雌性斑馬魚暴露于環境劑量三唑磷對親代生殖和氧化應激及子代胚胎發育的影響
2024-02-07 07:29:42汪和祥李瑞嬌張建祿張春云王立新熊冬梅
水生生物學報 2024年2期
汪和祥 李瑞嬌 張建祿 張春云 王立新 陳 緒 梁 昊 熊冬梅
(1.西北農林科技大學動物科技學院水產科學系, 楊凌 712100; 2.陜西省水產研究與技術推廣總站, 西安 710086;3.陜西省秦嶺生態安全重點實驗室, 陜西省動物研究所, 西安 710032)
三唑磷作為一種高效廣譜有機磷硫代磷酸酯類農藥, 通過抑制乙酰膽堿酯酶活性破壞神經系統功能而達到殺滅害蟲的效果, 在農業生產中被廣泛用作殺蟲殺螨劑, 以提高棉花、水稻、小麥、茶葉、水果和蔬菜等多種作物產量[1—6], 此外, 在漁業上還被用作海水貝類養殖池的清塘劑[7]。據報道2013—2018年, 我國三唑磷年均用量約1萬噸[8]。三唑磷的大量使用造成農產品和環境中的高殘留, 隨著地表徑流、污水排放和雨水沖刷等最終匯入江河或海洋[9,10], 且由于其半衰期較長, 進入水體后短時間內難以降解, 如印度運河中三唑磷半衰期約為25d[11]。我國福建省九龍江中有5個采樣點的三唑磷濃度在100 ng/L以上, 最高濃度為1055 ng/L[10];東海中三唑磷的殘留濃度高達8530.50 ng/L[12]; 溫州蒼南縣某網箱養殖水體中檢測到三唑磷含量為0.011 mg/L[13]。
水體中三唑磷殘留對水生生物造成負面影響已引起學者們的密切關注。有研究表明三唑磷對水生生物具有較高毒性, 可引起多種水生生物中毒、氧化損傷、組織結構破壞、miRNA表達水平變化甚至死亡[7,13,14—16]。三唑磷還可對魚類胚胎和仔魚產生影響, 如誘導稀有鯽(Gobiocypris rarus)胚胎及仔魚畸形率和死亡率增加, 導致其體長和心率下降[17], 造成草魚(Ctenopharyngodon idella)胚胎嚴重畸形和死亡[18]。三唑磷已成為嚴重威脅水生生態系統安全的污染物之一[14,19]。
三唑磷作為神經毒物, 除了造成生物明顯的神經損傷, 還具有環境雌激素效應, 影響受試動物的生殖功能[20]。例如大鼠(Rattus norvegicus)口服三唑磷引起卵泡閉鎖與顆粒細胞凋亡, 可能干擾雌激素/孕激素平衡, 對生育能力有潛在影響[21]; 雌性大鼠孕前口服三唑磷30d后產仔數略有減少, 造成子代皮毛發育顯著延遲、體重下降及子代性比顯著改變, 三唑磷影響雌性大鼠生殖性能[22]。此外, 三唑磷對褐飛虱(Nilaparvata lugens)和灰飛虱(Laodelphax striatellus)的產卵有顯著的刺激作用[23]。以上研究多聚焦于三唑磷的急性毒性試驗, 且所用三唑磷劑量遠高于環境中的實際濃度, 而環境劑量三唑磷長期暴露對魚類生殖毒性鮮有報道。
