鹽度和pH對平凡舟形藻生長和油脂積累的影響
2024-02-07 07:29:46信宏穎梁金燕王馨彤范亞文陸欣鑫隋豐陽
水生生物學報 2024年2期
關鍵詞:生長
信宏穎 梁金燕 王馨彤 劉 妍, 范亞文, 陸欣鑫, 隋豐陽,
(1.哈爾濱師范大學生命科學與技術學院, 哈爾濱 150025; 2.黑龍江省水生生物多樣性研究重點實驗室, 哈爾濱 150025)
利用可再生資源生產生物燃料是進行可持續發展的選擇[1]。微藻在工業上的應用已經引起了人們的關注, 但由于微藻對環境變化敏感, 進一步影響微藻產油效率[2]。大多數微藻具有將大氣中的二氧化碳通過光合作用轉化為生物質、脂肪酸和脂類等物質儲存在細胞內[3], 不僅使微藻在生產生物燃料方面具有開發潛力, 在生產高價值脂類化合物方面具有更高的應用價值[4]。微藻的總脂質含量約為干細胞重量的1%—70%, 油脂主要以甘油三酯和脂肪酸甲酯的形式存在, 適合生產生物柴油[5,6]。據報道, 許多微藻物種細胞中含有大量的脂質, 它們的脂質可以很容易地通過酯交換過程轉化為生物柴油, 因此表明了它們作為未來生物能源替代來源的重要性[7]。微藻分布范圍廣, 可以在多種水生生境中正常生長, 包括湖泊、池塘、河流和海洋,甚至廢水, 也可以在人為調控的不同條件下培養[8]。微藻具有廣闊的應用前景, 但目前微藻規模化生產中仍存在產量低和成本高的問題。微藻在培養過程中還需要嚴格控制各種環境因素, 如碳源[9]、鹽度[10]、氮[11]、溫度[12]、CO2濃度[13]和光照強度[14]等。少量的微量營養素, 如鐵[15]和鋅[16]等也是影響藻類生長和代謝的重要因素, 微藻利用各種機制來適應環境脅迫, 例如改變其形態和物質組成[17]。因此, 生產中可以使用不同應激源來提高某些藻類中色素和脂類成分。其中鹽度和pH是本研究的重點影響因素。
鹽度是影響微藻生長、脂肪含量和生化組成的重要因素之一。由于藻種的生理差異, 微藻對鹽度的適應性也不同[18]。在微藻生長過程中, 鹽脅迫會影響類胡蘿卜素合成和脂肪積累, 并影響脂肪酸組成[19]。有報道稱,Botryococcus braunii可以有效地將鹽排除在細胞外, 并在生長過程中積累甘油,作為對胞外鹽濃度的直接反應[20]。通過增加鹽度來脅迫藻類細胞, 可能會導致額外的脂肪積累[21]。在微藻生長過程中, 介質中的pH也起著重要的作用, 因為其影響二氧化碳濃度, 會導致物種的分布和碳的利用率不同。據報道, 萊茵衣藻的細胞器和其他內部過程需在胞液pH 7—7.1才能發揮作用[22],在酸性的外部條件下, 額外的ATP被消耗以維持胞液pH為7, 導致萊茵衣藻在酸性環境中的能效較低。當多余的ATP被耗盡時, 用于細胞生長和繁殖的能量就會減少, 在堿性pH中觀察到相反的反應。此外, 葉綠素含量也受到pH的影響, 因為pH決定了光合作用過程[23]。因此, 研究鹽和pH脅迫對藻株的生長和生化的影響尤為重要。
