植物生長調節劑與氮肥對鹽脅迫下水稻幼苗生理特性的影響
2024-02-23 06:52:02王亞新馮乃杰趙黎明鄭殿峰沈雪峰劉美玲杜有為
核農學報 2024年3期
王亞新 馮乃杰, 趙黎明, 鄭殿峰, 沈雪峰, 劉美玲 杜有為
(1廣東海洋大學濱海農業學院,廣東 湛江 524088;2國家耐鹽堿水稻技術創新中心華南中心,廣東 湛江 524088;3廣東海洋大學深圳研究院,廣東 深圳 518108)
水稻(OryzasativaL.)是全世界重要的糧食作物之一。近年來,氣候變化導致海平面上升,從而導致土壤鹽漬化,嚴重威脅水稻生長發育[1]。土壤鹽漬化影響全球20%以上的耕地,其中灌溉區占其一半,這一比例在未來仍將繼續增加[2]。中國有1 億公頃鹽堿地和234萬公頃海灘,預計到2050年,50%以上的可耕地將鹽堿化[3]。鹽脅迫是農業生產的主要制約因素之一,它會阻礙作物生長和發育,降低作物產量,主要表現在形態和生理生化方面。鹽脅迫對植物最直觀的影響是抑制地上部和根系生長,最終導致植物生物量減少[4]。因此,提高作物耐鹽性不僅可以提高鹽堿地利用率,還是保護全球糧食安全的重要保障。相關研究表明,鹽脅迫抑制水稻的光合作用和生長,從而導致生物量減少,最終導致產量下降[5-6]。因此,提高水稻耐鹽性是增加水稻產量的重要途經之一。
化學調控作為預防或緩解非生物脅迫的重要措施之一,在農業生產中得到廣泛應用。相關研究表明,葉面噴施5-氨基乙酰丙酸(5-aminolevulinicacid,5-ALA)具有克服不同作物應激環境有害影響的作用[7],能夠通過增強番茄(SolanumtuberosumL.)幼苗抗氧化系統和在冷脅迫下清除活性氧(reactive oxygen species,ROS)來促進植物生長和光合作用[8],還可以通過減輕丹參(SalviamiltiorrhizaBunge)的氧化損傷來增強其耐鹽性等[9]。乙酸二乙基氨基乙酯(2-diethylaminoethyl hexanoate,DTA-6)是一種促進性植物生長調節劑(plant growth regulators,PGRs),參與水稻作物的生理代謝反應[10-12]。前人研究表明,噴施DTA-6 可以提高決明子(CatsiatoraL.)的耐鹽性[12]和番茄[13]、黑麥(SecalecerealeL.)[14]對鎘脅迫的抗性,并通過促進鹽脅迫下植物的光合作用來促進幼苗生長,緩解鹽脅迫[12,15]。氮素作為作物必需營養元素之一,對水稻的生命活動和產量的形成具有重要意義。