水稻明恢826光身基因GL826的精細定位與候選基因分析
2024-02-23 06:51:40韋新宇張文靜曾躍輝肖長春黃建鴻許旭明
核農學報 2024年3期
韋新宇 張文靜 曾躍輝 肖長春 張 銳 黃建鴻 許旭明,
(1三明市農業科學研究院生物技術研究所,福建 三明 365500;2福建省(山區)作物遺傳改良與創新利用重點實驗室,福建 三明 365500;3三明市農業科學研究院水稻研究所,福建 三明 365500)
光身稻(Oryzaglaberrima)因其莖稈、葉片和谷殼等組織表面光滑無毛而得名,又叫光葉稻或光殼稻。光身稻具有廣親和性、對強光適應能力強、抗倒伏性強、適合機械化操作和節省儲運空間等優良特性[1-4],已作為雜交水稻育種重要的種質資源被加以研究和利用[5]。
植物表皮毛起源于表皮細胞,是一種表皮組織向外延伸的毛狀突起,在植物體中普遍存在,可由單細胞或多細胞構成,形態結構多樣[6-8]。水稻的表皮毛為雙細胞毛狀體,呈圓柱狀分布[9]。目前,對水稻表皮毛性狀的研究主要集中在葉片和穎殼[5]。光身稻的無表皮毛性狀主要表現為葉片和穎殼表面毛狀體的減少或缺失。已報道的與水稻茸毛生長發育相關的基因HL6[10]、HL1[11]和GL6[12],均已被定位在水稻第6 號染色體上,且葉片長毛表型均受細胞核單基因控制,屬顯性遺傳。已報道的水稻光身基因wmg2[13]位于第3 號染色體上,wmg1[13]位于第4 號染色體上,GL5[10]、NUDA/GL-1[14]、dep[9]、gl-l[15]、GLAG5[16]位于第5 號染色體上,GLR3[17]、OsSPL10[18]、gl6[19]、GLL[20]位于第6 號染色體上。其中,GL5基因被精細定位在水稻第5號染色體33 kb 區間內,GL5基因的表達水平降低導致表皮毛的缺失[10];NUDA/GL-1基因座被精細定位在水稻第5 號染色體28.5 kb 區間內,通過RNAi 和互補分析,確認一個編碼具有WUSCHEL結構域蛋白的同源基因(OsWOX3B)作為候選基因[14];dep基因被精細定位在水稻第5 號染色體22 kb 區間內,互補試驗證實dep基因對無毛表型具有調節作用[9]。位于第6號染色體上的OsSPL10啟動子區發生變異導致秈稻品種WD-17993出現無毛性狀[18],也有研究表明OsSPL10是水稻茸毛發育的正向調控基因,其與OsWOX3B、OsPLT2基因間存在遺傳互作,共同調控水稻表皮毛的起始發生和伸長[21]。
