桂花葉斑病病原菌的鑒定、生物學特性及防治藥劑篩選
2024-01-01 00:00:00劉向國
東南園藝 2024年4期


摘要要:【目的】桂花葉斑病嚴重影響桂花的觀賞價值,為明確其病原菌,并篩選出可用于防治桂花葉斑病的殺菌劑。【方法】從福州市金山公園采集桂花葉斑病樣品,切片鏡檢,采用組織分離法獲得真菌菌株,通過柯赫氏法則驗證所獲菌株是否為桂花葉斑病的致病菌;綜合形態特征和多基因(ITS、TUB、TEF、 CAL 和 HIS )序列的系統發育分析將該菌鑒定到種;在此基礎上,測定了該菌的生物學特性,采用含藥平板法測定了4種殺菌劑對病原菌的毒力。【結果】明確了菌株FZ-GH-1是福州桂花葉斑病的病原菌,其被鑒定為Diaporthe grandiflori;FZ-GH-1在12 h光暗交替和全黑暗條件下菌絲生長速率最快,生長最適溫度為24~28℃,最適pH值為5.0,生長所需最適碳源為葡萄糖,最適氮源為硝酸鈉和蛋白胨;在供試藥劑中,70%甲基硫菌靈可濕性粉劑和450 g/L咪鮮胺水乳劑對FZ-GH-1菌絲生長的EC50分別為0.108 mg / L
和0.053 mg/L,明顯低于30%代森錳鋅可濕性粉劑和20%三唑酮乳油。【結論】確定了福州桂花葉斑病的病原菌為Diaporthe grandiflori,甲基硫菌靈和咪鮮胺對該菌抑制作用強,研究結果可為后續研究桂花葉斑病的發病規律和化學防治提供理論依據。
關鍵詞:桂花葉斑病;間座殼屬;多基因系統發育分析;生物學特性;室內毒力測定
中圖分類號:S436.8" " " " " " " " " " " "文獻標識碼:A" " " " " " " " " " " "文章編號:2095-5774(2024)04-0250-10
Identification,Biological Characterization,and Fungicide Screening of Pathogen Causing Leaf Spot Disease of Osmanthus fragrans(Thunb.)Lour.
Liu Xiangguo
(Administrative Office of Jinshan Park in Fuzhou,Fuzhou,Fujian 350001,China)
Abstract:【Objective】Leaf spot disease of Osmanthus fragrans seriously affects its ornamental value. This study aims to identify its pathogen and screen fungicides for controlling the disease.【Method】Spotted leave samples of
O. fragrans were collected from Jinshan Park in Fuzhou and sample section was examined under a microscope. Fungal strains were obtained by tissue isolation method,and Koch’s postulates were used to confirm the pathogenicity of the obtained strains. The strain was identified to species level though morphological characteristics and phylogenetic analysis of multilocus sequences(ITS,TUB,TEF, CAL and HIS). On this basis,the biological characteristics of the strain were determined. The toxic effects of four fungicides against pathogen were evaluated in vitro. 【Result]】The strain FZ-GH-1 was clarified as the pathogen of leaf spot disease of O. fragrans,and was identified as Diaporthe grandiflori. The mycelial growth rate of FZ-GH-1 was fastest under 12-hour light/dark alternation and complete darkness conditions,with the optimal growth temperature being 24-28℃,optimal pH 5.0,optimal carbon source being glucose,and optimal nitrogen source being sodium nitrate and peptone. Of the tested fungicides,the EC50 of 70% thiophanate -methyl wettable powder and 450 g/L prochloraz aqueous emulsion on the growth of FZ-GH-1 mycelia was 0.108 mg/L and 0.053 mg/L,respectively,which was significantly lower than that of 30% mancozeb wettable powder and 20% triazolone emulsion. 【Conclusion】This study confirmed D. grandiflori as the pathogen of leaf spot disease of O. fragrans,and thiophanate-methyl and prochloraz had strong inhibition effects on this fungus. The findings of this study provided a theoretical basis for investigating the pathogenesis and chemical control of leaf spot disease of O. fragrans in future research.
