特色香料作物油樟葉內生真菌多樣性及其揮發性成分分析
2024-02-23 06:51:46劉雯雯胡連清杰周萬海馮瑞章
核農學報 2024年3期
劉雯雯 胡連清 陳 露 楊 麗 魏 琴 周 杰周萬海 , 馮瑞章
(1宜賓學院,四川省油樟工程技術研究中心,四川 宜賓 644007;2宜賓學院,農林與食品工程學部,四川 宜賓 644007;3四川宸煜林業開發有限責任公司,四川 宜賓 644007)
油樟(Cinnamomumlongepaniculatum)是我國特有的木本香料林業資源,其中提取的天然芳香油在醫藥、日化和食品等行業發揮著重要作用[1]。油樟葉部含油量最高,主要成分為1,8-桉葉油素[2]、α-松油醇、β-蒎烯和γ-松油烯等活性成分[3-4],是醫藥、日化和食品等生產的重要原料[5],且有研究表明油樟精油具有抗氧化、抑菌和抗腫瘤等作用[6-7]。關于油樟精油的提取和成分分析已有相關報道,例如尹浩等[8]研究表明采用酶輔助同蒸餾萃取法提取的油樟葉精油收率最高;寧登文等[9]發現4、8 和12 月采集的油樟葉出油率最高,樹齡21~40 年且嫩葉和混合葉的相對出油率最高;程賢等[10]研究結果顯示春季油樟葉精油提取率高于秋季,但兩季精油主要成分無顯著差異。四川宜賓油樟葉含油量約為江西、廣東及臺灣等省油樟油量的2.5倍,且比香樟葉含油率高2 倍[11],推測與其共生微生物等特殊環境條件有關[12]。
植物內生真菌指長期或生活史某一個階段定殖于健康植物內部而不引起宿主明顯病理癥狀的微生物[13],研究證實其種類和數量隨宿主植株的種類、季節或器官等不同而不同[14]。近年來,蛇足石杉[15]、粉色西番蓮[16]、樟葉越橘[17]等不同生境植物內生真菌得到了廣泛研究。部分內生菌可產生與宿主相同或相似的活性物質,在農業、醫藥和生物防治等方面具有較大潛力[18]。關于油樟內生菌的多樣性和功能性研究已有一些報道,如馮瑞章等[19]從油樟中篩選到3 株具有溶磷作用的內生細菌,對植物生長、抗性和生物防治具有應用效果;黃金鳳[20]通過紫外線誘變篩選出油樟葉中高產α-松油醇的內生細菌;嚴寬等[21-22]發現用油樟內生真菌2J1 和3J1 的孢子懸浮液處理油樟葉21 d 能顯著提高葉片的揮發油含量,建立的青霉菌屬油樟內生真菌和茉莉酸甲酯培養體系,促使了油樟細胞揮發油的合成。譚韻雅等[23]發現油樟內生真菌YY26 和YG42可促進1,8-桉葉油素等成分的積累,抑制了YG71代謝產物的分泌。但關于油樟葉可培養內生真菌的多樣性、菌株與產油量關聯性及菌株揮發性成分分析研究還較少。基于此,本研究以四川宜賓油樟為材料,測定不同株系和不同季節油樟葉的芳香油量,研究其葉部內生真菌的多樣性和分布情況,探討不同株系、季節的菌株種類與含油量關系,并測定菌株發酵液的揮發性成分,以期為油樟內生真菌資源多樣性及提供可應用的功能菌株奠定基礎。