明恢826(MH826)是三明市農業科學研究院利用親本IR249和NO3S2雜交配組,經多世代自交選擇、抗病鑒定以及與大量不育系測配篩選育成的優良光身稻恢復系。該恢復系具有株型好、結實率高、米質優、配合力強等優點,所配雜交稻組合已陸續進入生產應用,其育種利用和研究價值較好。但目前對MH826 的光身性狀及控制光身性狀的基因尚不了解。因此,本研究對MH826的光身性狀進行顯微觀察和遺傳學分析,利用分離群體分析法(bulked segregation analysis,BSA)和隱性群體分析法(recessive class analysis,RCA),對其光身基因進行精細定位和候選基因分析,以期為進一步明確其基因功能和分子機制奠定基礎,同時為光身稻品種及其光身基因在水稻育種上的應用提供理論依據。
1 材料與方法
1.1 試驗材料
利用光身稻恢復系MH826 與具有明顯表皮毛特性的水稻品種華占、明恢2155和9311雜交,獲得F1代。F1代自交后獲得F2代分離群體,用于光身基因的遺傳學分析和精細定位。2020 年晚季在福建沙縣三明市農業科學研究院水稻試驗基地種植親本、F1和F2代群體,常規田間管理。
1.2 表型鑒定與遺傳分析
分別在苗期、分蘗期和抽穗期對親本、F1和F2代分離群體單株進行表型鑒定和統計學分析。以手指自葉尖沿葉面和邊緣向葉基部觸摸無粗糙感作為光身表型鑒定依據。通過統計F2代分離群體中光身單株和有毛單株的分離比進行遺傳學分析,并利用χ2進行適合性檢驗。
1.3 掃描電鏡觀察
選取水稻分蘗期的葉片和灌漿期的種子用2.5%戊二醛固定液于4 ℃冰箱內固定1 h 以上,然后用0.2 mol·L-1的磷酸緩沖液(pH 值7.2)漂洗3 次,每次20 min,再用1%鋨酸固定1 h,然后用0.2 mol·L-1的磷酸緩沖液(pH 值7.2)漂洗3 次,每次20 min,再用梯度乙醇溶液進行脫水,每次20 min。之后在K850 CO2臨界點干燥儀(Quorum,英國)內進行干燥處理,再將樣品粘在樣品臺上用MC1000 離子濺射儀(HITACHI,日本)進行離子濺射鍍膜處理,最后利用SU8100 掃描電子顯微鏡(HITACHI,日本)觀察并拍照[22]。
1.4 基因的精細定位
采用改良的十六烷基三甲基溴化銨(cetyltrimethyl ammonium bromide,CTAB)法[23]提取水稻苗期葉片全基因組DNA。在F2分離群體中分別隨機選擇15 個光身單株和15個有毛單株提取基因組DNA,濃度統一稀釋為50 ng·μL-1,等量混合從而構建光身基因池和有毛基因池[24]。通過水稻生物信息學網站(http://www.gramene.org/)搜索PCR 擴增產物大小在300 bp 以內,且均勻分布于水稻12條染色體上的356 對簡單重復序列(simple sequence repeat,SSR)引物對親本和基因池進行多態性檢測并對F2隱性單株進行基因型分析,篩選與目標基因緊密連鎖的分子標記。