平凡舟形藻(Navicula trivialis)是本實驗室分離的一株生長快的淡水底棲硅藻, 該藻隸屬于硅藻門、硅藻綱、舟形藻目、舟形藻科、舟形藻屬, 經實驗室分離培養初步得到油脂含量較高的藻株, 有關該藻的生態習性及培養方法迄今為止鮮有報道。利用微藻生產生物柴油被認為是解決資源消耗、能源短缺和環境污染等問題的一種很有前途的策略[9]。因此, 藻類生物量和脂質生產力的優化是藻類生產生物燃料研究的主要重點。本研究將平凡舟形藻在不同NaCl濃度和pH條件下進行培養, 分析該藻株的最佳培養條件和油脂產量, 本研究的結果可能有助于了解不同的鹽度和pH如何提高微藻生長和油脂產量, 為進一步提高其油脂的產量奠定基礎。
1 材料與方法
1.1 藻種培養
本研究所用平凡舟形藻藻種由哈爾濱師范大學水生生物實驗室采集并分離純化, 采用水滴分離法, 用移液管和顯微鏡對微藻細胞進行多次分離,得到純化的平凡舟形藻, 并將平凡舟形藻藻種轉接至WC培養基中進行培養, WC培養基如表1所示。挑選生長良好的純種藻種放入光照培養箱中進行逐級擴大培養, 培養溫度為(25±1)℃, 光照條件為4000 lx, 光暗比時間為12h∶12h, 藻液每天定時搖動3—5次, 防止藻細胞附著在瓶壁上, 培養周期12d,達到對數生長期, 用于后續實驗。
1.2 鹽和pH脅迫實驗
將平凡舟形藻培養至對數生長期進行實驗,鹽和pH的 初始接種藻種密度分別為3×105和1×105cell/mL。采用不同濃度的鹽度和pH對平凡舟形藻分別進行單一鹽和pH的脅迫處理。鹽脅迫濃度分別為0、0.03、0.06、0.12、0.18和0.24 mol/L。用1.00 mol/L HCl和NaOH調節培養基pH至4.5、6.0、7.5、8.5、9.5和10.5。在相同的實驗條件下, 每個處理重復3次, 每天搖瓶5次, 將平凡舟形藻放入WC培養基中, 置于(25±1)℃的恒溫培養箱中培養, 光照強度4000 lx, 光暗比為12h∶12h, 分別在第0、第2、第4、第6、第8、第10和第12天時取樣進行相關指標的測定。
1.3 測定指標及方法
細胞密度依照設定的鹽濃度和pH脅迫后,采用分光光度法和血球計數法相結合的方法對平凡舟形藻細胞數量進行統計。得到細胞密度和透光率對應的標準曲線。取各處理組2 mL硅藻懸浮液, 在680 nm波長處測定透光率(以WC培養液為空白組)。根據標準曲線計算細胞數, 各處理組重復3次, 記錄12天內平凡舟形藻細胞數量的變化。
葉綠素a含量采用丙酮萃取法測定藻細胞中的葉綠素a含量[24]。取不同鹽濃度和pH脅迫后的藻液10 mL, 離心棄上清液, 使用5 mL 80%的丙酮溶液進行超聲提取(40 kHz, 30min)。樣品在4℃下提取24h, 在646和663 nm波長處測定吸光度(以80%的丙酮進行調零), 葉綠素a含量由以下公式計算:
式中,A663為663 nm處的吸光度;A646為646 nm處的吸光度。
葉綠素熒光參數取5 mL平凡舟形藻的藻液在暗處理下適應30min后, 使用雅欣葉綠素熒光儀(Yaxin-1161)測定平凡舟形藻相關葉綠素熒光參數[25]。