1 材料與方法
1.1 試驗材料
油樟葉采集于四川省宜賓市高縣月江森林經營所油樟母本園(北緯28°47′77″,東經104°59′76″)的5 個株系(1、14、16、40 和53 號株系),根據區域氣候特點,采樣時間為3月下旬(春季)、6月下旬(夏季)、9月下旬(秋季)和12月上旬(冬季)。采樣時,每個品系隨機選取成年健康油樟3株,每株樹間隔大于20 m,使用單繩攀爬等方式,到達大樹頂端,用滅菌剪剪取東南西北4 個方向的樹葉,每個方向取3條樹枝,選取枝條上端健康完整的樹葉,每個樹枝取10片,混合放入4 ℃冰盒帶回實驗室,用于后續內生真菌分離;枝條剩余葉片全部收集后用于油樟油提取。馬鈴薯葡萄糖瓊脂(potato dextrose agar,PDA)營養瓊脂、麥芽提取物瓊脂、察氏、馬丁、卵磷脂吐溫-80 營養瓊脂、玉米粉瓊脂均為成品培養基,購自北京三藥科技有限公司。
1.2 試驗方法
1.2.1 油樟葉精油的提取 將采集的部分油樟葉放置于室內避光處,待其自然干燥至恒質量后,根據《中華人民共和國藥典 2020 年版 四部》[24]通則2204 揮發油的測定方法進行提取和測量。
1.2.2 油樟葉可培養內生真菌的分離純化 收集健康新鮮的油樟葉于無菌取樣袋中,自來水沖洗組織表面2 h,轉入到滅菌瓶用無菌水沖洗3 次,95%乙醇漂洗1 min,再用無菌水沖洗1 次,4%(有效氯含量)次氯酸鈉漂洗4 min,最后用95%乙醇漂洗1 min,無菌水沖洗5 次,每次30 s。設置漂洗對照、印記對照和環境對照[25]。吸干油樟葉表面水分,利用無菌剪刀將葉片剪成約4 mm2的小葉塊,分別接種到PDA 營養瓊脂、麥芽提取物瓊脂、察氏、馬丁、卵磷脂吐溫-80 營養瓊脂和玉米粉瓊脂培養基上,每皿接種5 片,設置10 個重復,28 ℃恒溫培養。每24 h 觀察菌落情況并計數,根據各平板菌落形態隨機挑選具有代表性的單菌落,連續傳代培養至菌落形態單一。
1.2.3 內生真菌的內轉錄間隔區(internal transcribed spacer,ITS)序列分析 采用真菌通用引物(ITS1:5′-TCCGTAGGTGAACCTGCGG-3′、ITS4:5′-TCCTCCGC TTATTGATATGC-3′)擴增ITS 序列。PCR 反應體系為50 μL:ddH2O 21 μL,2×Taq PCR Master Mix 25 μL,20 μmol·L-1正向引物1 μL,20 μmol·L-1反向引物1 μL,DNA 模板2 μL。