PCR擴增反應體系為10 μL:2×TaqPCR MasterMix(艾德萊,北京)5.0 μL,50 ng·μL-1DNA 模板0.8 μL,10 μmol·L-1正、反向引物各0.5 μL,ddH2O 3.2 μL。PCR 擴增反應程序:94 ℃預變性3 min;94 ℃變性10 s,55~60 ℃退火10 s,72 ℃延伸10 s,35 個循環;72 ℃終延伸5 min;4 ℃保存。PCR 擴增反應結束后,將PCR 擴增產物用8.0%的非變性聚丙烯酰胺凝膠進行電泳,通過GelDoc XR+凝膠成像系統(Bio-Rad,美國)進行拍照觀察。另外,根據粳稻品種日本晴和秈稻品種9311 之間的基因組序列差異,利用Primer Premier 5.0 軟件進一步設計和開發分布密度更高的插入缺失(insertion-deletion,InDel)標記,用于光身基因GL826的精細定位。PCR 反應及電泳程序方法和步驟同上。利用Mapmaker/Exp3.0 軟件進行連鎖分析,采用Kosambi 運算函數將重組率轉換為遺傳距離,結合MapDraw v2.1 軟件繪制遺傳連鎖圖譜。根據水稻生物信息學網站(http://www.gramene.org/)提供的水稻基因組序列信息并通過染色體步移的方法構建物理圖譜。
1.5 候選基因分析
通過水稻基因組注釋數據庫(Rice Genome Annotation Project,RGAP,http://rice.plantbiology.msu.edu/)對精細定位區間內候選基因的功能進行預測,并結合實時熒光定量PCR(quantitative real-time PCR,qRT-PCR)以及克隆和測序技術對候選基因進行表達和序列分析,具體方法參照文獻[25]。
2 結果與分析
2.1 MH826表型鑒定和掃描電鏡分析
本研究以手指觸摸的方式對研究材料的表型進行初步鑒定。結果表明,具有表皮毛特性的水稻品種華占、明恢2155和9311的葉片和籽粒穎殼具有明顯的粗糙感,手指在葉片和穎殼表面滑動時存在明顯的阻滯感;而光身稻恢復系MH826 的葉片和籽粒穎殼觸摸光滑,無粗糙感。進一步采用掃描電鏡分析了具有表皮毛特性的水稻品種華占和光身稻恢復系MH826 分別在分蘗期葉片以及灌漿期籽粒的表皮毛發育情況。結果表明,華占和MH826 在葉片和籽粒表皮存在顯著的表皮毛差異。在華占的葉片和籽粒表皮可觀察到明顯的尖刺狀表皮毛,包括大刺毛和小刺毛,且在葉片表面分布相對密集(圖1-A~D)。而在MH826 的葉片和籽粒表皮僅存在少量的腺毛,未觀察到明顯的尖刺狀表皮毛,呈現表面光滑特性(圖1-E~H),其與手指觸感鑒定結果表現一致。
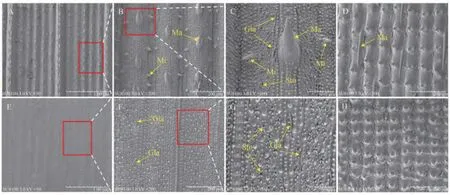