Key words:Leaf spot disease of Osmanthus fragrans;Diaporthe grandiflori;Multi-loci phylogenetic analyses;Biological characteristics;Fungicides screening
桂花(Osmanthus fragrans (Thunb.) Loureiro)又名木犀,是木犀科(Oleaceae)木犀屬(Osmanthus)植物,其花為名貴香料,并可作食品香料[1]。桂花原產我國西南部,在四川、廣東、廣西、云南、湖北等多個省區均有分布,國外也普遍栽種,如印度、尼泊爾、柬埔寨等地,具有很高的經濟、觀賞和藥用價值[2]。桂花在生長過程中容易遭受病害的感染,其中葉斑類病害是桂花常見病害,嚴重時會導致樹葉枯死,影響桂花的觀賞價值和經濟價值[3]。該類病害老葉更易感染,多發生于4-10月份,7-8月份高溫高濕的環境下易大范圍發生,在我國臺灣、廣西、江西、福建、安徽等多個省份均有發生[4-8]。前人曾研究報道交鏈孢菌(Alternaria sp.)、細交鏈孢菌 (A. tenuissima ( Fr.) Wiltshi)、葡萄座腔菌(Botryosphaeri dothidea)、木犀生尾孢(Cerospora osmanthicola P.K. Chi et Pai)、Diaporthe fusicola、變葉木葉點霉(Phyllosticta ghaesembillae Koorders)、木犀生葉點霉(P. osmanthicola Train.)等真菌可引起桂花葉斑病,造成的病害癥狀相似,病原難以明確[9-15]。不同的病原真菌對殺菌劑的敏感性存在差異,對于不同真菌引起的桂花葉斑病,應篩選相應的防治藥劑,采取有針對性的防治措施和方法。在化學防治方面,可在發病初期用波爾多液進行預防,發病嚴重時可施用代森鋅、波爾多液、多菌靈、高錳酸鉀、苯來特、可殺得和阿米西達等,但對于不同的病原真菌而言,防治藥劑會有所不同[4,16]。基于此,本文對福州市金山公園桂花葉斑病的病原菌進行了系統地種類鑒定和生物學特性分析,并測定了4種常用殺菌劑對病原菌的抑制作用,旨在明確福州桂花葉斑病的病原,并為制定科學的病害防治措施提供依據。
1 材料與方法
1.1 材料
1.1.1 供試病樣
2021年9月,從福州市金山公園采集的具有桂花葉斑病典型癥狀的樣品。
1.1.2 培養基
主要采用馬鈴薯葡萄糖瓊脂培養基(Potato Dextrose Agar,PDA)、察氏培養基和燕麥培養基(Oats Agar,OA),依據方中達(1998)描述的方法制備[17]。
1.1.3 儀器設備
離心機(JIDI-17R,廣州吉迪儀器有限公司);PCR 儀(ETC821,蘇州東勝興業科學儀器有限公司);凝膠成像儀(HT8500,北京鴻濤基業科技發展有限責任公司);鴻基電泳儀(HT-SUB03,北京鴻濤基業科技發展有限責任公司);生物顯微鏡(UB203i,重慶澳浦光電技術有限公司);生化培養箱(TRY-250,寧波賽福實驗儀器有限公司);植物生長箱(ZRY-380,寧波賽福實驗儀器有限公司)。