PCR 反應條件:94 ℃預變性10 min;94 ℃變性30 s,58 ℃退火30 s,72 ℃延伸45 s,35 個循環;72 ℃終延伸10 min。擴增產物用1%瓊脂糖凝膠電泳檢測,合格后送上海生工生物工程公司進行序列測定。將所得序列信息提交至NCBI 網站(https://www.ncbi.nlm.nih.gov/)BLAST程序進行序列同源性比對。
1.2.4 油樟葉內生真菌群落多樣性分析 選用4 個指標評價油樟葉內生真菌的多樣性,即物種豐富度S、Shannon-Weiner 多樣性指數H′、Simpson 優勢度指數D、均勻度指數E,具體計算公式為:
式中,N為樣本全部屬數之和;Pi為屬相對頻率,指樣本中分離某屬的菌株數占分離總菌株數的百分比。對油樟葉不同株系或季節產油率與葉中可培養內生真菌多樣性指數之間的相關性進行Pearson 相關分析,探究產油率對油樟葉中可培養內生菌群落組成的影響。
1.2.5 油樟葉內生真菌揮發性成分分析 通過氣相色譜/質譜聯用技術(gas chromatograph/mass spectrograph,GC/MS)對內生真菌發酵液進行揮發性成分分析。GC條件為:HP-5毛細管氣相色譜柱(30 m×320 μm×0.25 μm,安捷倫,美國),載氣為高純N2,流量26.56 mL·min-1,程序升溫,起始溫度80 ℃保持3 min,以5 ℃·min-1升溫至90 ℃保持2 min,5 ℃·min-1升溫至170 ℃保持2 min,5 ℃·min-1升溫至220 ℃保持2 min 結束。進樣口溫度240 ℃,進樣量1 μL,不分流進樣,尾吹流量25 mL·min-1。MS條件為:電子轟擊(electron impact,EI)離子源方式,離子源溫度為200 ℃,離子化電壓70 eV,掃描范圍50~650m/z[26]。根據上述分析結果,比較篩選能產與宿主揮發油主要成分相同的菌株。
2 結果與分析
2.1 油樟葉提取的精油含量
由表1可知,選用的5株油樟株系葉產油率存在差異。春季和夏季5 個油樟株系葉產油率均表現為16>40>53>14>1,但秋季表現為53>16>1>14>40,冬季表現為14>16>53>40>1。由季節平均產油率結果可知,春季油樟葉產油率最高,夏季最低,但各指標差異均不顯著。各株系油樟葉產油量以16號最高,1號最低,且差異顯著(P<0.05),表明株系特征是決定油樟葉產油率的關鍵。
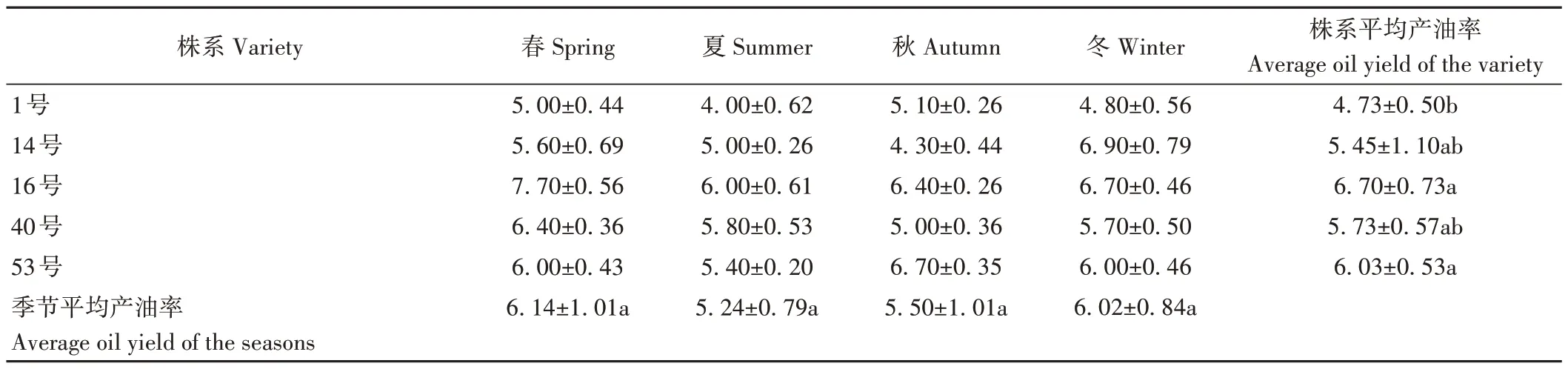
表1 不同株系和季節油樟葉的產油率Table 1 Oil production rate of C.longepaniculatum leaf in different varieties and seasons/%