圖1 華占和MH826的葉片、籽粒掃描電鏡觀察Fig.1 Observation of leaf and grain phenotypes of two parental lines (Huazhan and MH826) by scanning electron microscope
2.2 MH826光身性狀的遺傳學分析
為了對MH826 的光身性狀進行遺傳特性分析,本研究利用MH826 分別與具有表皮毛特性的水稻品種華占、明恢2155 和9311 配制雜交組合,獲得F1代并構建了3 個獨立的F2代分離群體。在分蘗盛期,分別對華占×MH826,明恢2155×MH826 和MH826×9311 獲得的F1代植株以及F2代群體進行葉片表型鑒定和統計學分析。結果表明,雜交獲得的所有F1代植株葉片均表現為明顯的表皮毛性狀,與具有表皮毛特性的親本葉片表型表現一致,說明MH826 的光身性狀屬于隱性遺傳。另外,在3 個獨立的雜交F2代群體中,顯性單株(有毛單株)和隱性單株(光身單株)表現出明顯的分離,且均符合孟德爾單基因控制的3∶1分離比(χ2<χ20.05)(表1)。以上結果表明,MH826的光身性狀由單個細胞核隱性基因控制。本研究將其命名為GL826( Glabrous Leaf 826)。
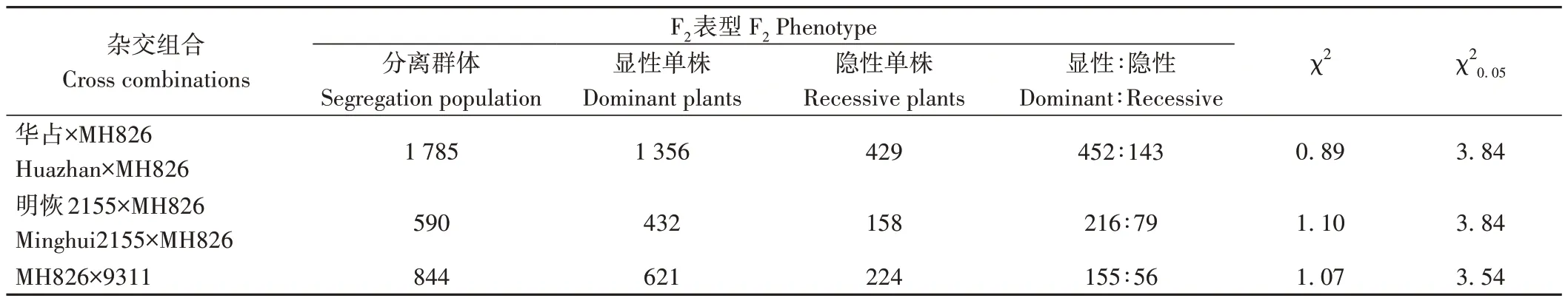
表1 MH826雜交組合的分離分析Table 1 Segregation analysis of MH826 crosses
2.3 GL826基因的精細定位
由表1 可知,華占×MH826、明恢2155×MH826 和MH826×9311 構建的3 個F2代分離群體植株數量分別是1 785、590 和844 個。綜合考慮基因精細定位對大群體數量的要求,本研究優先選用群體植株數量較大的華占×MH826 獲得的429 個F2隱性單株對光身基因GL826進行精細定位。首先利用平均分布于水稻12條染色體上的356 對SSR 引物對親本以及兩個極端基因池進行多態性篩選,通過對F2隱性單株進行基因型鑒定,并結合Mapmaker/Exp 3.0 進行連鎖分析。獲得一個位于水稻第5 號染色體上的SSR 標記Rm13 與GL826基因連鎖。進一步在Rm13 附近篩選分布密度更高的SSR 標記對兩親本、基因池以及F2隱性單株進行多態性檢測和基因型分析,獲得與GL826連鎖的SSR標記共7 對:Rm17731、Rm6300、Rm17787、Rm17812、Rm17819、Rm5579 和Rm405。并將GL826初步定位在Rm17787 和Rm17812 之間,相對遺傳距離分別為1.6 和2.4 cM(圖2-A)。