總脂含量使用Ryckebosch[26]的方法將培養12天的微藻離心, 棄上清(m總), 藻液與4 mL甲醇混合。加入氯仿2 mL和蒸餾水0.4 mL, 渦旋震蕩混合2min, 再加入氯仿和蒸餾水各2 mL, 渦旋震蕩混合。以2000 r/min的速度離心10min, 棄上層, 將下層轉移到1個離心管中。剩余固體用4 mL氯仿-甲醇1∶1反復萃取3次。使用旋轉蒸發儀進行干燥,稱取脂肪收集瓶和脂肪總質量(m1),用旋轉蒸發儀在40℃下旋轉蒸發除去溶劑(m2), 然后用重量法測定脂肪含量。每個處理重復3次, 用以下公式進行計算:
式中,X為樣品中脂肪含量;m1為脂肪收集瓶和脂肪總質量;m2為脂肪收集瓶質量;m總為樣品的質量。
數據處理所有處理3次重復, 數據分析都在IBM SPSS 26.0軟件中進行統計分析, 多重比較采用LSD法進行, 差異顯著性水平為0.05 (P<0.05)。
2 結果
2.1 鹽濃度和pH對平凡舟形藻生長的影響
鹽濃度的改變對平凡舟形藻的生長有顯著性差異。圖1A顯示了平凡舟形藻經不同鹽濃度脅迫培養12d后的生長情況, 平凡舟形藻的生長在各處理組不同水平間表現出明顯的差異。平凡舟形藻細胞密度在鹽濃度0—0.12 mol/L時隨著濃度的升高而增加, 在鹽度為0.12 mol/L細胞密度增長最快,第6天細胞密度顯著高于對照組(P<0.05), 后隨著鹽濃度的繼續升高會顯著降低。培養時間在第8天,鹽度為0.12 mol/L時, 細胞密度達到最大值, 為5.5×105cell/mL。在鹽濃度為0.03和0.06 mol/L時,生長呈緩慢上升的趨勢, 細胞密度最高可達4.3×105和4.9×105cell/mL。在鹽度為0.24 mol/L時,平凡舟形藻的細胞密度隨著培養時間的增長一直呈現下降趨勢, 說明鹽度過高不利于平凡舟形藻的細胞密度的增加。
在本研究中, 平凡舟形藻能夠在所測試的大多數pH(6.0、7.5和8.5)條件下生長(圖1B), 其中pH為7.5在第10天時平凡舟形藻細胞密度增加到最大值為1.4×106cell/mL, 細胞密度相較于其他處理水平有顯著增加(P<0.05), 其次細胞密度增長最快的是pH為8.5時。過酸或過堿的環境會導致平凡舟形藻的生長減慢, 如在pH為4.5、9.5和10.5細胞密度快速下降, 最高值分別為3.3×105、8.6×105和6.6×105cell/mL, 細胞密度比pH 7.5時低, 所以平凡舟形藻最適生長的pH為7.5。
2.2 鹽濃度和pH對平凡舟形藻葉綠素a含量的影響
在不同的鹽濃度下, 葉綠素a含量隨鹽度變化而變化。如圖2A顯示, 在6種鹽濃度下的平凡舟形藻葉綠素a的含量呈現先升后降的趨勢。在0.03和0.06 mol/L從第2天葉綠素a含量開始增加, 在第8天逐漸下降。當鹽濃度為0.12 mol/L時, 葉綠素a含量增長最快, 與對照組相比有顯著增加(P<0.05), 當鹽濃度增長到0.18和0.24 mol/L時, 隨著培養時間的增加, 葉綠素a含量積累緩慢, 處于較低水平。結果表明: 低濃度的鹽可以促進葉綠素a含量的增加, 當鹽濃度超過平凡舟形藻耐受的臨界值, 會抑制葉綠素a含量增加。

