究時發現,在低磷環境中,水稻的根生長加強,總長度增加,磷捕獲能力隨之提高,并最終變現為地上部生物量的提高。FERNANDEZ 等[30]在評估大豆、向日葵和玉米對磷的吸收效率差異中,3 種植物在低磷環境下均表現出更高的根冠比、比根長、根孔隙度和根通氣組織,以及更低的根密度。
本研究中,T1 處理增加了火龍果的總根長、總根系表面積,與上述小麥、水稻、大豆、向日葵等作物的研究結果[28-30]一致。火龍果方面,陳晨等[31]在土壤栽培條件下發現,供磷水平為0、25、50、100、200 mg/kg 條件時,根系在25 mg/kg和50 mg/kg 供磷水平下根長和根表面積達到最大,這與本研究T2 處理下根系生長達到最高水平相類似。與粗根相比,細根可以有效地增加根系的比表面積(即單位根重下的根系表面積),提高土壤中養分和水分的吸收,在形成和維持生理功能方面投入的C 較少,而粗根(常常木質化)投入的C 則相對較多[32]。T4 處理下火龍果降低了總根長、總根表面積,經過根系直徑等級細分后發現粗根數量有明顯的提升。
REICHERT 等[33]研究人員認為,中到高磷濃度環境中,植物群落主要集中于通過根系和叢枝菌根的磷獲取策略,而在中到低磷濃度環境中,則轉向通過磷酸酶和有機酸優先考慮磷的再吸收和挖掘策略。有研究發現,根系在低磷脅迫的條件下會導致根系分泌較多的有機酸使根際周圍難溶性的磷得到活化并最終被根系所吸收[34]。例如,玉米在低磷環境下促進根系檸檬酸分泌增加,生長介質pH 降低提高磷捕獲效率[35]。蠶豆、鷹嘴豆等豆科植物通過根系生理過程的變化,分泌質子、有機酸陰離子和酸性磷酸酶以活化土壤中被固定的難以被根系直接吸收利用的無機磷和有機磷[36]。本研究中,根系分泌物數量明顯增大是火龍果對低濃度磷的根際環境的響應,也是火龍果高效利用環境磷素的生理基礎。根的陽離子交換量(CEC)是單位數量根吸附的陽離子的物質的量,是一個反映根系的養分吸收能力的重要指標[37]。根系CEC 的高低直接關系到植物根系對土壤中陽離子的吸收,根系CEC 大,根系吸收土壤中的陽離子就多,同時間接性提高磷酸根離子的儲存,促進了磷酸根離子的吸收[38]。根系CEC 的多少直接影響植物對營養元素的吸收能力,根系CEC 大則植物通過根系吸收土壤中的營養元素就多[39]。本試驗中各供磷水平處理所表現的根系CEC 含量整體在20~35 cmol/kg 之間,依據《常見作物CEC 及吸收能力》[37],低于20 cmol/kg為吸收能力弱,20~35 cmol/kg 為中等吸收能力,高于35 cmol/kg 為強吸收能力的劃分標準來看,火龍果屬于中等吸收能力,然而T4 處理下,根系CEC含量顯著低于T1、T2 和T3 處理,表明根系的養分吸收能力受到了一定程度的抑制。
火龍果的磷營養方面,整體表現為:隨著供磷水平的提高,地上部母枝、一級側枝和二級側枝的磷濃度均呈上升趨勢, 當供磷水平達到0.1 mmol/L 時,母枝、二級側枝的磷濃度達到峰值水平,與更高強度供磷條件下枝條的磷濃度無顯著差異。一級側枝在T2 處理磷濃度低于T4 處理,可能是由于磷在植物體內是可移動元素,優先保障養分向新生枝條的轉移。
3.2 不同磷水平處理與火龍果地上部營養元素相關性
已有研究表明,磷肥過多會導致作物呼吸作用過于旺盛,造成繁殖器官提前發育引起作物過早成熟,此時作物干物質的消耗大于積累,氮素吸收減少消耗增加。因此,過量供磷不利于氮的吸收[40]。苗曉茸等[41]在探討滴灌條件下不同氮磷互作模式對綠洲區滴灌苜蓿生產性能及營養品質的影響中指出,過量的施磷可能會抑制紫花苜蓿對氮素的吸收和利用,從而增加苜蓿中的纖維含量,降低粗蛋白質含量。張清云等[42]在研究人工栽培甘草的最優混合施肥試驗中發現,同一施氮水平下,當磷過量時產量轉呈下降趨勢。本研究中,隨著供磷水平的提高,火龍果新生的二級枝條氮濃度顯著降低。供磷量大于T4 處理對火龍果的氮素營養產生顯著的負面作用。