通過比對粳稻品種日本晴和秈稻品種9311 基因組序列,在Rm17787 和Rm17812標記之間開發獲得插入缺失InDel 標記95 對,進一步對親本、基因池以及F2隱性單株進行多態性檢測和基因型分析,獲得與GL826緊密連鎖的特異性InDel標記19 對:InDel-02、InDel-11、InDel-13、InDel-14、InDel-17、InDel-24、InDel-26、InDel-28、InDel-31、InDel-40、InDel-44、InDel-48、InDel-61、InDel-64、InDel-68、InDel-70、InDel-71、InDel-72 和InDel-76。最終將光身基因GL826精細定位在分子標記InDel-70 和InDel-11 之間,且相對遺傳距離均為0.2 cM(圖2-A)。結合RGAP 數據庫,該精細定位區間對應粳稻品種日本晴的物理距離為141.03 kb(圖2-B)。參考日本晴各細菌人工染色體(bacterial artificial chromosome,BAC)克隆序列,并通過染色體步移的方法將每個與光身基因GL826連鎖的分子標記在染色體上的位置錨定在相應的BAC 克隆上,從而構建了覆蓋GL826基因位點的物理圖譜(圖2-B)。本研究用于GL826精細定位的InDel標記序列及擴增片段大小如表2所示。
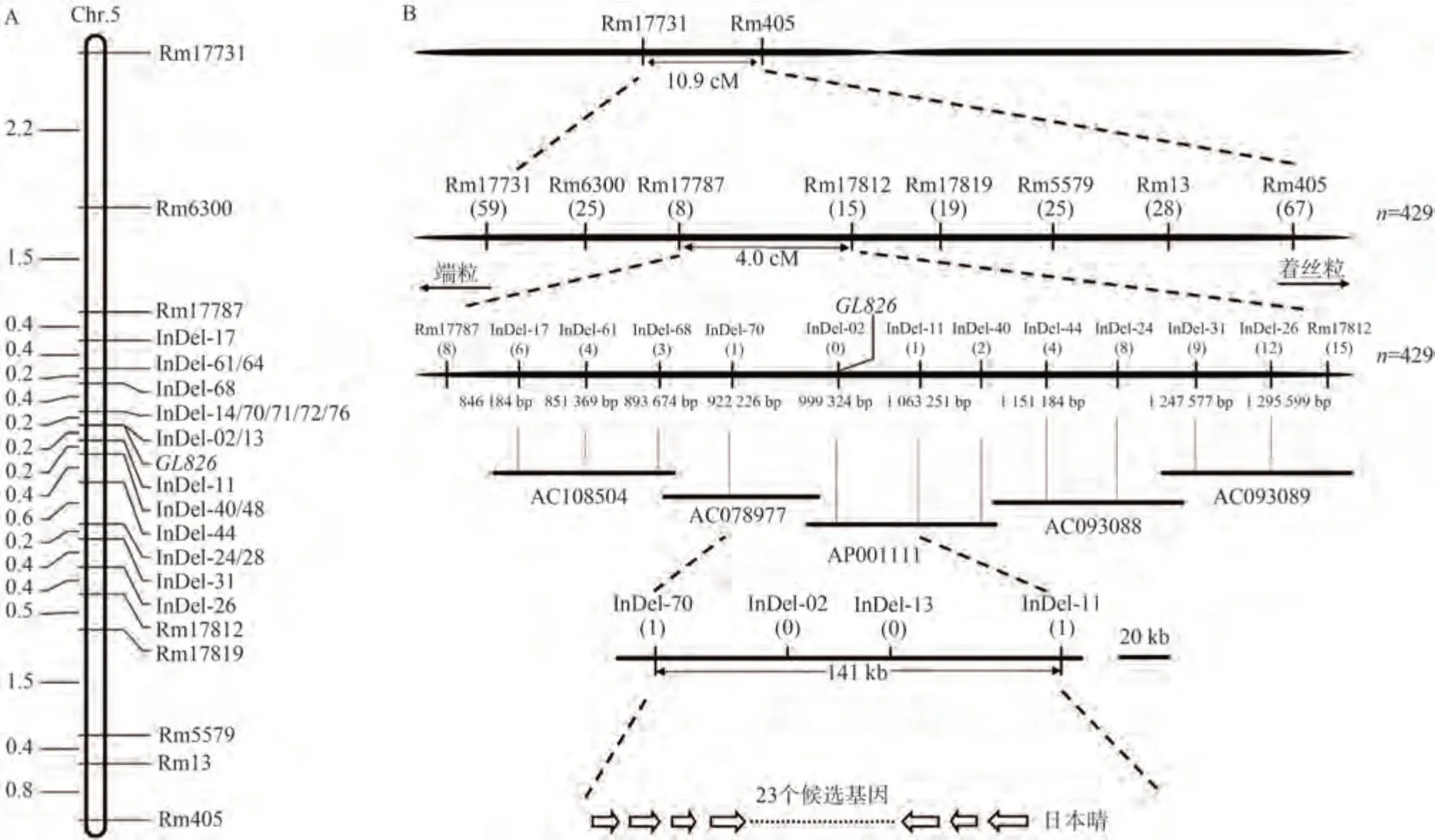
圖2 光身基因GL826精細定位Fig.2 Fine mapping of glabrous leaf gene GL826
2.4 GL826候選基因分析
參考RGAP 水稻基因組注釋數據庫,發現在該精細定位區間共預測有23 個開放閱讀框(ORFs)(表3)。其中候選基因LOC_Os05g02730為已報道的與水稻表皮毛發育相關的基因OsWOX3B,該基因編碼一個WOX3B轉錄因子家族蛋白,參與植物特異性橫向器官的發育調節,OsWOX3B基因突變將導致水稻葉片和籽粒穎殼表皮毛缺失,表現為植株整體光滑無毛的表型[9,14]。因此,本研究將LOC_Os05g02730作為GL826的重點候選基因并進行進一步的序列和表達分析。
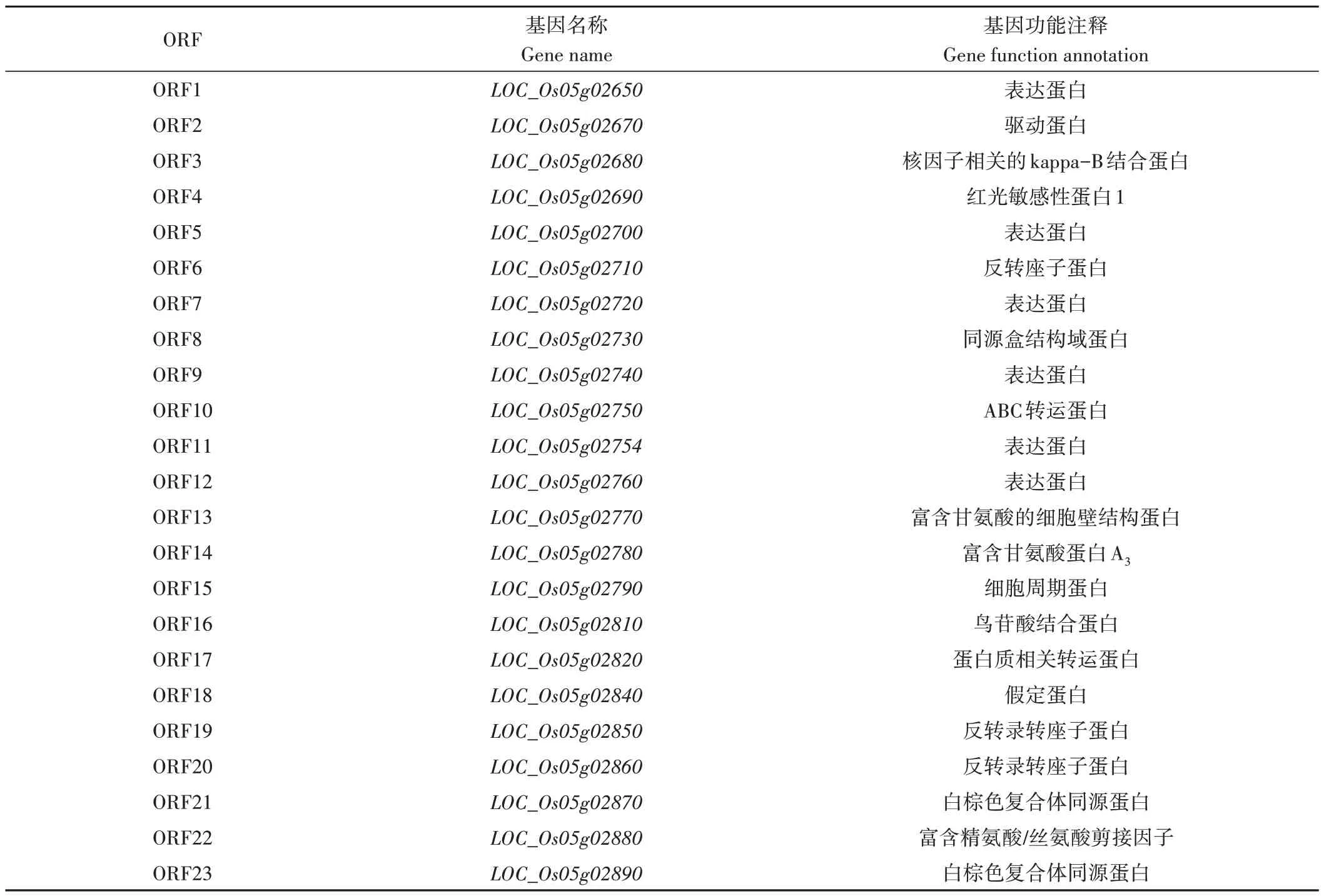
表3 23個候選基因功能注釋Table 3 Functional annotations of twenty-three candidate genes of GL826 based on the RGAP database