圖2 鹽度(A) 和pH (B)處理下平凡舟形藻葉綠素a含量Fig.2 Effect of salinity (A) and pH (B) on the chla content of Navicula trivialis
葉綠素a含量在不同pH處理條件下有顯著性差異(圖2B), 平凡舟形藻的葉綠素a含量最高值出現在pH為7.5時為1.48 mg/L。隨著pH的增加或減少,葉綠素a的含量顯著下降(P<0.05)。當pH為4.5、6.0、8.5、9.5和10.5時都抑制了葉綠素a的積累。當pH在堿性條件下, 如8.5、9.5和10.5, 葉綠素a含量相較于pH 7.5時顯著下降(P<0.05), 分別為1.28、0.89和0.8 mg/L時, 相對于pH 7.5降低了1.15、1.66和1.83倍。當pH在酸性條件下, pH為4.5和6.0葉綠素a含量分別為0.83和1.27 mg/L, 呈下降趨勢。所以pH為7.5更有利于平凡舟形藻葉綠素a含量的積累。
2.3 鹽濃度和pH對平凡舟形藻葉綠素熒光參數的影響
葉綠素熒光參數可以很好的反映出微藻細胞光合系統的生理狀態, 在研究不同環境條件下的光合效率具有高度敏感性和可靠性[27]。圖3A和3B顯示不同鹽度處理水平下的葉綠素熒光參數PSⅡ的最大光能轉化效率(Fv/Fm)和PSⅡ的潛在活性(Fv/Fo)的變化趨勢, 二者趨勢基本相同。在0—6d,各鹽度處理組的葉綠素熒光參數Fv/Fm和Fv/Fo不同, 但整體呈現上升趨勢。鹽度為0—0.12 mol/L時, 平凡舟形藻的Fv/Fm和Fv/Fo顯著高于鹽濃度0.18和0.24 mol/L組(P<0.05), 且在鹽度為0.12 mol/L時Fv/Fm和Fv/Fo的值最高。在培養8d后, 6組鹽濃度處理下的葉綠素熒光參數開始呈下降趨勢。結果表明, 高鹽度和低鹽度對平凡舟形藻的光合作用能力均有影響, 低濃度時平凡舟形藻的最大熒光比保持相對穩定。鹽度為0.12 mol/L時, 是最適促進平凡舟形藻的光合效率的鹽濃度。
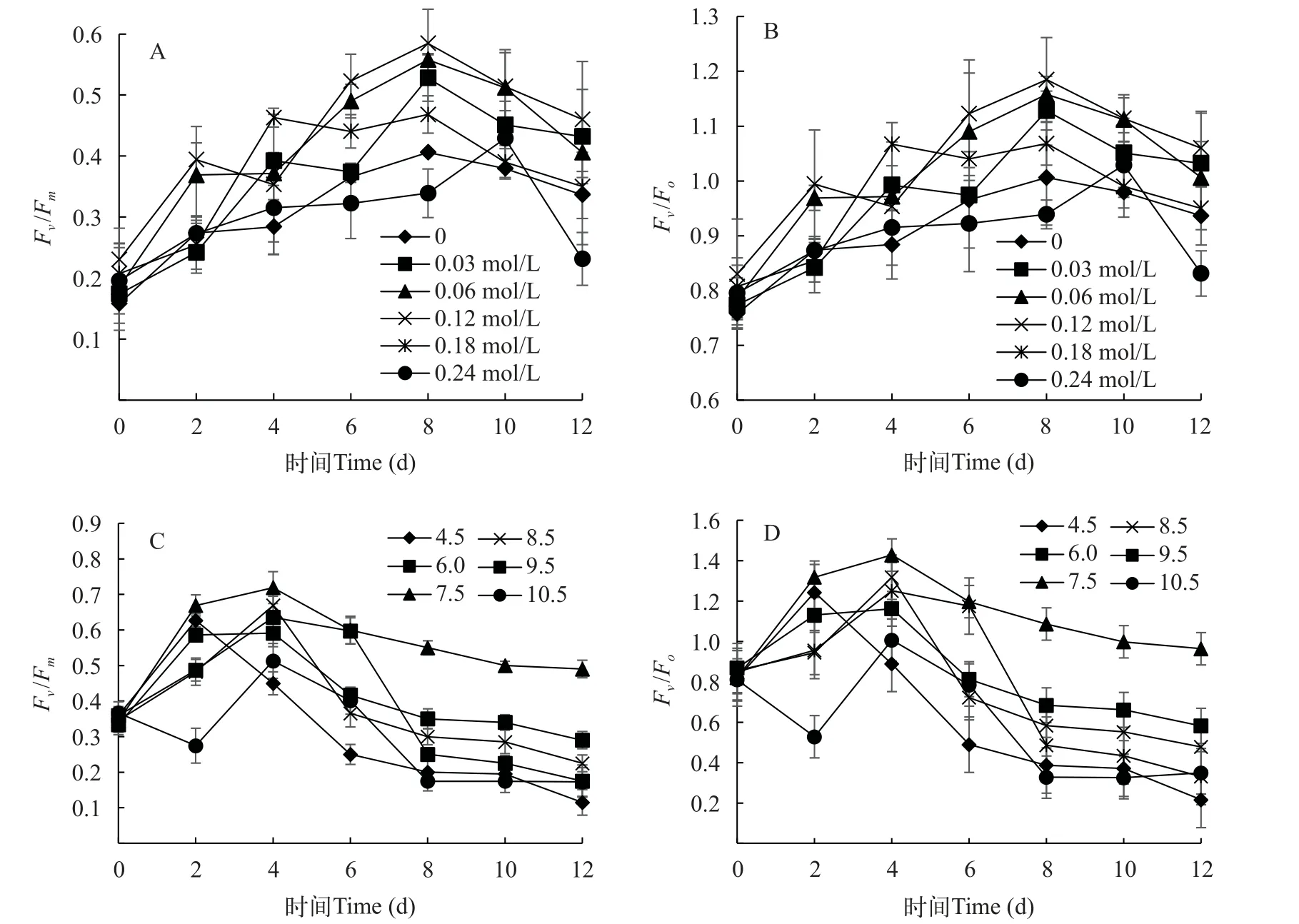
圖3 不同鹽濃度和pH下平凡舟形藻的葉綠素熒光參數Fv/Fm、Fv/FoFig.3 The effects of different salinities (A, B) and pH (C, D) on the Fv/Fm and Fv/Fo of chlorophyⅡ fluorescence parameters of Navicula trivialis
由圖3C和3D可知, 平凡舟形藻葉綠素熒光參數Fv/Fm和Fv/Fo在pH為4.5—10.5均呈現相似的趨勢, 都是先升高后降低, 在0—4d內,Fv/Fm和Fv/Fo葉綠素熒光參數數值迅速升高, 在pH為7.5時Fv/Fm和Fv/Fo葉綠素熒光參數值與其他處理水平有顯著差異(P<0.05), 隨著時間的增加, 在4d之后, 各處理組均呈現下降趨勢, pH為7.5的Fv/Fm和Fv/Fo值高于其余pH。尤其是pH為4.5和10.5時,Fv/Fm和Fv/Fo參數值呈現明顯的下降趨勢, 這可能是由于光合作用對pH的變化敏感, 一旦超過藻類最適pH, 光合速率將會下降。