然而,過度施肥和施肥不當是我國農業集約化生產的一個嚴重問題[16]。有研究表明,施氮可以緩解鹽脅迫,氮肥通過促進氮有機化合物的積累改善滲透調節、保護細胞結構和功能,從而緩解鹽脅迫對植物的不利影響[17-18]。隨著近年來農田氮肥的過量施用,對環境造成的污染也日益加重。近年來,利用調節劑提高氮素利用率已成為減施增效的有效措施,為解決環境問題提供了新途徑。
目前,關于氮肥和植物生長調節劑對緩解水稻幼苗鹽脅迫影響的對比研究較少,盡管有研究表明5-ALA、DTA-6 和氮肥在改善植物逆境方面具有積極作用[12,19-20],但有關鹽脅迫下5-ALA、DTA-6 和氮肥提高水稻耐鹽性的研究較少,且調節劑與氮肥在改善水稻鹽脅迫中的差異效果仍然未知。基于此,本研究設置兩個氮肥處理,在三葉一心期葉面噴施調節劑,研究其對水稻幼苗形態、膜損傷、抗氧化酶活性及抗氧化物質的影響,以期為氮肥和調節劑在水稻幼苗抗鹽栽培中的應用提供理論依據。
1 材料與方法
1.1 試驗材料
試驗于2021—2022年在廣東海洋大學濱海農業學院日光連棟溫室中進行,采用盆栽試驗,室內溫度白天平均溫度(33±2)℃,夜晚(30±2)℃,濕度控制在80%±5%。供試品種為黃華占,由廣東省農業科學院水稻研究所選育,深圳隆平金谷種業有限公司提供。供試肥料分別為尿素、過磷酸鈣、硫酸鉀。供試調節劑為5-ALA(阿拉丁,北京)和DTA-6(鄭州信聯生化科技有限公司)。試驗用土為磚紅土與河沙3∶1混合均勻。土壤理化性質為:pH值7.02、有機質3.52 g·kg-1、堿解氮1.26 mg·kg-1、速效磷2.62 mg·kg-1、速效鉀83.39 mg·kg-1。
1.2 試驗方法
本試驗為盆栽試驗,每盆3 kg土,試驗各處理詳情見表1,即(1)N1(0.1 g N/盆);(2)N2(0.15 g N/盆);(3)N1A:N1+40 mg·L-15-氨基乙酰丙酸(5-ALA);(4)N1D:N1+30 mg·L-1乙酸二乙基氨基乙酯(DTA-6);(5)N1S:N1+0.3% NaCl;(6)N2S:N2+0.3% NaCl;(7)N1AS:N1+40 mg·L-15-ALA+0.3% NaCl;(8)N1DS:N1+30 mg·L-1DTA-6+0.3% NaCl。
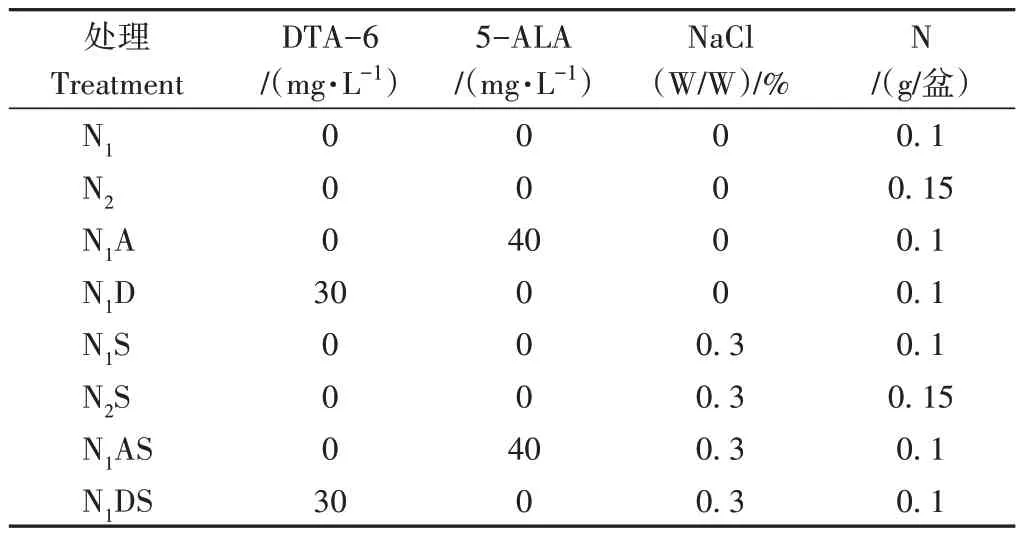
表1 不同處理調節劑、NaCl和N化肥養分用量Table 1 Different treatment regulators,NaCl and nitrogen fertilizer nutrient dosages