為了鑒定候選基因LOC_Os05g02730是否為GL826,本研究通過PCR 擴增和測序技術首先克隆和分析了光身稻恢復系MH826 和具有表皮毛特性的水稻品種華占、明恢2155 和9311 中LOC_Os05g02730基因的編碼序列。結果表明,華占、明恢2155、9311和MH826在LOC_Os05g02730基因的編碼區未檢測到堿基差異,且序列和已報道的OsWOX3B[9,14]基因序列完全一致(圖3-A)。即光身稻恢復系MH826 中含有LOC_Os05g02730(OsWOX3B)基因完整的編碼序列(coding sequence,CDS)。另外,進一步選擇LOC_Os05g02730基因起始密碼子上游1.8 kb 啟動子區域進行PCR 擴增和序列比對分析。結果表明,華占、明恢2155、9311和MH826 在該啟動子區域同樣未檢測到堿基差異并且和OsWOX3B基因啟動子序列表現一致(圖3-B)。因此,LOC_Os05g02730基因在具有表皮毛特性的水稻品種華占、明恢2155、9311 和光身稻恢復系MH826 中未鑒定到DNA 水平差異,包括編碼區和基因上游的啟動子區域。本研究用于LOC_Os05g02730基因CDS 和啟動子序列測序的特異性引物如表4所示。

表4 用于LOC_Os05g02730基因測序和表達分析的引物Table 4 Primers used for sequencing and expression analysis of LOC_Os05g02730 in this study
為了進一步確定候選基因LOC_Os05g02730即為光身基因GL826,本研究利用qRT-PCR 技術分析了LOC_Os05g02730基因在光身稻恢復系MH826 和具有表皮毛特性的水稻品種華占在不同時期以及不同組織部位中的表達差異,其中分別包括LOC_Os05g02730在苗期、分蘗期和抽穗期的根、莖、葉以及抽穗期幼穗組織中的表達。結果表明,LOC_Os05g02730基因在MH826 苗期、分蘗期和抽穗期的莖、葉片以及抽穗期的幼穗中的表達水平均極顯著低于具有表皮毛特性的水稻品種華占,尤其在葉片中的表達下調最為明顯(圖4)。說明LOC_Os05g02730基因在光身稻恢復系MH826 中的表達受到了嚴重的抑制,其表達量在不同生長發育階段不同組織中表現為不同程度的下調。因此,推測LOC_Os05g02730即為GL826基因。本研究用于LOC_Os05g02730基因表達分析的特異性引物如表4所示。
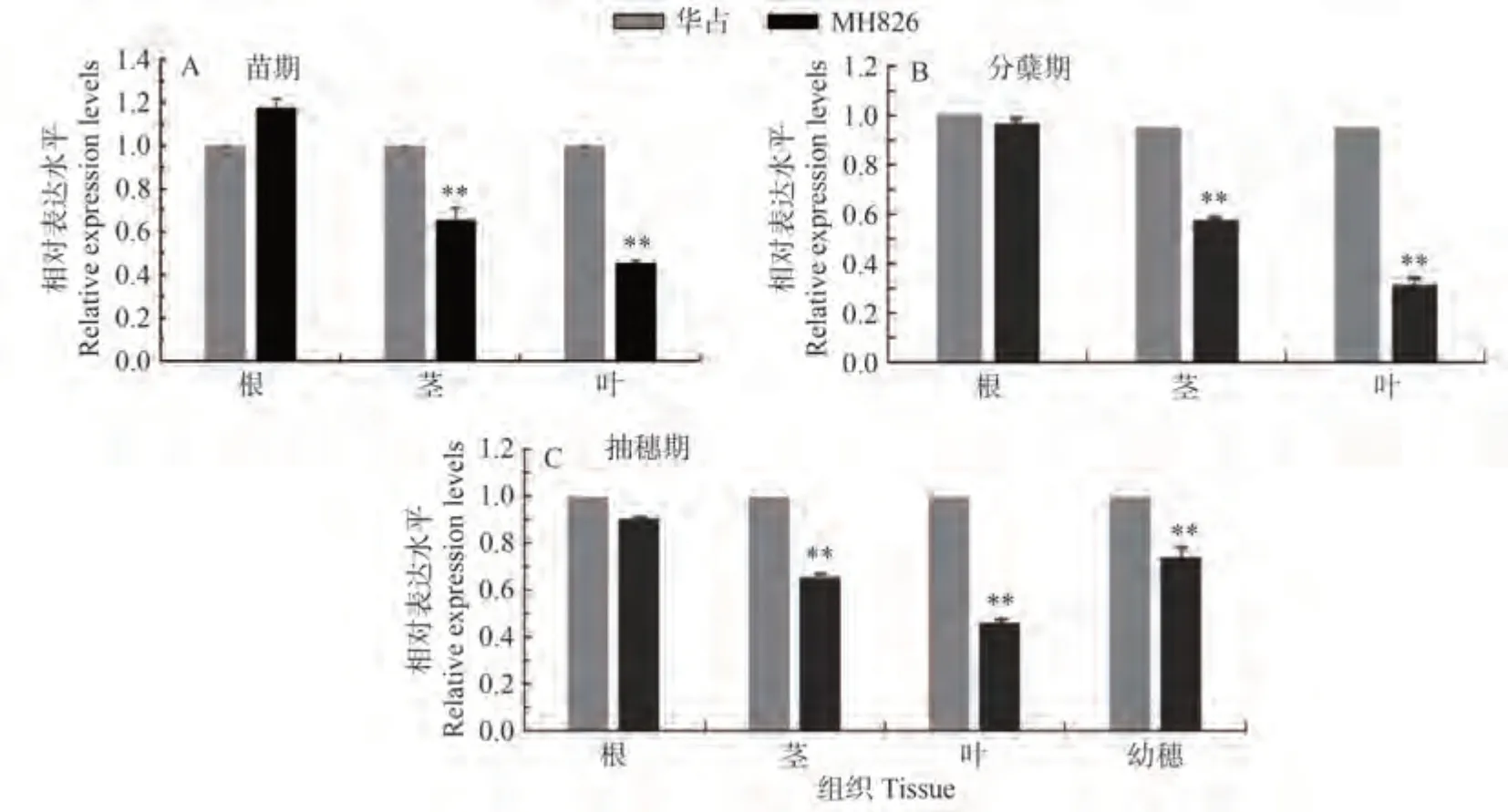
圖4 LOC_Os05g02730在華占和MH826不同時期不同組織中的表達模式分析Fig.4 Expression patterns of LOC_Os05g02730 in various tissues of huazhan and MH826 during different growth stages,determined by qRT-PCR analysis
3 討論