2.4 鹽濃度和pH對平凡舟形藻總脂含量的影響
鹽度脅迫在誘導藻類脂肪積累過程中起重要作用, 圖4A顯示了不同鹽濃度脅迫下平凡舟形藻的總脂含量, 可以看出隨著鹽濃度的增加, 當氯化鈉濃度從0 增加到0.24 mol/L時, 含油量總體呈上升趨勢。對照組總脂含量為13.8%, 0.03和0.06 mol/L的總脂含量增加, 分別為17.2%和23.4%。鹽濃度為0.12 mol/L的總脂含量與0.06 mol/L相比油脂含量增加, 當鹽濃度增加到0.18和0.24 mol/L時, 總脂含量持續增加, 分別為30.3%和34.9%, 與其他鹽處理組有顯著性差異(P<0.05), 鹽濃度0.24 mol/L的總脂含量比鹽濃度0.18 mol/L高了4.7%。
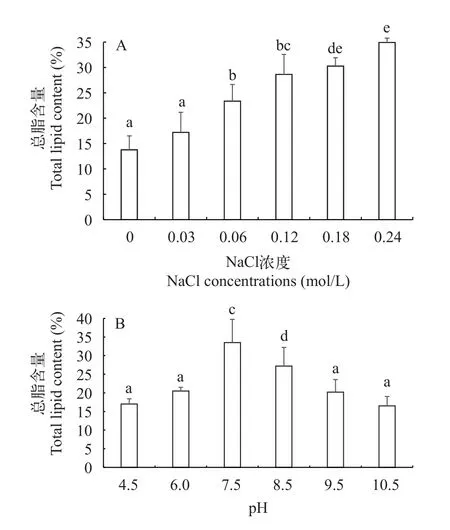
圖4 不同鹽濃度和pH下平凡舟形藻的總脂含量Fig.4 Effect of of salinity (A) and pH (B) total lipid content of Navicula trivialis
圖4B顯示了不同pH脅迫下平凡舟形藻的總脂含量, 可以看出不同的pH對總脂含量的影響不同。以pH為7.5的總脂含量最高, 為33.5%, 與其他pH處理組有顯著性差異(P<0.05)。pH 8.5次之, 為27.2%。酸性和堿性pH并不利于總脂含量的積累, 如4.5、9.5和10.5, 總脂含量分別為17.0%、20.2%和16.5%,三者pH的總脂含量差異不顯著(P>0.05)。pH為6.0時, 總脂含量達到20.5%, 由于在pH為7.5條件下的平凡舟形藻細胞密度和總脂含量最高, 因此確定了培養平凡舟形藻的最佳pH為7.5。
3 討論
鹽度和pH是影響微藻脂質合成的重要因素, 鹽度的升高和pH的改變有利于誘導飽和脂肪酸的合成[28]。由于鹽脅迫是大規模的, 如果能夠發現一種在鹽脅迫下可以積累脂肪的藻株, 這將為生物柴油的生產提供新的途徑。
3.1 鹽度和pH對平凡舟形藻生長的影響
鹽濃度的不同會改變微藻的生長速度和組成[29]。當微藻細胞暴露在鹽脅迫下時, 會出現特殊的適應機制, 例如細胞從活躍的分裂轉變為儲存能量, 積累滲透保護溶質以及保護生物化合物, 通過調節細胞膜的離子交換過程, 恢復生理生化過程[30,31]。pH決定了二氧化碳和必要營養物質的溶解度和可用性, 可以對藻類的新陳代謝產生重大影響[32]。較高的pH限制了二氧化碳中碳的可獲得性, 而二氧化碳反過來又抑制了藻類的生長[32,33]。當pH較高時, 藻類的碳以碳酸鹽的形式存在。較高的pH也會降低藻類對游離二氧化碳的親和力[33,34]。與堿性pH類似, 酸性條件會改變營養物質的吸收[35]或引起金屬毒性[36], 從而影響藻類的生長, 大多數藻類在中性pH為7.0—7.6附近能有最大限度的生長。改變鹽度和pH相對來說操作簡便, 有利于微藻生產生物柴油的規模化, 以增加更多的脂肪產量, 從而降低生物柴油的生產成本。