選擇飽滿完好的水稻種子,3%雙氧水消毒15 min后,用蒸餾水沖洗5次,在30 ℃恒溫箱中于黑暗條件下蒸餾水浸種和催芽各24 h。隨后取露白一致的種子點播于無孔花盆中,盆的規格為上口徑20 cm、下口徑15 cm、高18 cm,每盆均勻播種69 粒,株距0.5 cm。于播前1 d 澆肥水,每盆施P2O50.48 g、K2O 0.32 g,氮肥(尿素)播前施用,按表1 施入。正常水分管理到幼苗三葉一心期時進行葉面噴施5-ALA和DTA-6,噴施時間為天黑后19:00左右,施用劑量分別為40和30 mg·L-1[21],清水做對照。調節劑處理24 h 后進行鹽處理,即按土施加0.3%NaCl(W/W),將9 g鹽溶于1 L清水中緩慢澆入盆中,模擬大田試驗保留2 cm 的水層。每天用TR-6D 土壤鹽度計(山東恒美科技有限公司)實時監測鹽分,維持鹽濃度。每處理3 次重復,每重復10 盆,共計每處理30 盆,采用完全隨機擺放設計,鹽處理后1、4、7、10、13 d進行取樣用于各項指標的測定。
1.3 測定項目與方法
1.3.1 水稻地上部形態指標的測定 分取各個處理水稻幼苗,用水沖洗干凈,從中選取具有普遍代表性幼苗20 株,地上和地下分開,分別用直尺、游標卡尺測量其苗高和莖基寬。地上部和地下部分樣包裝后放入烘箱于105 ℃殺青30 min、80 ℃烘干48 h 至恒重后,稱量地上和地下干物質量,根據以下公式計算充實度和壯苗指數[22]:
1.3.2 膜 損 傷 指 數 的 測 定 丙 二 醛(malondialdehyde,MDA)含量參照許向陽等[23]的方法采用硫代巴比妥酸法測定。用0.75 mL 上清酶提取液和1.0 mL 硫代巴比妥酸(thiobarbituric acid,TBA)混合測定。將混合物在沸水浴中加熱30 min,冷卻至室溫,10 000 r·min-1離心15 min,得到上清液。分別測定450、532 和600 處的吸光度OD450、OD532、OD600,每個處理3 次重復。根據以下公式計算葉片中丙二醛含量:
過氧化氫含量參照Jessup 等[24]的方法測定。取0.5 g 葉樣品,加入 5 mL 0.1% 三氯乙酸(trichloroaceticacid,TCA)溶液,在液氮中研磨,并以10 000 r·min-1離心10 min。然后將0.5 mL 上清液加入0.5 mL 的10 mmol·L-1磷酸鈉緩沖液(phosphate buffer saline,PBS)和1 mL 碘化鉀(potassium iodide,KI)溶液中,并于28 ℃黑暗反應1 h,使用GENESYS 180 紫外可見光分光光度計(美國賽默飛公司)測定180 nm處的吸光度。