MH826 是三明市農業科學研究院經多世代自交選擇、抗病鑒定以及大量不育系測配篩選育成的優良光身稻恢復系。本研究通過手指觸感和掃描電鏡觀察進行光身表型鑒定,結果顯示MH826 的葉片和穎殼表面手指觸摸光滑,無粗糙感。掃描電鏡同樣未觀察到明顯的尖刺狀表皮毛,呈現表面光滑表型,二者鑒定結果一致。在明確了MH826 光身表型的基礎上,利用MH826 與具有表皮毛特性的水稻品種華占、明恢2155和9311 雜交,構建多個F2代分離群體對其光身性狀進行遺傳學分析。結果表明MH826 的光身性狀由細胞核隱性單基因控制。目前已報道的調控水稻光身性狀的基因NUDA/GL-1[14]、OsWOX3B[26]、dep[9]、GLR1[13]、GL5[27]、gl1[28]、OsSPL10[29]、GLR3[17]、glr2[30]、GLL[20]等也均受細胞核隱性單基因控制。
本研究利用MH826與華占雜交構建的429個F2代隱性(光身性狀)單株對GL826基因進行精細定位。將GL826基因定位在水稻第5號染色體短臂端,位于分子標記InDel-70 和InDel-11 之間,相對遺傳距離均為0.2 cM,區間物理距離為141.03 kb。參考RGAP 數據庫,發現在該精細定位區間共注釋有23 個完整的ORF。其中LOC_Os05g02730基因[9,14]為前人已報道的與水稻表皮毛性狀發育相關的基因。因此,本研究重點將LOC_Os05g02730作為GL826的候選基因,并作進一步的序列比對和表達模式分析。結果發現LOC_Os05g02730基因在光身稻恢復系MH826 苗期、分蘗期葉片、莖以及抽穗期葉片、莖和幼穗組織中的表達表現出不同程度的下調,說明LOC_Os05g02730基因在光身稻恢復系MH826 中的表達受到了嚴重抑制。Zhang等[14]認為LOC_Os05g02730基因與WUSCHEL-like基因(OsWOX3B)同源。OsWOX3B屬于WOX3家族中的一個亞族,其主要調控水稻葉片形態的發生以及表皮毛的生長和發育,同時影響水稻側生器官的發育和形成。序列分析結果表明,LOC_Os05g02730與水稻的OsWOX3以及擬南芥的PRS、玉米的NS1和NS2具有高度相似性[9,30-31],其功能對于表皮細胞的分化至關重要。因此,綜合以上表型鑒定、構建3個獨立群體的遺傳分析、精細定位、LOC_Os05g02730基因的表達模式分析及其功能注釋等分析結果,本研究認為候選基因LOC_Os05g02730(OsWOX3B)即為GL826基因。然而本研究在進行基因序列比對時發現,LOC_Os05g02730基因在光身稻恢復系MH826 和具有表皮毛特性的水稻品種華占、明恢2155 和9311 之間均未檢測到DNA水平上的差異,包括CDS 編碼區和基因上游的啟動子區域(圖3)。