2.2 油樟葉內生真菌的分離及群落組成分析
從油樟葉分離的菌株經過分離純化共獲得43 株內生真菌(電子附表1),不同株系和季節的內生真菌菌株數存在差異,根據株系分類,1、14、16、40、53 號油樟株系分別分離到8、12、8、10和5株菌株;根據季節分類,春夏秋冬季油樟葉分別分離到7、25、2和9株菌株。經鑒定43 株內生真菌歸屬于2 門、4 綱、9 目、11 科、13屬,其中優勢門為子囊菌門(Ascomycota),優勢綱為座囊菌綱(Dothideomycetes),優勢目為葡萄座腔菌目(Botryosphaeriales)、格孢腔菌目(Pleosporales),優勢科為葉點霉科(Phyllostictaceae),優勢屬為葉點霉屬(Phyllosticta)。不同株系油樟葉內生真菌分布顯示,1 號株系油樟葉分離的8 株菌鑒定為1 門2 綱4 目4 科5 屬,5 個屬包括鏈格孢屬(Alternaria)、斜銼蛤屬(Pseudoplagiostoma)、刺盤孢屬(Colletotrichum)、球座菌屬(Guignardia)和Phyllosticta屬;14號株系油樟葉分離的12 株菌鑒定為1 門3 綱5 目7 科7 屬,7 個屬包括Phyllosticta、Colletotrichum、擬莖點霉屬(Phomopsis)、Pseudoplagiostoma、間座殼屬(Diaporthe)、黑孢霉屬(Nigrospora)和曲霉屬(Aspergillus);16 號株系油樟葉分離的8 株菌鑒定為1 門3 綱5 目5 科6 屬,6 個屬包括枝孢菌屬(Cladosporium)、Guignardia、Phyllosticta、Colletotrichum、Nigrospora和青霉菌屬(Penicillium);40號株系的10株菌鑒定為1門3綱4目4科5屬,5個屬包括Aspergillus、Guignardia、Phyllosticta、Colletotrichum、Pseudoplagiostoma;53號株系油樟葉分離的5株菌鑒定為2 門4 綱5 目5 科5 屬,5 個屬包括Colletotrichum、Phyllosticta、Penicillium、蟻巢傘屬(Termitomyces)和黑管菌屬(Bjerkandera)。