本研究表明: 在鹽濃度0上升到0.24 mol/L這個區間時, 鹽濃度為0—0.12 mol/L, 對平凡舟形藻細胞密度和葉綠素a含量起促進作用, 當鹽濃度為0.12 mol/L時, 平凡舟形藻的細胞密度和葉綠素a含量達到最大值。Salama等[37]報道稱, 在低鹽濃度對微藻生長速度有促進作用。平凡舟形藻隨著鹽濃度的逐漸增加到0.24 mol/L時, 細胞密度和葉綠素a含量逐漸降低。據Markou等[38]報道隨著鹽濃度的增加, 小球藻YH703的干重和細胞密度顯著降低。細胞密度和葉綠素a隨著時間的推移而降低后, 可能是因為在營養耗竭后達到穩定階段, 微藻細胞密度下降, 生長速度下降, 證明高鹽度會抑制微藻的生長。這或許是由于低濃度鈉離子提高了細胞離子交換能力, 加快了藻類新陳代謝, 從而促進了藻細胞的生長, 而在高鹽度條件下, 藻細胞需要更多的能力來維持正常的滲透壓和離子濃度, 用于生長的能量減少[39]。葉綠素熒光參數Fv/Fm和Fv/Fo在鹽濃度為0—0.12 mol/L時, 平凡舟形藻的光合效率高,隨著鹽濃度的上升, 光合效率大幅度降低, Feng等[40]發現鹽度過高或者過低都會影響梅尼小環藻的光合能力和PSⅡ活性。這表明在適宜的鹽度下微藻的葉綠素熒光參數呈現增加趨勢。藻細胞生長較慢, 因為鹽度過低影響藻細胞滲透壓, 影響藻細胞吸收營養鹽的能力, 因此也影響微藻的生長。產生這種現象的原因可能是Na+在滲透調節和細胞膜電位調節中起重要作用, 然而, 過量的NaCl導致與光系統Ⅱ結合的多肽解離, 從而降低了光合機制的電子傳遞[41]。
3.2 鹽度和pH對平凡舟形藻葉綠素a含量的影響
一般認為, 不同藻類的最適pH不同, 極端的pH條件影響藻類的光合作用、生長和養分同化[42]。在本研究中, 在酸性條件下(pH 4.5和6.0)和堿性條件下(pH 9.5和 10.5), 這4個pH使平凡舟形藻的細胞密度、葉綠素a含量顯著減少, 對葉綠素熒光參數Fv/Fm和Fv/Fo也產生了抑制, 反映了pH過高或者過低對反應中心PSⅡ具有破壞作用, 使之受到限制[43]。在pH為7.5時, 細胞密度和葉綠素a含量達到最大值,葉綠素熒光參數Fv/Fm和Fv/Fo高, 其次是pH為8.5。王銘等[44]報道, 在pH為7.5時微藻的生理狀態與光合作用表現為最佳。由此可見, 過酸或者過堿都不利于平凡舟形藻的生長, pH為7.5是最適宜平凡舟形藻生存的環境。Sakarika等[45]的研究表明在堿性值7.5和8.0時, 微藻的生長超過對照組20%—25%,表明在這個狹窄的pH范圍內生長條件是最優的。微藻培養過程中過酸或者過堿會對平凡舟形藻的細胞產生一定的傷害, 使其破壞或死亡, 影響其生長。
3.3 鹽度和pH對平凡舟形藻油脂含量的影響