這與NUDA/GL-1[14]、OsWOX3B[26]、dep[9]報道的研究結果一致,即LOC_Os05g02730基因在有毛水稻和光身水稻品種間不存在DNA 水平上的差異,因而推測其光身表型可能由表觀遺傳修飾引起。其中關于DNA 甲基化的研究較多,如水稻甲基化轉移酶SDG714的功能缺失,會導致基因組甲基化水平明顯降低,進而導致植株表現出光葉和光殼表型[32]。無毛非洲栽培稻的穎殼無毛性狀是由于hAT轉座子插入到控制毛狀體發育基因GLAG5(GLABROUS GLUME 5)上,引發DNA 甲基化,進而抑制該基因的表達而產生無毛性狀[16]。另外,組蛋白乙酰化也被發現參與調控植物表皮毛的發育,如GCN5(GENERAL CONTROL NON DEREPRESS-IBLE 5)通過組蛋白乙酰化作用于GL1、GL2和GL3的啟動子,參與抑制擬南芥的表皮毛發育起始[33]。miRNA 作為一類進化上保守的小分子RNA,已有研究表明其參與了植物表皮毛的發育,如在擬南芥中,SPL(SQUAMOSA PROMOTER BINDING PROTEIN LIKE)是調控表皮毛發育的一類轉錄因子,miRNA156通過調控SPLs的表達來調控表皮毛發育,miRNA156的參與會導致SPLs的表達量增加,擬南芥主莖上表皮毛的數量相應減少[34]。另有研究報道,水稻光身突變體的光葉或光殼表型的形成可能由RNA 的折疊等二級結構變化導致[5,28]。由此可知,關于光身稻恢復系MH826 光身基因GL826的具體調控機理有待進一步研究。
前人對水稻光身基因的研究多采用光身突變體或遠緣種質作為研究材料,而本研究則采用新育成的已在生產上應用的光身稻恢復系作為研究材料,這使得定位和克隆的基因更能與生產實際結合,能夠直接在育種和生產實踐中加以利用。近年來,光身稻品種的選育和種質資源的應用越來越多。張世輝等[35]利用光殼遺傳背景材料AF6-82/R80 與三合占/農香16 雜交,經多代自交、回交,育成香型優質稻品種農香32。陳錦文等[36]以光身稻澳大利亞-112 與明恢77 雜交,經多世代選育,育成光身稻恢復系泉恢404,并配組育成雜交稻組合福農優404。本課題組利用光身稻恢復系MH826 配制的雜交稻新組合瀘優826、廣優826、明優826 均表現產量高、米質好、千粒重大、抗倒伏性強等優點,應用前景較好。
4 結論
光身稻恢復系MH826 的光身表型由單個細胞核隱性基因控制。利用MH826 與華占雜交構建的F2分離群體將GL826 基因精細定位在水稻第5 號染色體短臂端,位于InDel-70 和InDel-11 之間約141.03 kb 區域內。通過序列分析和表達模式分析,確定候選基因LOC_Os05g02730即為GL826基因。
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