1.1.4 殺菌劑
80%代森錳鋅可濕性粉劑(四川瀾爾科技有限公司)、45%咪鮮胺水乳劑(上海滬聯生物藥業(夏邑)股份有限公司)、70%甲基硫菌靈可濕性粉劑(江蘇龍燈化學有限公司)和20%三唑酮懸浮劑(四川瀾爾科技有限公司)。
1.2 方法
1.2.1 樣品采集與病原菌的分離純化
2021年9月在福州市金山公園進行病害調查時,觀察拍攝桂花葉斑病的田間癥狀,同時采集具有典型癥狀的葉斑病樣品。
用無菌水清洗植物病樣表面后擦干,在超凈工作臺上用無菌剪刀剪取植物病樣病健交界處
5 mm×5 mm的組織塊,依次放入75%酒精、2%次氯酸鈉、75%酒精中消毒30 s、1 min、30 s,后用滅過菌的ddH2O各漂洗 3次,每次漂洗30 s,再用無菌濾紙將其表面水分吸干,最后放入事先備好的9 cm的PDA平板上,每皿PDA平板上放4塊組織塊。將PDA平板倒置放入26 ℃、12 h 光暗交替的生化培養箱內培養,3~4 d 后挑取植物組織塊周圍有明顯特征差異的菌絲塊轉移到新的PDA平板中,置于26℃、12 h光暗交替的生化培養箱內培養。培養3~4 d后觀察純化后培養的菌落是否為單一純菌落,若為單一純菌落,則表示純化成功;若不是,則需繼續純化,直到每個培養皿上的菌落都是單一純菌落。從培養7 d的純化菌株挑取菌絲塊,將其轉移至PDA凍存管斜面中,在26 ℃、12 h 光暗交替的生化培養箱內繼續培養5~7 d后,置于4 ℃冷藏保存。
1.2.2 病原菌的致病性測定
采用柯赫氏法則驗證分離菌株的致病性。將供試菌株置于26 ℃、12 h 光暗交替的生化培養箱內培養5 d,用打孔器在菌落邊緣打取直徑0.5 cm菌餅,備用。摘取健康的桂花葉片用清水洗凈后擦干表面;在干凈的培養皿內放入滅過菌的濾紙并用無菌水潤濕,然后將準備好的葉片放入培養皿中,用水噴濕葉片表面;用10 μL 槍頭在葉片葉脈左右兩邊上、中、下三個部位進行輕微造傷,將菌餅放于右側造傷處,左側放上同樣大小的PDA培養基塊作為空白對照,每個葉片3個重復;將培養皿蓋好蓋子放于26℃的植物生長箱中,每日進行噴水保濕,培養7 d 后觀察發病情況。再采用組織分離法分離發病葉片的病原菌,觀察分離后獲得的真菌菌株的形態特征是否與最初分離純化得到的形態特征吻合。
1.3 病原菌的鑒定
1.3.1 形態特征觀察
將供試菌株在26 ℃、12 h 光暗交替的生化培養箱內培養7 d,觀察菌落形態特征,并拍照記錄。在顯微鏡下觀察供試菌株的孢子產生情況,如已產孢,觀察記錄產孢結構和孢子形態,并進行初步鑒定。
1.3.2 多基因序列分析
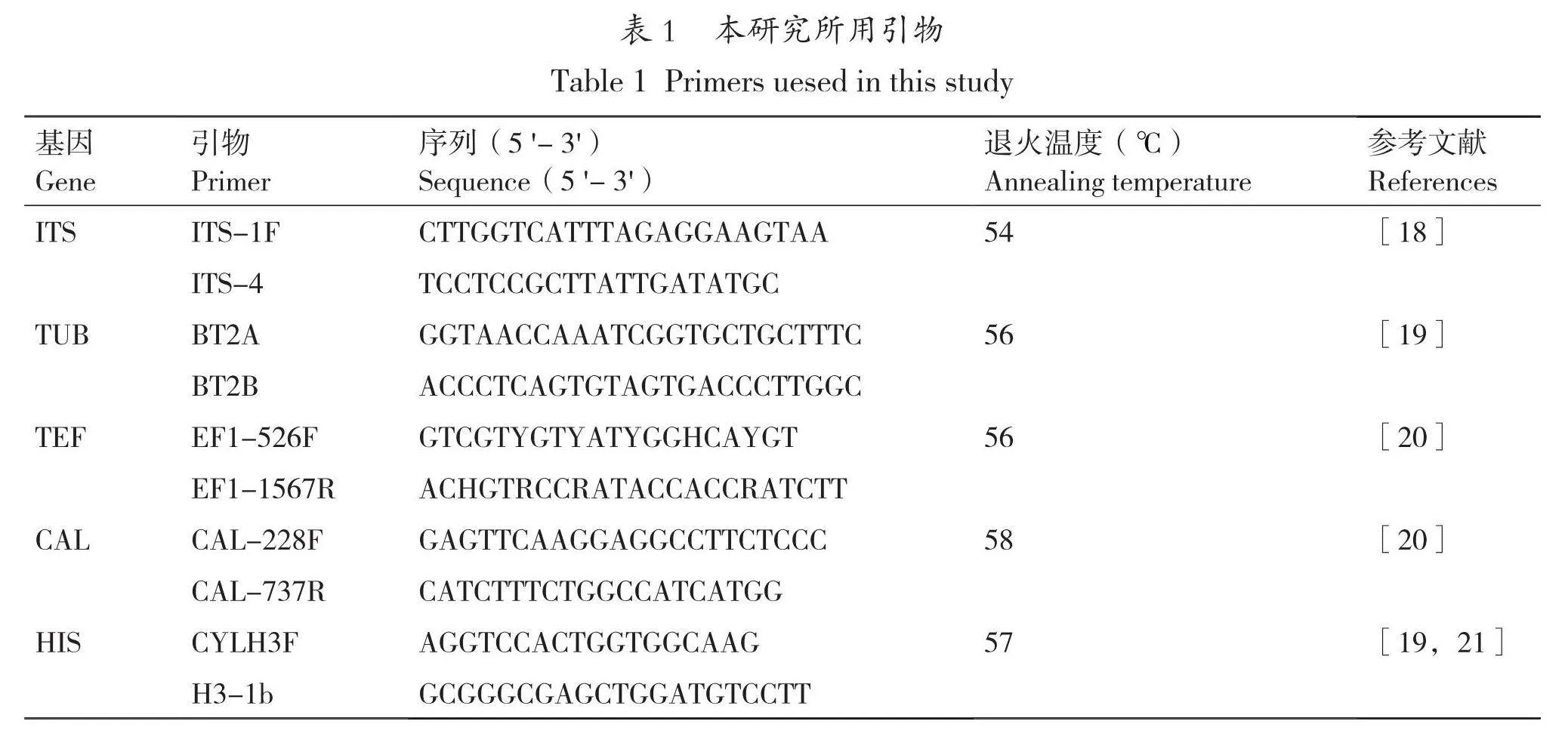