根據不同季節油樟葉內生真菌分離結果可知,春季7株菌鑒定為1門2綱3目5科5屬,5 個屬包括Cladosporium、Colletotrichum、Phomopsis、Pseudoplagiostoma和Diaporthe;夏季的25株菌鑒定為1門3 綱3 目3 科5 屬,5 個屬包括Guignardia、Phyllosticta、Aspergillus、Penicillium和Colletotrichum;秋季2 株菌鑒定為1 門2 綱2 目2 科2 屬,分別為Alternaria和Phyllosticta;冬季9株菌鑒定為2門2綱5目5科5屬,5個屬包括Colletotrichum、Pseudoplagiostoma、Nigrospora、Termitomyces和Bjerkandera。
通過圖1可知,5個不同株系油樟葉內生真菌屬類數量差異較小,其中5 株共有屬為Colletotrichum和Phyllosticta。Penicillium和Cladosporium為16 號株系特有屬。表明油樟不同株系葉中可培養內生真菌具有一定相似性,但同時也具備各自多樣性。
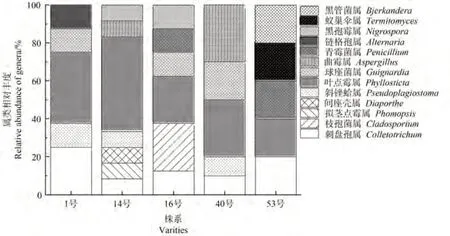
圖1 基于屬水平的不同株系油樟葉內生真菌群落圖Fig.1 Community of endophytic fungi in different varieties of C.longepaniculatum leaves based on genus level
由圖2可知,春夏冬3季屬類數量相同,秋季最少,僅有2屬。春夏秋冬季未見共有屬,春夏秋3季共有屬為Colletotrichum。Cladosporium、Phomopsis、Diaporthe為春季特有屬;Guignardia、Aspergillus、Penicillium 僅出現在夏季;Alternaria僅出現在秋季;Nigrospora、Termitomyces、Bjerkandera僅出現在冬季,可能與不同季節產油率有關。表明季節亦是影響油樟不同株系葉內可培養內生真菌多樣性的主要因素之一。
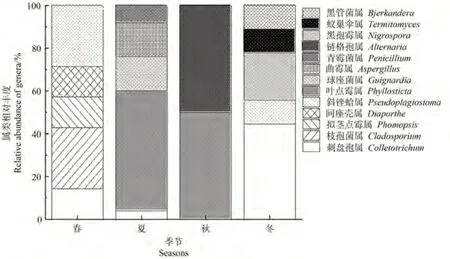
圖2 基于屬水平的不同季節油樟葉內生真菌群落圖Fig.2 Community of endophytic fungi in different seasons of C.longepaniculatum leaves based on genus level
2.3 油樟葉內生真菌的多樣性及相關分析