魚類是水生生態系統中的重要組成部分, 是水生食物鏈中的高級消費者, 對水域生態環境具有重要的指示作用。斑馬魚(Danio rerio)作為模式生物之一, 具有體型小、繁殖周期短、雌雄特征明顯、體外受精和胚胎透明等優勢, 被廣泛用作發育生物學和毒理學研究的受試生物[24,25]。
本研究將3月齡雌性斑馬魚暴露于環境濃度三唑磷21d, 探究其對親代生殖力、子代胚胎發育及親子代氧化應激的影響, 旨在評估三唑磷對淡水生態系統的風險, 積累三唑磷對魚類的亞慢性毒理試驗數據。
1 材料與方法
1.1 實驗試劑
三唑磷(純度98.9%, CAS: 24017-47-8)購于成都艾科試劑公司, 三唑磷母液(107μg/L)以二甲基亞砜(DMSO, 99.5%)為助溶劑進行配制, 并于4℃黑暗條件保存, 暴露期間取母液用除氯自來水配制所需濃度。本試驗所用試劑均為分析純等級。
1.2 實驗魚
三月齡野生型斑馬魚購自中國科學院水生生物研究所(湖北武漢)。實驗魚在正式試驗開始前于(26±1)℃的循環除氯自來水中馴養適應2周, 光暗周期為14h∶10h(明/暗), 每日投喂商品飼料2次, 日投喂量不超過體重的2%。
1.3 毒性試驗與自然繁殖及采樣
在馴養結束后隨機挑選健康的雌性斑馬魚進行暴露試驗, 并記錄初始體重。前文提到三唑磷的半衰期約25d且自然水域檢測到的殘留濃度為8.53 μg/L[11,12], 因而本文選擇環境劑量三唑磷(10 μg/L)進行21d亞慢性暴露, 以0.001% (v/v)DMSO作為溶劑對照組。每個處理組設3個重復, 每個重復9尾魚,每48h更換暴露液總體積的1/2。暴露試驗期間保持與馴養期間相同的喂養條件, 暴露結束后稱量雌魚終末體重(n=3), 同時將全部雌魚與未暴露雄魚置于清水中按1∶1配對, 次日早晨7:00繁殖產卵, 產卵持續2h后收集受精卵并用清水洗滌兩次以備后續試驗。受試雌魚均只進行一次配對產卵。雌魚在產卵結束2h后立即以間氨基苯甲酸乙酯甲磺酸鹽(MS-222, 100 mg/L)進行麻醉, 解剖并取出肝臟, 液氮速凍后存于-80℃用于酶活測定等, 同時取出性腺并稱重, 用于計算性腺成熟系數(GSI;n=27), 然后將未產卵雌魚的性腺固定于波恩氏液。
1.4 胚胎發育觀察