1.3.3 光合色素含量的測定 參考Xu 等[25]的方法,并略有改動。通過鉆孔取樣,并在室溫為30~45 ℃的暗室中用乙醇提取24 h,直到葉片完全變白。然后測定提取液在470、649、665 nm 波長下的吸光度OD470、OD649、OD665,并根據以下公式計算葉綠素a(chlorophyll a,Chl a)、葉綠素b(chlorophyll b,Chl b)、總葉綠素和類胡蘿卜素(carotenoid,Car)含量。每個處理3 次重復。
1.3.4 生理生化指標的測定 按照Lee 等[26]的方法提取粗酶,稱取0.5 g 水稻鮮葉,用10 mL(pH 值7.8)PBS 均質,并在4 ℃下,10 000 r·min-1離心15 min 得到粗酶提取物,每個處理3 次重復。用Macadam 等[27]描述的硝基四唑(nitrotetrazolium blue chloride,NBT)法計算超氧化物歧化酶(superoxide dismutase,SOD)活性,根據Macadam等[27]描述的愈創木酚法測定過氧化物酶(peroxidase,POD)活性,根據Li 等[28]描述的過氧化氫法測定過氧化氫酶(catalase,CAT)活性,根據Nakano 等[29]描述的方法測定抗壞血酸過氧化物酶(ascorbate peroxidase,APX)活性。根據Ellman[30]描述的方法測定還原型谷胱甘肽(glutathione,GSH)含量,參考Bradford[31]的方法測定可溶性蛋白含量。
1.4 數據處理
采用Microsoft Excel 2010 進行數據處理,SPSS X9軟件進行單因素或雙因素方差分析(ANOVA),多重比較采用鄧肯多重檢驗(Duncan’s Test),用Origin 2021軟件繪制圖表。
2 結果與分析
2.1 PGRs和施氮量對水稻幼苗形態的影響