選擇北京聚合美生物科技有限公司提供的試劑盒,并按照其說明進行DNA提取。用通用引物ITS1/ITS4對提取的菌株DNA進行PCR擴增。經1.2%瓊脂糖凝膠電泳檢測成功后,將PCR擴增產物送至尚亞生物技術有限公司進行測序。測序結果在NCBI數據庫(https://blast.ncbi.nlm.nih.gov/Blast.cgj)中進行BLAST比對分析。根據序列同源性和形態特征初步確定病原菌種類。然后在NCBI搜索該種病原菌的ITS、TUB、TEF、CAL、HIS序列,將所獲得的序列作為靶標序列設計特異擴增引物。各基因對應的引物名稱及其序列見表1。25 μL PCR反應體系包括2×Taq PCR Master Mix 12.5 μL,ddH2O 9.5 μL,10 μmol/L上下游引物各1 μL,模板DNA 1 μL。反應程序:94℃預變性5 min;94℃變性30 s,相應退火溫度(表1)下退火30 s,72℃延伸45 s,ITS 32個循環,TUB、TEF、CAL和HIS 35個循環;72℃終延伸10 min。PCR擴增產物用1.2%瓊脂糖凝膠電泳檢測。檢測成功后,將 PCR 擴增產物送至尚亞生物技術有限公司測序。
檢查測序結果,及時修正,并將5個基因的序列上傳到NCBI。用Clustalx v.1.83軟件對單個基因的序列與靶標序列進行比對[18],再在BioEdit中對比對后的序列進行人工校正[19],選擇Diaporthella corylina作為外群種,然后在IQ – TREE中構建最大似然樹(maximum likelihood,ML)[20],和在MrBayes中構建貝葉斯系統發育樹(Bayesian inference,BI)[21]。接著使用PhyloSuite v1.2.1[22]的Concatenate Sequence 選項將5個基因序列進行串聯,用Partitionfinder2計算每個基因的最佳核苷酸替換模型以及相關參數,隨后用MrBayes建立多基因的貝葉斯系統發育樹和IQ-TREE建立多基因的最大似然樹,最后將所得的系統發育樹在Powerpoint中進行美化。
1.4" 病原菌生物學特性研究
1.4.1 溫度對菌絲生長的影響
將供試菌株26 ℃、12 h 光暗交替的生化培養箱內培養5 d,用打孔器在菌落邊緣打取直徑0.5 cm菌餅,將菌餅轉接至PDA培養基平板,分別置于24 ℃、26 ℃、28 ℃、30 ℃ 和32 ℃ 等5個溫度梯度,12 h 光暗交替的生化培養箱中培養7 d。采用十字交叉法測量菌落直徑。每個處理重復3次[22-24]。
1.4.2 光照對菌絲生長的影響
將供試菌株26 ℃、12 h 光暗交替的生化培養箱內培養5 d,用打孔器在菌落邊緣打取直徑0.5 cm菌餅,將菌餅轉接至PDA培養基平板,分別置于持續黑暗、12 h 光暗交替和持續光照等3個光照條件,28 ℃的生化培養箱培養7 d。采用十字交叉法測量菌落直徑。每個處理重復3次。
1.4.3 pH值對菌絲生長的影響
將供試菌株26 ℃、12 h 光暗交替的生化培養箱內培養5 d,用打孔器在菌落邊緣打取直徑0.5 cm菌餅,分別轉接至pH值為5.0、 6.0、7.0、8.0、9.0等5個pH值梯度的PDA培養基平板上,置于28 ℃、12 h 光暗交替的生化培養箱中培養 7 d。采用十字交叉法測量菌落直徑。每個處理重復3次[22-24]。