收集受精卵, 統計胚胎受精率。在體視鏡下(Phenix-XTL-3B)用數碼相機(ZXcamera-1600)觀察受精卵并測量卵徑、初孵仔魚體長。在胚胎發育48h后, 于光學顯微鏡(Motic-BA210LED)下測定胚胎心率。仔魚孵化出膜后24h統計孵化率及畸形率。胚胎受精率為每個重復中未受精胚胎數與總胚胎數的比值(n=3)。在統計受精卵直徑時, 從每尾雌魚所產受精卵中選取30枚進行測量, 計算每個重復中受精卵直徑平均值, 初孵仔魚體長以相同的方法進行統計(n=3)。每尾雌魚所產胚胎隨機選取20枚統計心率, 用于計算每個重復的心率均值(n=3)。孵化率為每個重復中孵化胚胎數占受精卵總數的百分比(n=3)。初孵仔魚畸形率為每個重復中畸形個體數占總數的比例(n=3)。
1.5 卵母細胞各時相數量統計
未產卵雌魚卵巢于波恩氏液固定24h后進行乙醇梯度脫水、二甲苯透明和石蠟包埋等, 將包埋塊切至5 μm厚度并進行HE染色, 封片干燥后在光學顯微鏡下觀察并統計第1—5時相卵母細胞的數量,卵母細胞時相劃分采用劉敏等[26]方法。對照組和處理組各隨機選取6尾雌魚制作性腺切片(n=6), 每張切片計數視野不少于10個。
1.6 生化指標測定
測定乙酰膽堿酯酶(AChE)、丙二醛(MDA)含量、超氧化物歧化酶(SOD)、谷胱甘肽過氧化物酶(GPx)、谷胱甘肽-S-轉移酶(GST)及羧酸酯酶(CarE)的活性來評估三唑磷處理后對雌魚及其子代氧化損傷和解毒酶活性。生化指標測定試劑盒均購自南京建成生物工程研究所。分別從每個重復隨機選取3尾雌魚的肝臟(親代)和20尾初孵仔魚(子代)進行混樣(n=3), 樣品按1∶9(重量體積比)加入生理鹽水并磨成勻漿, 參照試劑盒說明書測定以上指標。
1.7 統計分析
采用卡方檢驗比較已產和未產雌魚數占樣本總數比例的差異顯著性; 本試驗數據均采用均值±標準差(mean±SD)表示, 并使用SPSS 23.0軟件以獨立樣本T檢驗進行統計分析,P<0.05為差異顯著。
2 結果
2.1 雌魚體重變化與性腺成熟系數
為評估三唑磷暴露21d后對雌魚生長的影響,測量了暴露期間雌魚體重增長量以及暴露后雌魚的性腺成熟系數(GSI)。對照組雌魚平均增重0.072 g,處理組雌魚平均增重0.070 g, 兩組無顯著差異(P>0.05;圖1A)。對照組雌魚GSI為22.24%, 處理組雌魚GSI為23.37%, 與對照組相比差異不顯著(P>0.05;圖1B)。
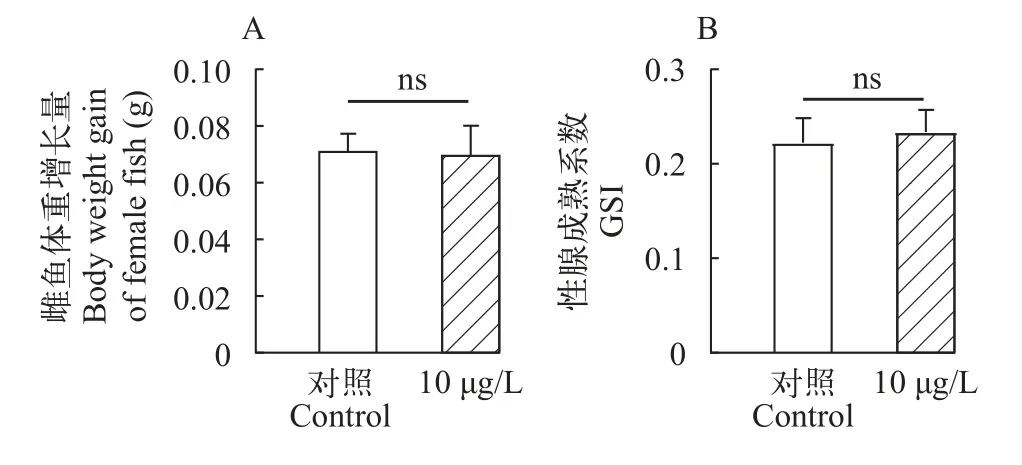