鹽度脅迫影響著與微藻生長發育相關的各種生理生化機制, 它也可以引起微藻脂肪含量的增加,它在引起脂肪酸代謝變化方面也發揮了重要作用[46]。在本研究中, 在0—0.24 mol/L隨著鹽濃度的逐漸升高, 總脂含量漸漸增加由13.7%上升到34.9%。對于脂肪含量, Hu[47]發現鹽度的上升伴隨著滲透壓的增加, 藻類的總脂含量略有增加。在不同鹽濃度脅迫下, 微藻的生長條件會發生改變, 從而對總脂含量產生影響。本研究表明, 高鹽度條件(鹽度高于0.12 mol/L)會抑制藻類的生長, 但高鹽度有利于藻類總脂的積累。當氯化鈉濃度從0.06增加到0.4 mol/L時, 普通小球藻的含油量可以提高到49%[48], 比本實驗中的總脂含量高。有研究表明梅尼小環藻在鹽度為25 g/L脂質含量達到最高, 達21%[49], 比本實驗研究中的低。所以, 最適鹽度和葉綠素含量趨勢的差異可能因藻種而異。因此, 應選擇合適的鹽度來提高脂肪產量。這說明鹽濃度的增加可以促進平凡舟形藻的油脂積累。有可能是因為在高濃度鹽調控下微藻的生長需要更多能量, 使藻細胞內儲存的油脂含量相應增加[50]。也有研究表明, 細胞脂質的增加可能是對滲透脅迫引起的微藻細胞損傷而形成的一種防御機制[51]。鹽脅迫有效地促進了微藻細胞脂質的積累。到目前為止, 已有研究證明鹽脅迫可以有效促進微藻細胞中脂質的積累[3,52,53], 但與此同時, 通常也會顯著抑制微藻的生長, 導致最終微藻的脂質生產力不能得到很大的提高[54,55]。為了克服生物量產量和脂質生產之間的矛盾, 需要一種允許最佳生長速度和允許脂質增強的培養模式。有報道稱在微藻大規模培養中, 首先提供最適合微藻生長的最佳條件, 隨后需要具有應激條件的介質來誘導脂質生物合成[56], 以確保微藻的生長和收獲更多油脂。
pH的不同也引起了平凡舟形藻總脂含量的變化。pH為7.5時總脂含量最高, 其余pH處理水平下的總脂含量低。對于pH脅迫, 據報道在最適pH條件下培養微藻可以提高生物量和脂肪含量, 在pH為7.5時, 可以顯著提高微擬綠球藻的生物量和總脂含量[57]。但也有研究表示微藻在堿性的環境下利于油脂積累[58], 與本研究結果不同。所以不同的藻種對pH適應性不同, 應選用合適的pH。在pH為7.5時是最有利于平凡舟形藻的生長, 也可以提高其總脂含量。Sakarika等[59]報道稱微藻細胞在適應了新的pH條件后, 代謝被激活, 除了繁殖外, 微藻還消耗提供的碳源(葡萄糖), 以積累脂質。
淡水藻株和海水藻株在不同環境因素下誘導油脂含量不同, 本實驗中所使用的藻種為淡水藻株,不同淡水藻株在鹽度和pH脅迫下的生長狀況和總脂含量也不相同。根據藻類及其自然棲息地的不同, 誘導脂質積累的最佳氯化鈉濃度水平也不同。淡水藻株Desmodesmus abundans在添加氯化鈉的情況下, 可獲得較高的油脂產率[56]。Chlorella sorokinianaHS1是一株淡水小球藻, 但其在海水鹽度下也能生長, 并積累脂質, 在淡水中培養, 隨后在咸水中積累脂肪。這可以緩解大規模養殖中的應激誘導等生產限制, 特別是營養限制, 以實現更高的脂肪生產率[60]。豐富柵藻在pH為6和8時是最適合生長和油脂積累[58]。淡水微藻在環境脅迫下特別是在海水鹽度或者高于海水鹽度的條件下, 會導致微藻油脂的積累。使微藻在淡水或廢水中培養, 導致微藻生物量的增加, 然后在鹽水中培養以積累脂質。這可以緩解生產限制, 以實現更高的脂質生產力。此外, 也有一些關于鹽度和pH對各種海洋微藻的生長和脂肪積累影響的報道。