1.4.4 碳、氮源對菌絲生長的影響
將察氏培養基中蔗糖換成相同含量的葡萄糖、乳糖、麥芽糖和可溶性淀粉,并設置不加任何碳源的培養基作為對照組,備用。將其中硝酸鈉換成相同含量的硝酸鉀、蛋白胨、氯化銨、尿素和硫酸銨,并設置以不加任何氮源的培養基為對照,備用。將供試菌株在26 ℃、12 h 光暗交替的生化培養箱內培養5 d,用打孔器在菌落邊緣打取直徑0.5 cm菌餅,分別轉接至上述培養基平板上,置于28 ℃、12 h 光暗交替的生化培養箱中培養7 d,采用十字交叉法測量菌落直徑,每個處理重復3次[22-24]。
1.5 室內防治藥劑篩選
將供試殺菌劑用無菌水配置為有效成分含量為10 000 mg/L的母液,置于65℃恒溫水浴待用;取事先制備的PDA培養基加熱融化;分別將供試殺菌劑稀釋為系列濃度倒入培養基,制成終濃度為0.01、0.05、0.10、0.50、1.00、5.00、10.00 mg/L的系列含藥培養基并倒板備用。將供試菌株在26 ℃、12 h 光暗交替的生化培養箱內培養5 d,用打孔器在菌落邊緣打取直徑0.5 cm菌餅,分別將菌餅轉接至含藥PDA培養基平板,置于28 ℃、12 h 光暗交替的生化培養箱中培養7 d。以無菌水為對照。采用十字交叉法測量菌落直徑。每個處理重復6次。利用Excel計算供試殺菌劑對菌絲生長的抑制率。
1.6 數據分析
以供試藥劑各濃度的對數值為自變量(X),以菌絲生長抑制率對應的機率值為因變量(Y) 建立毒力回歸方程,并計算各藥劑對供試菌株的抑制中濃度(EC50) 及相關系數(r)。采用 SPSS統計軟件對在不同溫度、pH、光照、碳氮源等培養條件下得到的試驗數據以及 EC50進行差異顯著性分析。
2 結果與分析
本研究在采集到的桂花葉斑病樣品中分離培養獲得了20個菌落形態相似的菌株,通過菌落形態觀察和ITS序列比對初步確定這20個菌株為同一種菌,且為間座殼屬真菌。后續的研究均用菌株號為FZ-GH-1的菌株。
2.1 形態特征描述
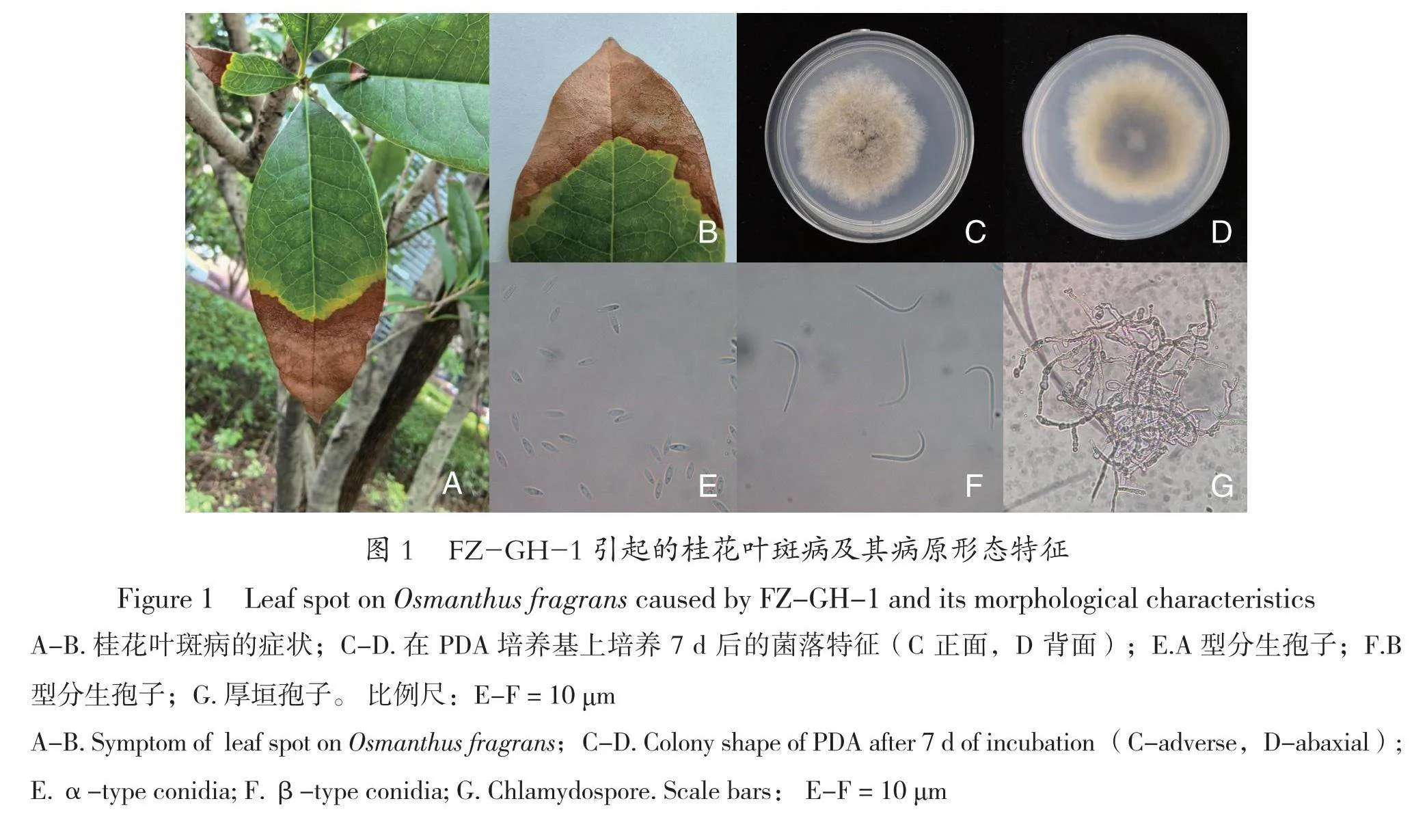
病害癥狀:病害從葉尖開始,逐步蔓延至接近葉片中間處。病斑為淡黃褐色至淡褐色,病健交界處為黃褐色,病斑上密生黑色小點(見圖1 A-B)。
培養性狀:FZ-GH-1菌絲生長較快,在28 ℃、12 h 光暗交替的人工氣候培養箱中培養7 d 后,菌落直徑達6.5 cm左右。菌落正面初期為灰色,后期變為灰綠色,氣生菌絲較多,從菌落中間開始產生黑色顆粒狀物,并向四周擴散(見圖1 C);菌落背面初期為灰色,后期中間部分顏色較深,灰黑色至灰綠色,邊緣顏色較淺,多為灰色(見圖1 D)。
顯微特征:在PDA培養基上,較難產生分生孢子,可產生厚垣孢子(見圖1 G)。在OA培養基上,可產生A型和B型兩種類型的分生孢子。A型分生孢子梭形至長紡錘狀,無色,內含1~2個油滴,大小為(6.6~10.3)μm×(1.4~2.9)μm,能萌發(見圖1 E);B型分生孢子細長,鉤狀,大小為(19.7 ~25.6)μm×(0.9~1.5)μm,不能萌發(見圖1 F)。