圖1 三唑磷暴露21d后雌魚體重增長量和GSIFig.1 Body weight gain and GSI in female fish after 21d of exposure to triazophos
2.2 產卵雌魚數及其產卵量
如圖2所示, 對照組27尾雌魚中產卵14尾, 未產卵13尾; 三唑磷處理組的產卵雌魚數為20尾, 7尾未產卵; 卡方檢驗結果顯示, 處理組與對照組間無顯著差異(P>0.05; 圖2A)。對照組雌魚平均產卵數為219.7粒, 而處理組雌魚產卵數顯著增加至250.2粒(P<0.05; 圖2B)。
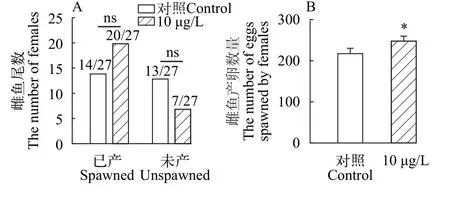
圖2 三唑磷暴露21d后產卵雌魚數及其對應的產卵量Fig.2 Number of spawning females and their corresponding spawning numbers after 21d of triazophos exposure
2.3 卵巢組織切片各時相卵母細胞數量
為探討三唑磷暴露對親代雌魚卵巢發育是否產生影響, 制作卵巢組織切片并統計各時相卵母細胞數量(圖3)。經三唑磷暴露21d后, 處理組第1時相與第2時相卵母細胞數量與對照組相比差異均不顯著(P>0.05)。處理組第3時相的平均卵母細胞數為5.7個, 顯著低于對照組(7.5個;P<0.05)。對照組第4時相的平均卵母細胞數為2.8個, 處理組為3.1個,差異不顯著(P>0.05)。對照組和處理組中第5時相卵母細胞平均數量分別為10和10.7個, 與對照組相比, 處理組第5時相卵母細胞數量顯著增加(P<0.05)。
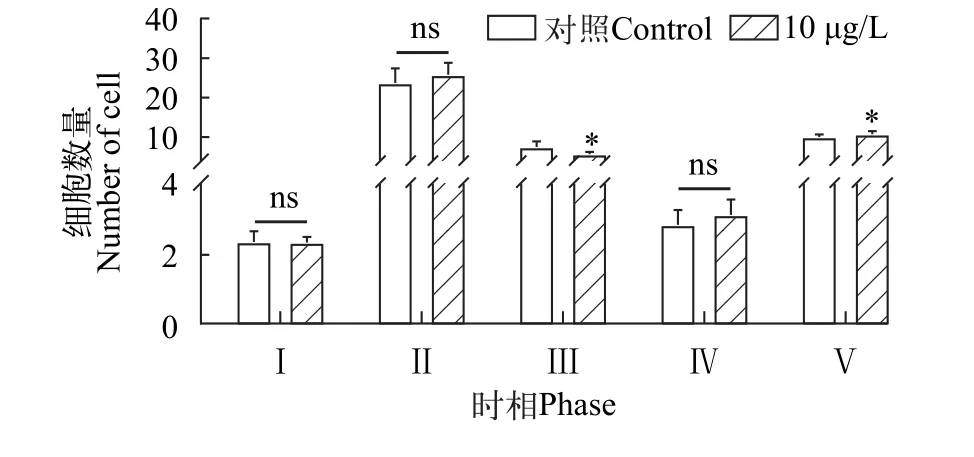
圖3 三唑磷暴露21d后雌魚卵巢組織切片中Ⅰ—Ⅴ時相卵母細胞數量Fig.3 Number of oocytes in Ⅰ—Ⅴ phase in ovarian tissue sections of female fish after 21d of triazophos exposure
2.4 生化指標
AChE 活性如圖4所示, 與對照組相比, 處理組雌魚肝臟中AChE活性顯著降低(P<0.05; 圖4)。對于雌魚所產仔魚的AChE活力, 三唑磷處理組AChE活力較對照組有下降趨勢, 但差異不顯著(P>0.05; 圖4)。
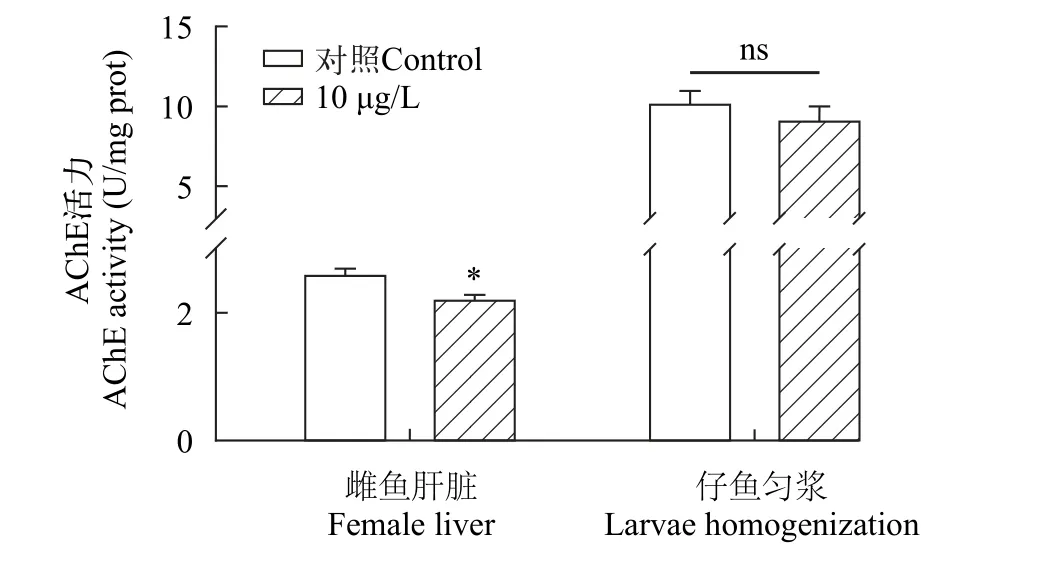