多樣性分析可顯示宿主植物在生長過程中內生菌群落結構的變化規律,常采用物種豐富度(S)、Shannon-Wienner 指數(H′)、Simpson 指數(D)和均勻度指數(E)等反映多樣性指標。其中H′與物種豐富度關系最密切,數值越大表明物種數量越多,分布越均勻;D數值越大,顯示優勢種越明顯;E敏感性高于H′,用于分析內生真菌群落分布的均勻程度。通過表2和3可知,不同株系油樟葉內生真菌S指數差異不明顯,14 號株系最大,16號株系次之;H′指數表現為16號>53號>14號>40 號>1 號;D指數為16 號>53 號>40 號>1 號>14 號;均勻度指數E為53號株系最大,16號株系次之,14號株系最小。不同季節油樟葉內生真菌S指數春季和夏季值最大,秋季最小;H′指數和D指數均表現為春季>冬季>夏季>秋季;4 季均勻度指數E差異不明顯,冬季最大,秋季最小。由此可知,16 號株系和春季油樟葉內生真菌群落的多樣性相對最豐富,53 號和冬季次之,1號和秋季最低。因此,株系和季節是影響油樟葉中可培養內生真菌多樣性的重要因素。進一步Pearson 相關分析結果發現,不同株系或不同季節油樟葉精油產率與葉中可培養內生真菌多樣性指數無顯著相關關系,揮發油對可培養內生真菌的影響還應做進一步分析。
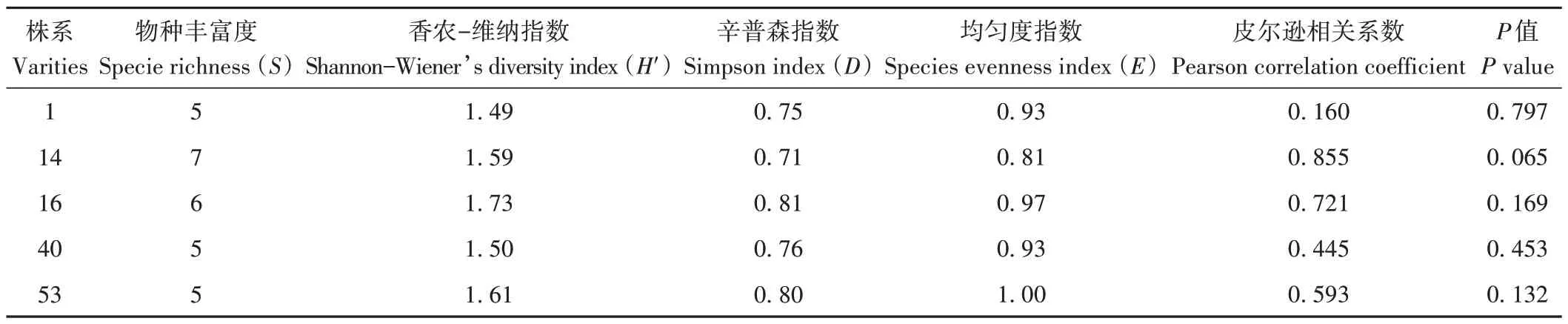
表2 不同株系油樟葉內生真菌的多樣性指數Table 2 Diversity index of endophyte fungi in C.longepaniculatum leaves of different varieties
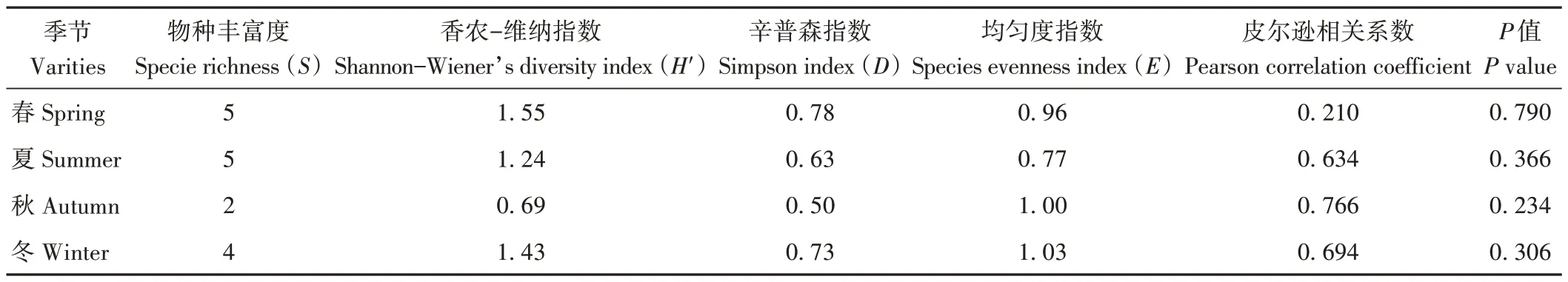
表3 不同季節油樟葉內生真菌的多樣性指數Table 3 Diversity index of endophyte fungi in C.longepaniculatum leaves of different reasons
2.4 產香能力測定
對43 株內生真菌發酵液的GC/MS 分析結果中(表4),共鑒定出69種主要揮發性成分,含醇類化合物13種、酸類化合物11種、酯類化合物11種、烯類化合物6 種、酮類化合物10種、醛類化合物7種、酚類化合物8種和其他類3 種。揮發性成分中有多種產香氣物質,可作為食品添加劑和香料配方,同時也檢測到部分菌株產與油樟精油主要成分相同的物質,如X29 內生真菌發酵液產生的桉葉油醇,相對質量分數為0.39%;D72 發酵液產生的芳樟醇,相對質量分數為0.42%;A39 發酵液產生的紅沒藥醇,相對質量分數為0.97%(表5)。X29和A39均屬于Phyllosticta。

表4 油樟葉內生真菌揮發性成分鑒定結果Table 4 Results of identified volatile components of endophyte fungi from C.longepaniculatum leaves
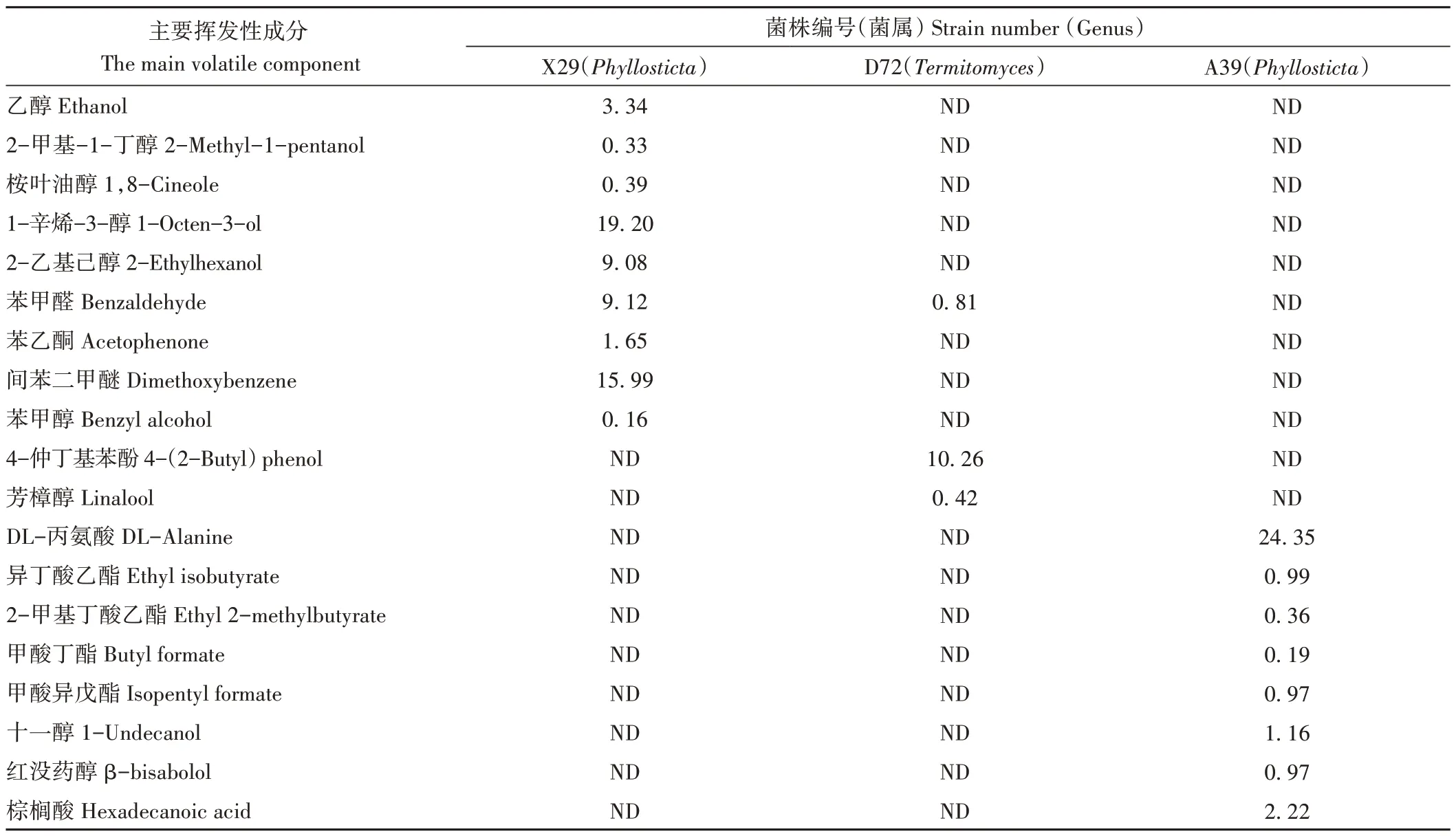