Sun等人2021年發表的新種Diaporthe grandiflori(菌株號SAUCC194.22)[28],A型分生孢子的大小為8.3 μm×(2.8~3.3)μm,B型分生孢子的大小為 (21.5~30.5)μm×(1.5~2.1)μm,菌株FZ-GH-1形態特征和分生孢子形態結構與其相似,因此,將菌株FZ-GH-1初步鑒定為Diaporthe grandiflori。
2.2 病原菌的致病性測定
桂花葉片在接種FZ-GH-1的第7 d 后出現明顯病斑,且有擴大趨勢。病斑內部淡褐色,外部棕褐色(見圖2)。對發病部位進行再分離發現,獲得的菌株的形態特征與最初分離得到的菌株的形態特征一致,說明FZ-GH-1是引起桂花葉斑病的病原菌。
2.3病原菌的分子鑒定
對菌株FZ-GH-1 的ITS、TUB、TEF、CAL 和
HIS基因進行測序,分別獲得909 bp、 659 bp、981 bp、485 bp和416 bp的序列,合并后做系統發育分析的5個基因序列(含gap)總長為3 450 bp。將菌株FZ-GH-1和其他間座殼屬真菌一起構建了ITS、TUB、TEF、CAL 和HIS 等5個基因聯合的系統發育樹,菌株FZ-GH-1和Diaporthe grandiflori(菌株號SAUCC194.22)形成獨立分支,最大似然支持率為100%,后驗概率為0.99(見圖3)。
此發育樹以 Diaporthella corylina(CBS 121124) 作為外群種;系統發育樹上顯示貝葉斯后驗概率(PP)≥ 0.80 和最大似然支持 (MLBS)≥ 50%,貝葉斯后驗概率低于 0.80 和最大似然支持低于 50% 用 - 表示。本研究獲得的菌株用紅色標記;比例尺表示每個位點的預期核苷酸替換數。
2.4 病原菌的生物學特性
觀察比較菌株FZ-GH-1在不同培養條件下的菌絲生長情況,發現供試的幾個條件均能影響FZ-GH-1的菌絲生長。
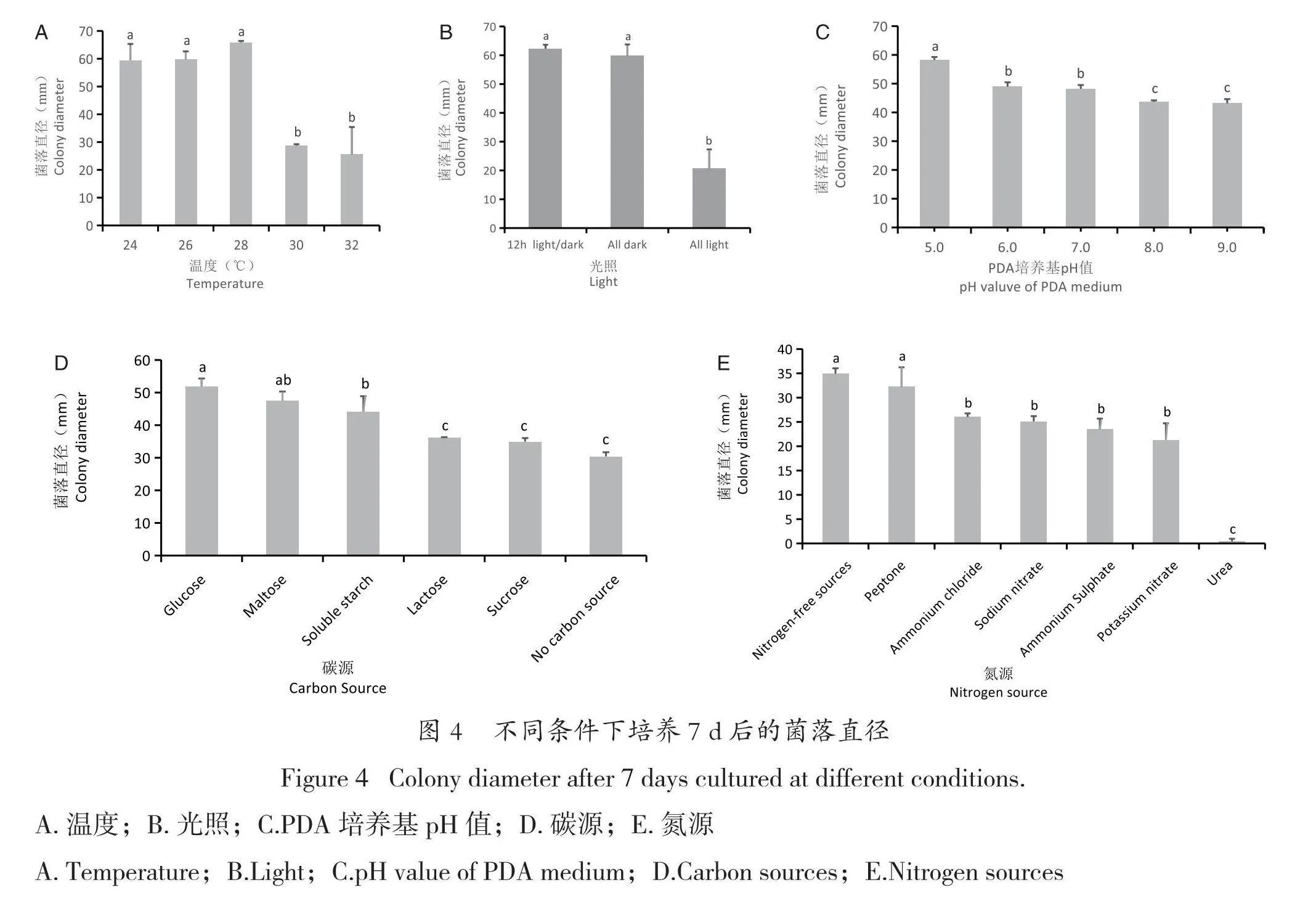
FZ-GH-1在24 ℃、26 ℃、28 ℃、30 ℃ 和32 ℃ 等5個溫度中均能生長,但在24~28 ℃ 菌絲生長快,菌落直徑顯著大于30 ℃和32 ℃的菌落直徑(見圖4A)。
FZ-GH-1對光照要求不高,在光照條件為12 h
光暗交替和全黑暗情況下菌絲均能快速生長,菌落直徑差異不顯著,但其菌落直徑顯著大于全光照培養條件下的菌落直徑(見圖4B)。
FZ-GH-1在pH值 5.0~9.0的條件下均能生長,在pH值為5.0時生長速度最快,其菌落直徑與在其他pH值條件下生長的菌落直徑差異顯著;pH值為6.0和7.0條件下的菌落直徑相似,雖小于pH值為5.0的菌落直徑,但顯著大于pH值為8.0和9.0的菌落直徑(圖4C)。
供試的幾種碳源中,葡萄糖最利于FZ-GH-1的菌絲生長,其菌落直徑顯著大于其他供試碳源的菌落直徑;其次是麥芽糖和可溶性淀粉;蔗糖、乳糖和不加糖的菌落直徑則顯著較小(圖4D)。