圖4 三唑磷暴露21d后雌魚肝臟及其所產仔魚AChE活性Fig.4 AChE activity in liver of female fish and their offspring after 21d of triazophos exposure
抗氧化指標通過測定SOD、GPx活性和MDA含量來評估三唑磷暴露對雌魚及其子代造成的氧化損傷。三唑磷暴露21d后雌魚肝臟SOD活性顯著升高(P<0.05), 但其子代SOD活性與對照組相比無顯著差異(P>0.05; 圖5A)。處理組雌魚肝臟中MDA含量顯著增加(P<0.05), 在其子代中也發現了類似結果(P<0.05; 圖5B)。與對照組相比, 經三唑磷處理的雌魚肝臟GPx活力顯著增加(P<0.05), 而其子代GPx活力則顯著降低(P<0.05; 圖5C)。
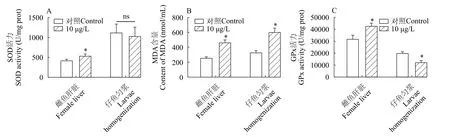
圖5 三唑磷暴露21d后雌魚肝臟及其所產仔魚部分抗氧化指標的變化Fig.5 Changes of some antioxidant indices in liver of female fish and their offspring after 21d of triazophos exposure
解毒酶活性GST和CarE的活性被用來評估雌魚及其子代對三唑磷的解毒能力。與對照組相比, 處理組雌魚肝臟中檢測到GST活性顯著升高(P<0.05), 然而其仔魚中GST活性顯著降低(P<0.05;圖6A)。與對照組相比, 在三唑磷暴露后, 雌魚及其仔魚CarE活性顯著提高(P<0.05; 圖6B)。
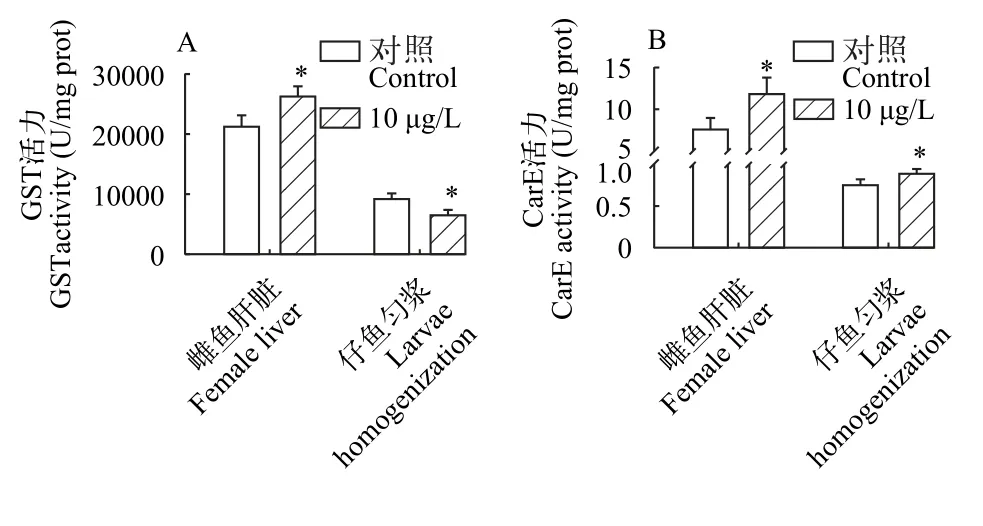
圖6 三唑磷暴露21d后雌魚肝臟及其所產仔魚的解毒酶活性Fig.6 Detoxification enzyme activity in the liver of female fish and their offspring after 21d of triazophos exposure
2.5 胚胎發育相關指標
檢測子代胚胎心率、仔魚孵化率和畸形率等作為評估三唑磷暴露雌魚后對所產子代的發育毒性指標。與對照組相比, 三唑磷處理后雌魚所產胚胎受精率顯著降低(P<0.05; 表1), 受精卵直徑顯著減小(P<0.05; 表1)。對照組胚胎平均心率為110.1次/min, 而處理組胚胎平均心率顯著下降(103.2次/min,P<0.05; 表1)。處理組胚胎孵化率與對照組相比無差異(P>0.05; 表1)。與對照組相比, 處理組初孵仔魚體長顯著縮短(P<0.05; 表1), 仔魚畸形率高于對照組7.2倍(P<0.05; 表1), 處理組仔魚畸形如圖7,主要表現為脊柱彎曲及卵黃囊腫大。