表5 3株產精油成分菌株的主要揮發性物質含量Table 5 Contents of main volatile components in three strains producing essential oil/%
3 討論
內生真菌是參與宿主植物生長、發育及代謝等過程的重要因子,為開發潛在菌株并分析與宿主功能的相關性,采用傳統分離培養法對不同株系和不同季節油樟葉內生真菌進行分離純化和鑒定,可知菌株數量和種屬隨株系和季節的不同而不同,其中16 號株系和春季菌群多樣性相對最豐富,53號和冬季次之,1 號和秋季最低。周寧等[27]發現不同季節蔗葉內生真菌多樣性和分布存在較大差異。呂立新等[28]分析春、夏和秋茅蒼術葉的內生真菌群落分布,表明菌群隨季節的變化具有一定的演替規律,夏季真菌多樣性最高。周萬海等[5]研究了不同季節油樟葉內生細菌的群落組成,發現秋季多樣性種群較豐富且分布均勻,冬季菌株豐富度和均勻度均較低。因此,菌株多樣性可能因宿主生長環境、品種、株齡等不同而不同,且內生真菌和內生細菌具有非一致的變化趨勢。由蔡媛等[29]報道可知多花黃精中分離的內生細菌鑒定為19 門419 屬,而其內生真菌僅鑒定為7 門187 屬。任婷月等[30]采用高通量測序技術對3 個不同品種大麥內生真菌和細菌進行測定,顯示不同品種的內生菌群落組成差異均較大,與本試驗結果相似。內生真菌群落結構與植株狀態、生長和代謝關聯。楊琴等[31]研究顯示不同樹齡銀杏葉的優勢菌門均為子囊菌門,曲霉屬、假絲酵母屬、球腔菌屬等為優勢菌屬。Wu 等[32]研究發現艾蒿葉片內生真菌優勢屬為擔孢酵母屬(Erythrobasidium)和擲孢酵母屬(Sporobolomyces)。本試驗分離的43 株內生真菌優勢門為Ascomycota,優勢屬為Phyllosticta,與前人研究具有較大差異。同時,本實驗室前期利用高通量測序技術對秋季油樟葉片內生真菌類群進行分析,顯示優勢屬為子囊菌門未分類屬(unclassified Ascomycota)、釘孢屬(Passalora)和馬氏霉屬(Uwebraunia)[33];而本研究中來自秋季油樟葉的菌株僅有2 株,隸屬于Alternaria和Phyllosticta,推測是由于受到培養方法、培養基類型和條件局限性所致。