本研究表明,鐵元素在母枝和一級側枝部位與磷素呈現負相關性。前人研究發現,磷-鐵之間存在顯著的拮抗作用,磷含量過高,可利用的鐵含量低,易造成作物缺鐵及低產雙重風險[43]。QI等[44]發現大量的磷遇到鐵和鋁基氧化物會成為具有低溶解性、固定作用的穩定態。而低磷情況下,有研究結果表明植株會提高對鐵的吸收[45]。GUO 等[46]的研究亦發現磷養分信號中心調控因子PHRs 和鐵養分信號核心調控因子HRZs 共同介導了磷-鐵信號的互作過程,磷養分匱乏時,低磷脅迫通過PHRs 間接抑制HRZs 的轉錄,導致磷饑餓響應相關基因大量激活,促進了鐵養分從地下向地上部的轉移;反之,磷的過量積累時則嚴重抑制鐵在植物體內的有效轉移。
火龍果的鈣營養狀況也顯著收到供磷水平的影響。一級側枝和二級側枝各營養元素之間的相關性分析結果表明,磷濃度與鈣濃度在母枝和一級枝條中呈現顯著正相關,而與新生的二級枝條呈現顯著的負相關。磷與鈣之間存在復雜的交互作用,不同作物體內磷鈣交互作用存在差異[47]。番茄幼苗、生菜及稻米三者體內的磷與鈣之間存在拮抗作用[48-49],而索炎炎等[50]探究了外源磷鈣肥配施對花生生長、產量和磷鈣吸收利用的影響,發現花生對磷、鈣吸收存在協同效應。本研究中磷和鈣的關系在老枝與新枝中表現不同,可能與鈣在植物體內移動性差有關,但是具體的機制還不清楚,有待進一步探究。
磷和鎂對植物生長發育都有很大的影響。鎂是植物葉綠素的中心原子,調控植物光合作用,參與大部分酶的激活過程,維持植物組織結構的穩定[51],過量的磷導致鎂元素吸收受到干擾[52]。隨著施磷劑量的提高,火龍果植株體內鎂含量出現了先增加后降低的趨勢[53]。研究還表明,鎂-磷互作通過AUX1、PIN2、PIN3 的信號途徑影響根的生長和方向性,通過生長素的再分配和累積,加速了細胞延展和根偏向性,進而調節主根的伸長和定向生長[54]。低鎂高磷環境下,將導致擬南芥初生根的生長偏離垂直方向[55]。因此,包括氮、鈣、鎂、錳、鋅在內的5 種營養元素均與磷元素吸收濃度呈現顯著的負相關性。
目前,磷-鋅拮抗作用機制的說法多樣且復雜,拮抗作用主要分為在土壤中和在植物體內2個過程[56]。有研究表明,隨著施磷量的增加,玉米籽粒中鋅、鐵和酚類物質的含量顯著降低[2],高磷條件下單位根干重吸鋅量減少,說明根系對鋅的吸收受供磷的影響[57],也會抑制根對鋅的吸收[58]。
綜上,盡管供磷水平對植株地上部的生長沒有產生顯著影響,即未影響到干物質量的積累。但T4 處理顯著抑制了火龍果植株鐵、鋅、鈣等元素的吸收,可能造成對果實品質的影響。因此,火龍果生產基地對于磷肥的運籌,基于“量入為出”的原則下進行適度削減。
本研究僅對火龍果苗期枝條生長和養分積累做了相關研究,供磷水平對果實磷吸收、產量以及中、微量元素含量和品質的影響尚不清楚,有待進一步加強。
4 結論
不同磷水平處理對火龍果植株的養分吸收量、根系形態、生理過程存在明顯的劑量效應,即:T1處理條件下,火龍果根系總根長、根表面積、有機酸和根系陽離子交換能力提高,T2處理表現出根系直徑更粗,陽離子吸附能力更強的趨勢。
不同磷水平處理的火龍果地上部磷積累量及各部位枝條的磷吸收濃度均高于未施磷處理;另一方面,隨著磷濃度增加,火龍果對磷的吸收量沒有發生顯著變化;T3 處理和T4 處理對火龍果生長積累影響不顯著,且對氮、鈣、鎂、鐵、鋅等營養元素的吸收出現了抑制現象,容易對火龍果后續的生長發育造成中、微量元素營養不良的影響。