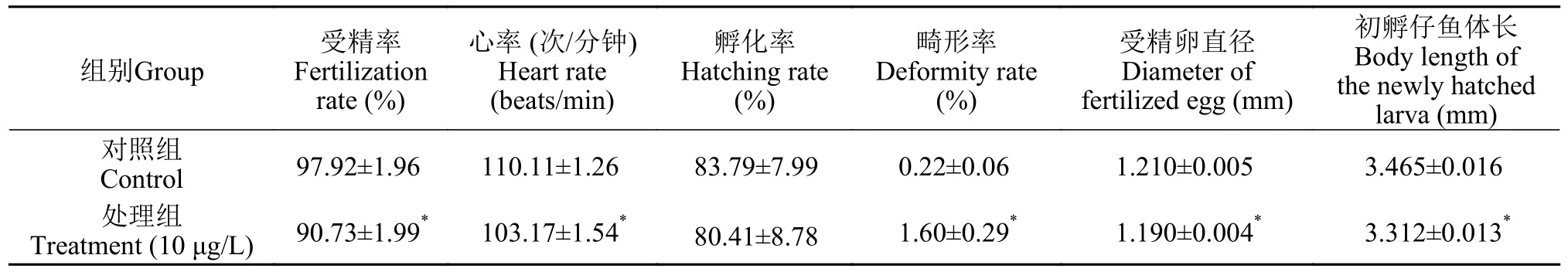
表1 雌魚所產子代胚胎發育相關指標Tab.1 Related indicators of embryo development of offspring produced by female fish

圖7 雌魚暴露于三唑磷21d后所產仔魚畸形圖Fig.7 Malformation of larvae when their parent female exposed to triazophos for 21d
3 討論
3.1 三唑磷暴露對受試動物生殖的影響
本研究中環境濃度三唑磷暴露21d后雌魚體重增長量與性腺成熟系數(GSI)無顯著變化, 但雌魚產卵量顯著增加(圖2B)。通過觀察卵巢組織切片來進一步探討雌魚產卵量增加是否與卵巢中成熟期卵母細胞的數量有關, 結果顯示處理組成熟期卵母細胞數量與對照組相比顯著增加(圖3)。在三唑磷的靶標生物上發現類似的結果, 如三唑磷暴露對三種飛虱產卵有顯著的刺激作用[23], 導致褐飛虱的繁殖力增強[27]。但也有研究表明三唑磷降低受試動物的生殖力, 如雌性大鼠孕前口服三唑磷30d后產仔數略有降低, 幼崽在30d發育過程中體重減輕[22]。本研究表明環境相關濃度三唑磷暴露增強雌魚的產卵量, 這與飛虱上的研究結論相似, 卻與大鼠的研究結果相反, 這可能是物種差異導致。
3.2 三唑磷暴露對AChE、氧化應激和解毒相關生化指標的影響
乙酰膽堿酯酶(AChE)是以三唑磷為代表的有機磷農藥的作用靶酶。三唑磷通過抑制中樞和周圍神經系統中的AChE來發揮其作用[28]。據報道,三唑磷暴露可抑制鯉(Cyprinuscarpio)腦內AChE 活性, 且抑制作用會隨著暴露濃度的升高而增強[29];三唑磷對金魚(Carassius auratus)腦、脾、腎和肝臟組織中AChE活性的抑制作用隨濃度的升高逐漸增強[16];三唑磷暴露雄性翡翠貽貝(Pernaviridis)15d后, 鰓中AChE活力顯著降低[30];稀有鯽胚胎和仔魚體內AChE活性對三唑磷的響應呈低劑量促進高劑量抑制的模式[17]。本研究中三唑磷暴露21d后雌魚肝臟AChE活力顯著降低, 仔魚AChE活力也出現下降趨勢(圖4), 表明三唑磷對雌魚及所產仔魚造成了不同程度的神經毒性。