2.1.1 PGRs 和施氮量對苗高、莖基寬和地上干重的影響 由表2 可知,調節因子(Re)主效應和鹽濃度×調節因子(Sc×Re)互作均顯著或極顯著影響了苗高、莖基寬和地上干重。NaCl 脅迫整體降低了苗高、莖基寬和地上干重,其中NaCl 脅迫后1~13 d,苗高、莖基寬和地上干重表現為,N1S 處理較N1分別下降2.26%~7.92%、4.67%~15.96%和12.21%~25.12%,N2S 處理較N2處理分別下降1.24%~4.78%、2.88%~12.82%和5.69%~19.61%,說明鹽脅迫抑制了水稻幼苗生長。從氮肥角度看,增施氮肥能夠促進幼苗生長,尤其在NaCl 脅迫下,N2S 處理苗高、莖基寬和地上干重分別較N1S 處理提高3.23%~20.76%、7.94%~14.78% 和17.18%~39.30%,這說明增施氮肥能夠緩解NaCl 脅迫對水稻幼苗生長的毒害作用。從PGRs 角度看,在鹽脅迫后1~13 d,N1A 處理苗高和地上干重分別較N1顯著增加6.29%~22.71% 和15.37%~41.02%,N1AS處理苗高、莖基寬和地上干重分別較N1S 處理顯著提高12.61%~27.59%、19.61%~32.91% 和31.08%~78.95%,其次是N1DS 處理也顯著提高了苗高、莖基寬和地上干重,說明噴施兩種PGRs 不僅對幼苗形態生長有促進作用,還能夠緩解鹽脅迫帶來的傷害。在NaCl 脅迫4 d 時,N1AS 和N1DS 處理各指標較N2S 處理增幅較大,苗高分別顯著增加20.00%和24.00%、莖基寬分別顯著增加19.43%和18.40%、地上干重分別顯著增加36.82%和29.38%。兩種PGRs 在促進水稻幼苗生長,特別是改善鹽脅迫對水稻的損傷中均能達到增施定量氮肥效果。
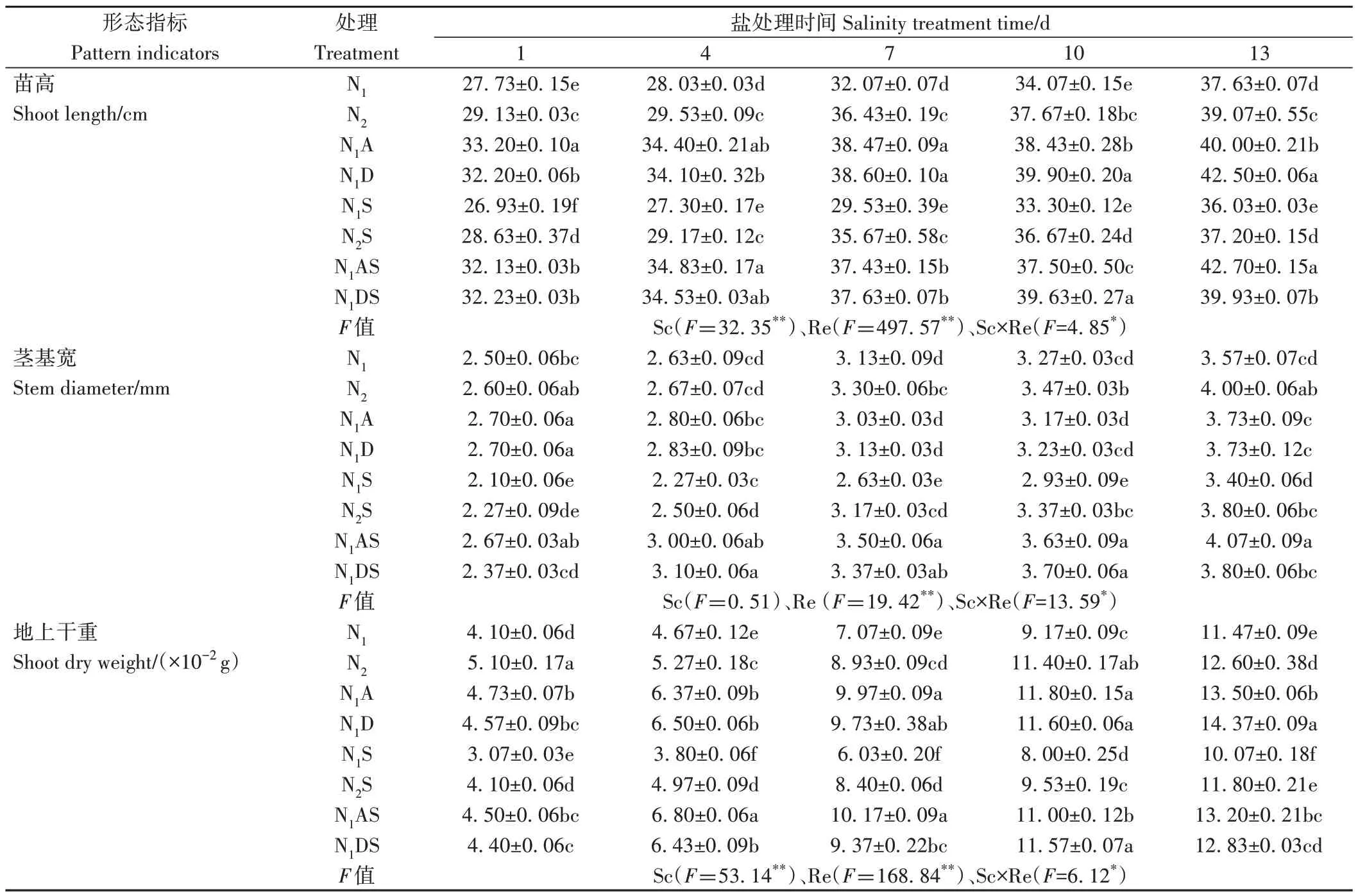
表2 PGRs和施氮量對NaCl脅迫下水稻幼苗的影響Table 2 Effects of PGRs and nitrogen application on rice seedlings under NaCl stress
2.1.2 PGRs和施氮量對水稻幼苗充實度和壯苗指數的影響 由圖1 可知,NaCl脅迫后1~13 d,充實度和壯苗指數表現為,N1S處理較N1分別下降7.35%~22.97%和10.53%~23.38%;N2S 處理較N2處理分別下降4.50%~18.20%和7.71%~12.14%。說明NaCl脅迫抑制了水稻生長。從氮肥角度看,增施氮肥能夠促進幼苗生長,尤其在NaCl脅迫下,N2S處理充實度和壯苗指數較N1S 處理整體分別顯著提高8.23%~25.76%和7.84%~44.71%,這說明增施氮肥能夠緩解水稻NaCl脅迫。