在選擇的幾種不同氮源中,硝酸鈉和蛋白胨作為氮源時,FZ-GH-1的菌絲生長最快,菌落直徑顯著大于其他氮源;在氮源為尿素的培養基中,FZ-GH-1的菌絲幾乎不生長,菌落直徑顯著小于其他供試氮源(圖4E)。
2.5 室內防治藥劑篩選
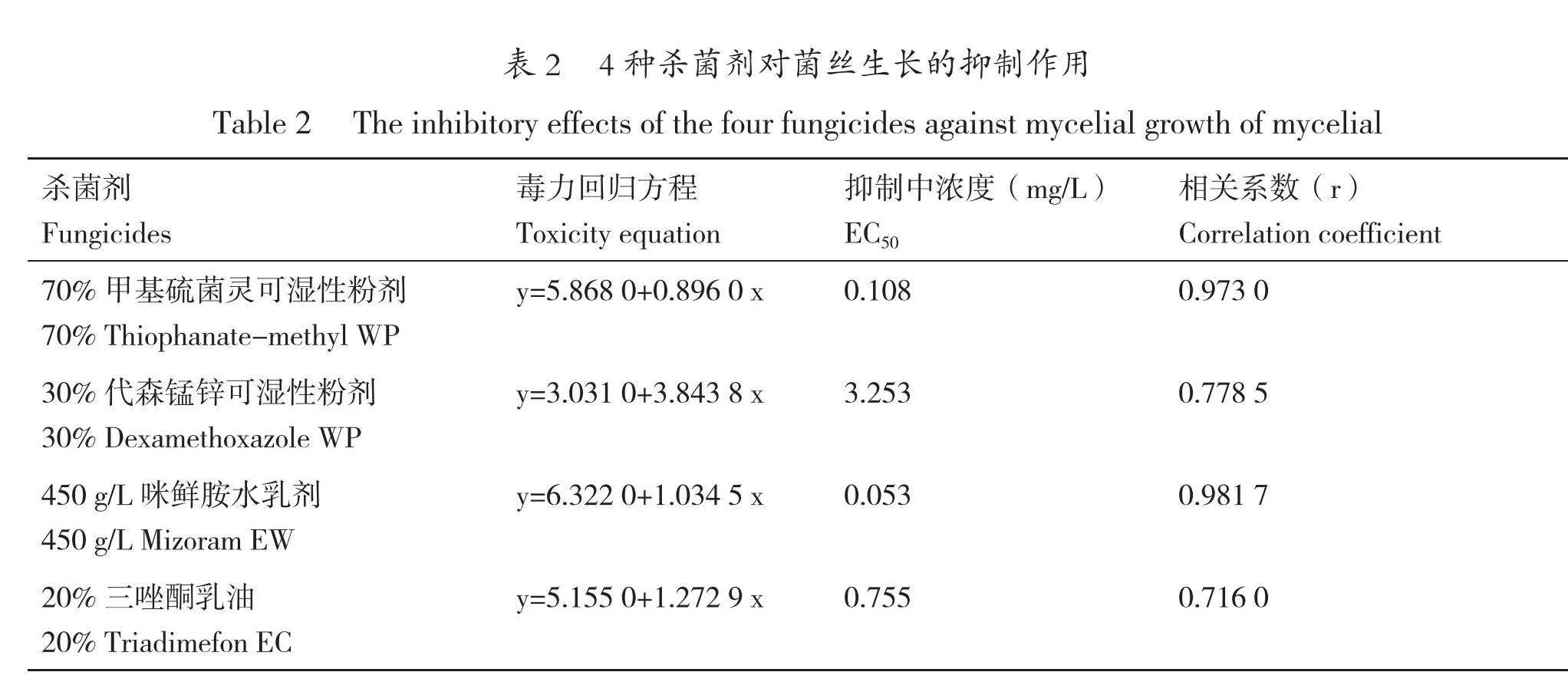
室內藥劑毒力測定結果表明,70%甲基硫菌靈可濕性粉劑和450 g/L咪鮮胺水乳劑對菌絲生長的抑制作用強,EC50分別為0.108 mg/L和
0.053 mg/L;30%代森錳鋅可濕性粉劑和20%三唑酮乳油對菌絲生長的EC50分別為3.253 mg/L和0.755 mg/L,30%代森錳鋅可濕性粉劑在稀釋倍數達到10 000倍時菌絲已可以較為正常地生長,但20%三唑酮乳油不同稀釋倍數對菌絲生長抑制作用不明顯(見表2)。
3 結論與討論
本研究從福州市金山公園內采集了有葉斑病典型癥狀的桂花葉片,通過分離純化,獲得了葉斑病的病原菌FZ-GH-1菌株,結合形態特征和5個基因(ITS、TUB、TEF、CAL和HIS)的系統發育分析,將FZ-GH-1鑒定為D.grandiflori。間座殼屬(Diaporthe)真菌隸屬于子囊菌門(Ascomycota)盤菌亞門(Pezizomycotina)糞殼綱(Sordariomycetes)間座殼目(Diaporthales)間座殼科(Diaporthaceae),無性階段為擬莖點霉屬(Phomopsis)[26]。該屬真菌的分類跟其他真菌類群相同,以前主要以形態特征為主,現在綜合形態特征和多基因系統發育分析可以更準確的分類鑒定到種[23,27]。
此外,本研究還系統地研究了溫度、光照、pH值、碳氮源等對FZ-GH-1的菌絲生長的影響,結果發現FZ-GH-1的適宜生長溫度為24~28℃,光暗交替培養和全黑暗培養有利于該菌株的菌絲生長,這與賈鑫宇等的研究結果類似[28]。FZ-GH-1的最適生長pH值為 5.0,在pH 8.0的條件下則生長緩慢,說明在酸性環境下有利于病原菌生長,可適當噴堿性殺菌劑或生長調節劑來控制桂花葉斑病。最適碳源為葡萄糖、麥芽糖和可溶性淀粉,最適氮源為硝酸鈉和蛋白胨。病原菌FZ-GH-1對碳氮源的要求不嚴格,這說明D. grandiflori的潛在寄主可能較為廣泛,為防控桂花葉斑病,需要進一步明確該病原菌的寄主范圍和侵染來源。
為了更好地防治該病害,本研究對FZ-GH-1菌株進行室內藥劑毒力測定試驗,結果發現70%甲基硫菌靈可濕性粉劑和450 g/L咪鮮胺水乳劑2種藥劑對于病菌的抑制效果好。年文楷等研究發現氟啶胺對D. amygdali的抑菌活性最高[29],這說明不同種類的間座殼菌對殺菌劑的敏感性不同。本研究中僅測定了4類藥劑對FZ-GH-1的抑制效果,后面仍需進行更多種類的殺菌劑篩選,以方便防治桂花葉斑病的過程中交替用藥,避免產生抗藥性。
總之,本研究證明福州市金山公園桂花葉斑病的病原菌為D. grandiflori,并明確了其基本生物學特性和2種有效防治藥劑,為該病的防控提供了理論依據。
參考文獻:
[1]中國植物志編輯委員會,中國科學院.中國植物志[M].北京:科學出版社,1993.