MDA常被用作評估水生生物在毒物暴露下氧化應激狀態的有效生物標志物,以反映細胞和組織的損傷水平[31]。本研究中三唑磷暴露導致雌魚肝臟及其所產仔魚MDA含量顯著增加(圖5B), 表明雌魚肝臟及其所產仔魚均遭受氧化損傷。生物體內存在多種機制保護機體免受外來物質造成的應激, 如抗氧化物。SOD-CAT系統提供了抵抗氧化應激的第一道防線,其中SOD催化超氧陰離子自由基()和H+分別轉化為無毒的H2O2和O2,CAT則將H2O2分別轉化為無毒的H2O和O2[32,33]。在抗氧化過程中,GPx也發揮了關鍵作用,其主要以過氧化物為底物,將其催化還原為O2和H2O[34]。本研究結果顯示三唑磷暴露后雌魚肝臟SOD和GPx活性顯著提高(圖5A和5C), 提示機體抗氧化系統產生了應答, 從而保護機體免受氧化損傷。前人也報道過類似結論, 經有機磷農藥毒死蜱暴露后凡納濱對蝦(Litopenaeus vannamei)體內MDA含量上升, GPx活性顯著提高[35], 高濃度毒死蜱誘導翠鱧(Channa punctata)體內MDA增加, SOD和GPx活力增強[36]。值得注意的是, 本文在仔魚體內卻觀察到GPx活性顯著下降, SOD活性也呈下降趨勢(圖5A和5C)。據報道, 三唑磷對稀有鯽胚胎和仔魚酶活性的作用特征是低劑量刺激轉變為高劑量抑制[17]。在本試驗中仔魚酶活性變化與雌魚相反可能是由于仔魚對三唑磷更為敏感, 三唑磷暴露親代雌魚導致所產仔魚抗氧化系統受損, 氧化平衡被打破。另外,研究指出三唑磷暴露會造成金魚體內的組織特異性抗氧化反應[16], 本研究測定抗氧化指標時, 雌魚選取肝臟進行測定, 仔魚由于肝組織精準取樣困難則是全魚勻漿, 出現差異的原因也可能與組織特異性抗氧化反應有關。
生物體內存在多種解毒酶以維持機體正常生命活動, GST作為一種生物轉化酶, 可催化谷胱甘肽(GSH)與多種親電物質或異種生物偶聯反應, 具有抗外來生物的活性, 從而保護組織免受氧化應激[37—39]。CarE是生物體內重要的解毒酶和代謝酶,它以水解和結合蛋白兩種方式對殺蟲劑解毒, 尤其對有機磷類殺蟲劑的代謝作用明顯[40]。在本研究中, 三唑磷暴露后雌魚肝臟GST和CarE活性顯著提高(圖6A和6B), 表明機體對三唑磷造成的毒性產生了響應, 解毒酶積極參與了對三唑磷的解毒過程。在仔魚中同樣觀察到CarE活性提高(圖6B), 表明仔魚對三唑磷的解毒也產生了響應; 與之相反,仔魚GST活性下降(圖6A)。