從PGRs角度看,在NaCl脅迫后4~13 d,N1A處理充實度和壯苗指數分別較N1處理顯著提高10.77%~17.57%和29.17%~69.31%,N1AS、N1DS 處理充實度和壯苗指數分別較N1S 處理提高10.65%~40.26%和9.80%~66.47%、15.04%~33.98%和41.18%~75.23%,說明噴施兩種PGRs 不僅對幼苗形態生長有促進作用,還能夠緩解NaCl 脅迫帶來的傷害。從綜合角度看,在NaCl脅迫13 d 時,N1AS 和N1DS 處理壯苗指數較N2S 處理增幅較大,分別增加43.33%和36.93%。說明兩種PGRs 改善NaCl脅迫下水稻生長效果均優于增施氮肥效果,以5-ALA效果最佳。
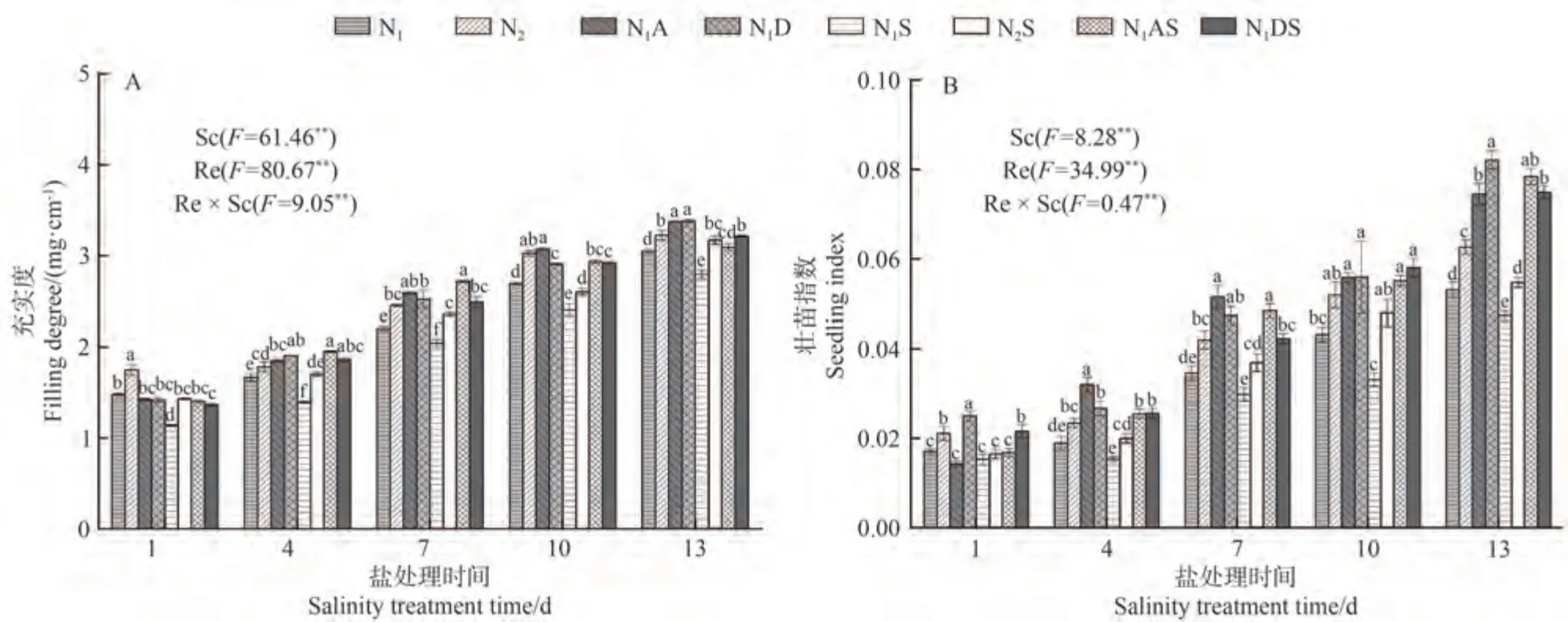
圖1 PGRs和施氮量對水稻幼苗充實度和壯苗指數的影響Fig.1 Effects of PGRs and nitrogen application on filling degree and seedling index of rice seedlings
2.2 外源5-ALA、DTA-6 和氮肥對水稻幼苗光合色素含量的影響
由表3 可知,Sc、Re 主效應和Sc×Re 互作均極顯著影響了水稻葉片葉綠素a、葉綠素b和總葉綠素含量(P<0.01)。首先,NaCl 脅迫影響幼苗的光合色素合成。NaCl 處理1~13 d,與N1相比,N1S 處理水稻葉片的葉綠素a、葉綠素b、總葉綠素和類胡蘿卜素含量分別下降1.25%~25.50%、9.45%~27.29%、3.86%~25.45% 和5.67%~21.36%;NaCl處理1~10 d,與N2相比,N2S處理的葉綠素a、葉綠素b、總葉綠素和類胡蘿卜素含量分別下降2.70%~26.98%、4.79%~47.91%、9.09%~24.68%和7.48%~17.25%。說明NaCl 脅迫抑制了幼苗的光合色素合成。其次,在氮肥利用效率上,NaCl 脅迫處理1~13 d,N2S 處理的葉綠素a、葉綠素b、總葉綠素和類胡蘿卜素含量分別較N1S處理增加4.32%~29.41%、4.67%~34.39%、4.43%~29.34% 和4.01%~26.46%,這說明增施氮肥能夠促進NaCl 脅迫下葉片光合色素合成。