梁英等[61]研究表明鹽度對6種海洋硅藻生長與脂肪酸均有不同影響,證明鹽濃度對于不同藻種作用不同。有報道研究了在同一鹽度下5株海洋硅藻的總脂含量, 從而篩選出3種具有開發潛能的海洋硅藻[62]。江靈芝等[63]以蛋白核小球藻820為實驗材料, 研究了3種鹽度對其生長和油脂含量的影響, 以了解該小球藻對鹽度的適應能力。研究結果表明, 蛋白核小球藻820的生長隨鹽度增加而變慢, 而油脂含量隨鹽度增加而升高。在300 mmol/L氯化鈉培養處理的原殼小球藻雖然生物量只有對照的73.55%, 但是總脂含量卻是對照的2.22倍, 可見一定濃度的氯化鈉處理可顯著提高原殼小球藻的油脂含量[64]。當培養液鹽度上升至2.0%時, 是不添加NaCl培養基中微藻的油脂含量的1.6倍, 表明通過一定的鹽度脅迫將微藻的油脂含量提升了60%左右。鹽度對于提高微藻的油脂含量具有明顯的促進作用[65]。Mus等[66]在研究pH對三角褐指藻的實驗中發現在pH從8.0變化為7.5時, 脂肪含量增加。目前, 利用海洋微藻生產油脂的研究較多, 對淡水微藻相關研究相對較少, 本研究可以為淡水硅藻產油方面提供基礎資料。
隨著水體鹽堿化程度的日益嚴重和淡水資源的日益匱乏, 藻類被視為發展生產可持續能源的最佳原材料, 鹽脅迫是最常見的一種環境刺激, 在微藻細胞中, 鹽脅迫可以影響脂類的合成, 微藻在一定程度上都可以耐受一定的鹽濃度, 但是過量的鹽脅迫也會影響光合作用進而影響生物量和脂質合成量。氯化鈉是非常廉價的鹽類, 同時也是營養的組成部分, 通過調整培養基中氯化鈉的濃度達到不影響生物量同時增加脂含量, 可以大大降低生產成本。添加氯化鈉和找最適pH是一種經濟適用的增脂策略, 可考慮應用于微藻生物柴油的生產。
4 結論
微藻的生長受到光照強度、營養元素、溫度和pH等培養條件的影響, 進而影響微藻的油脂產量, 為探究平凡舟形藻在不同鹽度和pH培養條件下生長和油脂含量的影響, 進行為期12d的脅迫培養。結果表明, 鹽濃度較低時可以促進平凡舟形藻的生產能力, 隨鹽濃度升高, 平凡舟形藻細胞生長受到抑制, 鹽濃度為0.12 mol/L是適合平凡舟形藻生長的鹽濃度。但總脂含量則相反, 隨著鹽濃度的不斷增加至0.24 mol/L, 總脂含量達到34.9%。在pH 7.5的培養條件下, 平凡舟形藻表現出較高的細胞密度增長, 油脂含量升高。藻類可以在應急條件下產生大量的脂質, 這些脂質可以很容易地轉化為可替代的燃料, 是一種可持續生物燃料來源。本研究結果表明, 適當改變培養基中鹽的濃度和pH, 可以調控平凡舟形藻的生長速度和油脂含量, 本研究為微藻生產生物柴油提供參考數據。微藻作為可持續的生物能源原料, 具有巨大的發展潛力。在營養物質耗盡的情況下, 淀粉合成會轉向油脂的合成, 因此脅迫條件將是提高微藻油脂含量的重要手段之一。如何將培養條件與脅迫方式有機結合, 從而使微藻細胞既能達到生物量最大, 也能保證油脂的積累量最多, 還需進一步進行深入研究。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