部分內生菌能夠增強宿主植物的健康狀態并促進分泌有效的代謝產物[34],通過與宿主共生誘導能產生與宿主植物相同或相似的代謝產物,如紫錐菊[35]莖葉組織分離的內生細菌可以影響植物揮發性產物和生物堿的合成。暗隔內生菌能促進水稻生長、營養物質的吸收并增強抗逆性[36]。本研究中,雖然春季16號株系油樟葉產油量最高,且內生真菌豐富度、均勻度最佳,但Pearson 相關分析發現,不同株系或不同季節油樟葉精油產率與其中可培養內生真菌多樣性指數無相關性,這與嚴寬等[21-22]的研究結果不一致;上述結果的差異可能與研究材料形態特征不同有關,亦可能與宿主不同株系或環境下含有的特定內生菌誘導產生特定代謝物有關,后續應對不同株系或不同季節油樟葉精油主要成分與內生真菌多樣性進行分析,以確定揮發油含量和成分如何影響其中內生真菌。本研究進一步分析還發現,Penicillium和Cladosporium為16號株系特有屬,Cladosporium、Phomopsis、Diaporthe為春季特有屬,屬于此分類的菌株可能能促進油樟葉精油中特定成分的合成進而提高精油的積累,但相關機制后續還應深入探究。
目前關于菌株的揮發性成分研究主要集中于抗菌性、香氣和病蟲害防治等方面,如楊黎華等[37]從香茅草分離的內生真菌中篩選得到1 株長喙殼屬菌株,其發酵液具有清甜香和花香,利用氣相色譜/質譜儀鑒定出乙縮醛二乙醇、檸檬烯、月桂烯、月桂醛等74種香料物質;馮福山等[38]發現枯草芽胞桿菌Y13 發酵液產生的揮發性物質對油茶炭疽病菌具有較好生防效果;陶玲蕓等[39]發現棘孢木霉揮發性次級代謝產物中6-正戊基-2H-吡喃-2-酮(6-PAP)成分能抑制尖孢鐮孢菌。本試驗從油樟葉中提取的精油具有特殊芳香氣味,通過GC/MS對內生真菌發酵液進行分析鑒定,可知菌株能產生具有醇類、酸類、酯類、烯類、酮類、醛類及酚類等69 種主要揮發性成分。其中苯乙酸乙酯具有濃烈而甜的蜂蜜香氣;甲酸乙酯具有辛辣的刺激味和菠蘿樣的果香香氣;6-戊基-2H-吡喃-2-酮可用于食品用香料,呈蘑菇、藍干酪或乳品香氣;對甲基苯酚呈煙熏、草藥氣味,尤其是桉葉油醇、芳樟醇和紅沒藥醇,它們屬于油樟精油成分,說明油樟內生真菌產生與宿主相同的產物,如X29、D72、A39 菌株,且X29 和A39 均為Phyllosticta,但含量較少,后續將展開進一步條件優化提高產量,并探索其功能。
4 結論
本研究采用傳統微生物純培養法研究了四川宜賓地區特色香料作物油樟葉部內生真菌群落的結構和多樣性,并測定其產油量和菌株發酵液揮發性成分。結果表明,不同株系和不同季節油樟葉的內生真菌在數量、種群及組成上存在差異,油樟葉中可培養內生真菌是一個重要的資源庫,其中16 號株系和春季油樟葉內生真菌群落的多樣性相對最豐富。通過產油率和可培養內生真菌多樣性指數關聯分析,證實兩者無顯著相關關系。此外通過菌株發酵液成分分析,鑒定到醇類、酯類、烯類及酮類等化合物,并顯示菌株能分泌產生精油類代謝產物,具有進一步開發香料成分的潛力。后續應加大對微生物功能的探索,對功能性菌株開展應用研究。