再次,從PGRs調控效果看,N1A處理的葉綠素a、葉綠素b、總葉綠素和類胡蘿卜素含量分別較N1處理增加4.21%~15.37%、6.05%~89.66%、9.90%~29.97%和1.66%~52.74%;N1AS 處理的葉綠素a、葉綠素b、總葉綠素和類胡蘿卜素含量分別較N1S 處理增加13.06%~35.31%、24.31%~197.63%、15.90%~69.35%和14.82%~44.24%,N1DS 處理葉綠素a、葉綠素b、總葉綠素和類胡蘿卜素含量則分別增加18.42%~42.45%、2.76%~129.26%、15.11%~60.65%和4.19%~45.41%。說明5-ALA 和DTA-6 均能夠緩解NaCl 脅迫對水稻幼苗葉片光合色素合成的抑制,以葉面噴施5-ALA 調控效果最佳。N1AS 和N1DS 處理總葉綠素含量較N2S 處理在NaCl 脅迫13 d 時的增幅較大,分別增加30.93%和24.21%。說明兩種PGRs 在減小NaCl 脅迫下光合色素降解速度方面的效果均強于增施定量氮肥,這有利于逆境條件下水稻植株進行光合作用,減輕鹽脅迫對水稻幼苗的傷害。
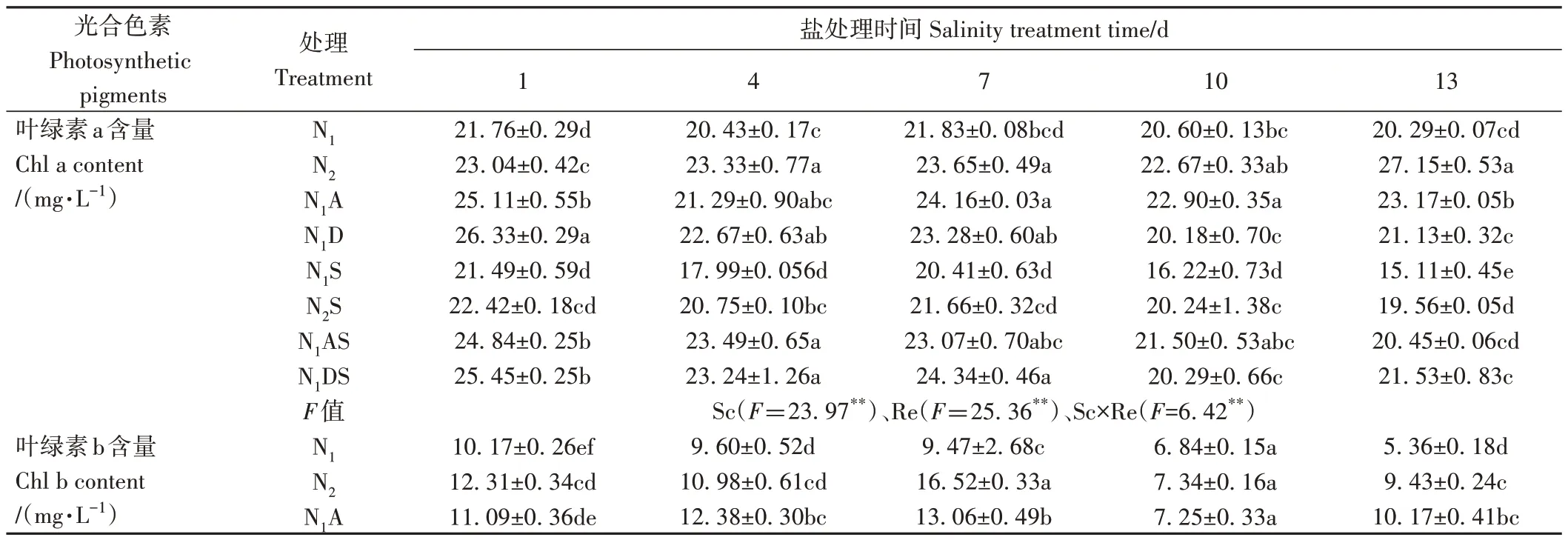
表3 PGRs和施氮量對水稻幼苗光合色素含量的影響Table 3 Effects of PGRs and nitrogen application on photosynthetic pigment content of rice seedlings
2.3 PGRs和施氮量對水稻幼苗膜損傷程度的影響
由表4 可知,Sc、Re 主效應和Sc×Re 互作均極顯著影響了水稻葉片MDA 和H2O2含量(P<0.01)。NaCl 脅迫處理1~13 d,與N1相比,N1S 處理丙二醛和過氧化氫含量分別顯著增加5.75%~22.11%和7.45%~16.39%;與N2相比,N2S 處理上述指標分別顯著增加5.97%~19.54%和3.67%~11.62%,這說明NaCl 脅迫使葉片產生過量活性氧,引起膜脂過氧化物MDA 的積累,破壞細胞膜,阻礙水稻生長。而增施氮肥能夠維持葉片活性氧的產生和清除的動態平衡,尤其在NaCl脅迫處理4~13 d,與N1S 處理相比,N2S 處理的丙二醛和過氧化氫含量分別顯著降低17.93%~36.54%和10.11%~20.95%,這說明增施氮肥可以有效提高水稻抗逆性。
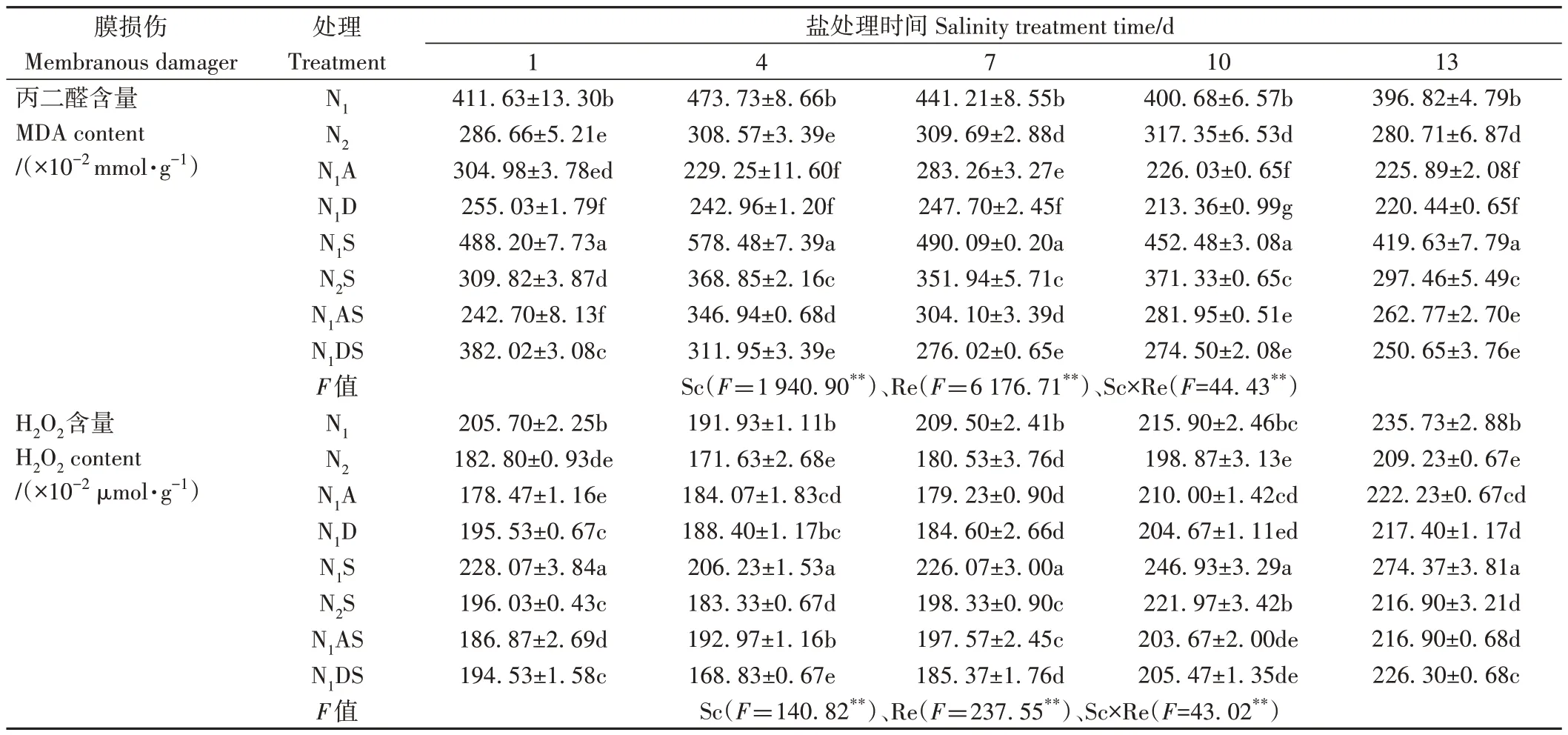
表4 PGRs和施氮量對水稻幼苗膜損傷程度的影響Table 4 Effects of PGRs and nitrogen application on the degree of membrane damage of rice seedlings