[2]任全進,朱洪武,于金平.桂花資源的利用價值[J].中國野生植物資源,1999(4):34-35.
[3]易雄輝.桂花葉斑病的發生與防治[J].特種經濟動植物,2006(3):42.
[4]楊玉婷,黃淑容,龍儀珂,等.桂花病蟲害綜合防治研究進展[J].熱帶農業科學,2022,42(3):73-82.
[5]Ariyawansa H A,Tsai I,Hozzein W N,et al. Leucaenicola osmanthi sp. nov. (Bambusicolaceae,Pleosporales),causing leaf spot of Osmanthus fragrans in Taiwan[J]. Phytotaxa,2020,437(1): 23-31.
[6]謝玲,唐晨光,岑貞陸,等.廣西桂花葉枯病病原菌鑒定及其生物學特性研究[J].西南農業學報,2009,22(5):1358-1362.
[7]Tang Z,Sun X,Ye M,et al. First report of leaf spot on Osmanthus fragrans caused by Colletotrichum gloeosporioides in Jiangxi Province of China[J]. Plant Disease,2018,102(5): 1035.
[8]張橋.桂花葉枯病病原菌生物學特性研究[J].現代農業科技,2016(11):153-154.
[9]袁蒲英,朱天輝.桂花葉枯病病原菌鑒定及生物學特性研究[J].四川林業科技,2008(4):21-28.
[10]王曉梅,黃琦,李玉. 桂花葉斑病病原鑒定及其生物學特性研究[J]. 吉林農業大學學報,2011,33(2): 151-157.
[11]朱天輝.園林植物病理學[M].北京:中國農業出版社,2003: 168-169.
[12]李永麗,周洲,源春彥,等.引致桂花葉斑病的葡萄座腔菌生物學特性及藥劑毒力測定[J].北方園藝,2014(19):117-120.
[13]Si Y Z,Guo X P,Li D W,et al. First report of Diaporthe fusicola causing leaf blotch of Osmanthus fragrans in China[J]. Plant Disease,2021,105(4): 1193.
[14]杜亞敏.引起獼猴桃枝枯和葉斑的間座殼菌種類鑒定及致病性研究[D].武漢:華中農業大學,2020.
[15]李媛,石凌波,費諾亞,等.藍莓間座殼莖潰瘍病病原鑒定及生物學特性研究[J].植物保護,2017,43(1):89-94.
[16]楊曉.桂花樹主要病蟲害的發生和防治技術[J].現代農業,2018(11):12-13.
[17]方中達.植病研究方法[M].北京:中國農業出版社,1998.
[18]White T J,Bruns T,Lee S,et al. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics[M]. In: PCR Protocols: A Guide to Methods and Applications. (Innis MA,Gelfand DH,Sninsky JJ,White TJ eds). Academic Press: New York,1990: 315–322.
[19]Glass N L,Donaldson G C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes[J]. Applied and Environmental Microbiology,1995,61: 1323–1330.
[20]Carbone I,Kohn L M. A method for designing primer sets for speciation studies in filamentous ascomycetes[J]. Mycologia,1999,91: 553-556.
[21]Crous P W,Groenewald J Z,Risède J,et al. Calonectria species and their Cylindrocladium anamorphs species with sphaeropedunculate vesicles[J]. Studies in Mycology,2004,50: 415-430.
[22]Santos L,Alves A,Alves R. Evaluating multi-locus phylogenies for species boundaries determination in the genus Diaporthe. PeerJ,2017,5: e3120.
[23]Huang F,Hou X,Dewdney M M,et al. Diaporthe species occur in on citrus in China[J]. Fungal Diversity,2013,61(1): 237-250.
[24]林宇,李增平,吳如慧,等.非洲楝擬莖點霉葉斑病病原菌鑒定及其生物學特性的測定[J].熱帶生物學報,2019,10(1):34-40.
[25]Sun W,Huang S,Xia J,et al." Morphological and molecular identification of Diaporthe species in south-western China,with description of eight new species[J]. Mycokeys,2021,77: 65-95.
[26]高雅慧. 間座殼屬真菌的分類及系統演化[D]. 北京:中國科學院大學,2017.
[27]Santos L,Alves A,Alves R. Evaluating multi-locus phylogenies for species boundaries determination in the genus Diaporthe[J]. PeerJ,2017,5: e3120.
[28]賈鑫宇,東保柱,孟煥文,等. 沙棘干腐病菌的鑒定及其生物學特性[J]. 植物病理學報,2023,53(3): 522-526.
[29]年文楷,竇濤,蔡玉彪,等. 一株地黃莖部致病菌的鑒定及防治藥劑篩選[J]. 植物病理學報,2022,52(2):129-137.
(責任編輯:黃雄峰)