前人研究表明三唑磷對稀有鯽胚胎和仔魚GST活性影響呈低濃度促進、高濃度抑制的趨勢[17], 在本研究中仔魚GST活性下降可能的原因是仔魚對于三唑磷更為敏感, 較低劑量的環境濃度三唑磷暴露親代雌魚抑制仔魚GST活性。此外, 研究表明, 一些魚類對外來物的反應過程中GST活性變化與GSH含量密切相關[41],如一種鮰(Ictalurus melas)汞暴露后觀察到GSH含量增加, 但GST活性降低[42]。本研究中三唑磷暴露后雌魚及所產子代GST活性變化的原因可能與GSH含量有關, 這需要進一步研究確定。
3.3 毒物暴露對胚胎發育的影響
許多研究證明毒物會導致受試動物發育異常。據報道, 大鼠口服三唑磷后其子代觀察到步態受損、前后肢癱瘓和發育過程中體重減輕等[22]。兩種殺蟲劑暴露21d后導致大型溞(Daphnia magna)體長縮短[43]。有機汞暴露導致斑馬魚仔魚眼睛發育異常、脊柱畸形和尾鰭彎曲[44], 汞短期暴露胚胎導致牙鲆(Paralichthys olivaceus)仔魚出現各種畸形[31]。高濃度三唑磷暴露草魚胚胎導致受精卵嚴重畸形和死亡、仔魚無法正常生存[18]。三種六色環十二烷暴露會造成斑馬魚胚胎心率降低, 導致仔魚畸形率和死亡率增加[45]。在三唑磷72h急性暴露后, 稀有鯽胚胎及仔魚死亡率和畸形率隨暴露濃度的增加而增加, 體長和心率隨暴露濃度的增加而逐漸降低[17]。在本試驗中, 經三唑磷暴露的雌魚所產子代中受精卵直徑減小、胚胎心率降低、初孵仔魚體長縮短及仔魚畸形率上升(表1), 這與前人探究毒物對胚胎和仔魚毒性的研究結果相似[17,18,31,44,45]。本研究還觀察到雌性斑馬魚經環境相關濃度的三唑磷暴露后胚胎受精率降低(表1),表明對其所產子代也造成明顯毒性。在本研究中,環境濃度三唑磷暴露親代雌魚造成子代的氧化應激, 推測子代胚胎發育不良可能是三唑磷誘導氧化應激造成的, 這需要進一步研究證實。此外, 本研究中三唑磷暴露后雌魚體重與GSI并無顯著變化,但雌魚產卵量卻顯著提高, 值得注意的是, 子代受精卵直徑與初孵仔魚體長顯著減小, 可能原因是當親代雌魚在應對污染物暴露脅迫的過程中, 通過增加子代數量及幼仔個體變小來維持穩定的種群數量。
4 結論
綜上所述, 本研究表明環境相關濃度三唑磷暴露親代雌性斑馬魚可提高雌魚的產卵量, 抑制雌魚肝臟及所產仔魚體內AChE活性, 誘導雌魚肝臟及其所產仔魚氧化應激并提高雌魚和仔魚體內解毒酶活性。此外, 親代雌性斑馬魚暴露于三唑磷導致其所產子代胚胎畸形、個體變小和心跳變緩等不良反應。本研究結果積累了三唑磷對魚類的生殖毒性的基礎數據, 可為合理評估三唑磷對淡水生態系統風險提供參考依據。