外源施加兩種PGRs 對水稻幼苗有一定的調控效果。N1A 處理MDA 和過氧化氫含量整體分別較N1處理顯著降低25.90%~51.61%和4.10%~14.45%,N1AS處理丙二醛和過氧化氫含量分別較N1S 處理顯著降低37.38%~50.29%和6.43%~20.95%,N1DS 處理的丙二醛和過氧化氫較N1S 處理分別降低21.75%~46.07%和14.71%~18.14%,這說明外源施加兩種PGRs 均可減少葉片ROS積累,維持細胞膜穩定性。在NaCl脅迫10 d時,N1AS和N1DS處理MDA含量較N2S處理降幅較大,分別降低24.07%和26.08%;H2O2含量則分別降低8.24%和7.43%。說明兩種PGRs 在緩解水稻膜損傷方面的效果均優于增施定量氮肥。
2.4 PGRs和施氮量對水稻葉片抗氧化物質的影響
2.4.1 對水稻葉片SOD 活性的影響 由圖2-A 可知,鹽脅迫影響水稻葉片SOD 活性,調節因子可以改善鹽脅迫對水稻的傷害。隨NaCl 脅迫時間的延長,葉片SOD活性呈先上升后下降的趨勢。NaCl脅迫處理第1、第4 和第7 天,N1S 處理SOD 活性分別較N1處理顯著增加16.32%、12.65%和9.82%;N2S 處理上述指標分別較N2增加6.66%、6.63%和3.92%。NaCl 脅迫處理第13 天,與N1、N2處理相比,N1S、N2S 處理SOD 活性分別顯著下降13.72%、6.09%。表明在NaCl 脅迫下,水稻先產生應激反應,SOD 活性增加。隨著脅迫時間的增加,抗氧化系統遭到破壞,SOD活性降低。從氮肥角度看,增施氮肥可以增加SOD 活性,尤其在NaCl 脅迫處理4~13 d,N2S 處理SOD 活性較N1S 處理顯著增加5.21%~17.18%,表明施加氮肥可以提高NaCl 脅迫下水稻葉片SOD 活性。從PGRs 角度看,N1A 和N1D 處理的SOD 活性分別較N1處理顯著增加7.30%~21.11%和5.49%~28.68%,N1AS 處理和N1DS 處理分別較N1S處理增加15.59%~23.60%和6.24%~20.86%,說明兩種PGRs 均可以提高NaCl 脅迫下水稻葉片的SOD 活性。從綜合角度看,N1AS 和N1DS 處理SOD 活性較N2S處理在NaCl 脅迫13 d 時的增幅較大,分別增加11.01%和8.55%。說明兩種PGRs 對NaCl 脅迫下水稻葉片SOD活性的提升效果優于增施定量氮肥。
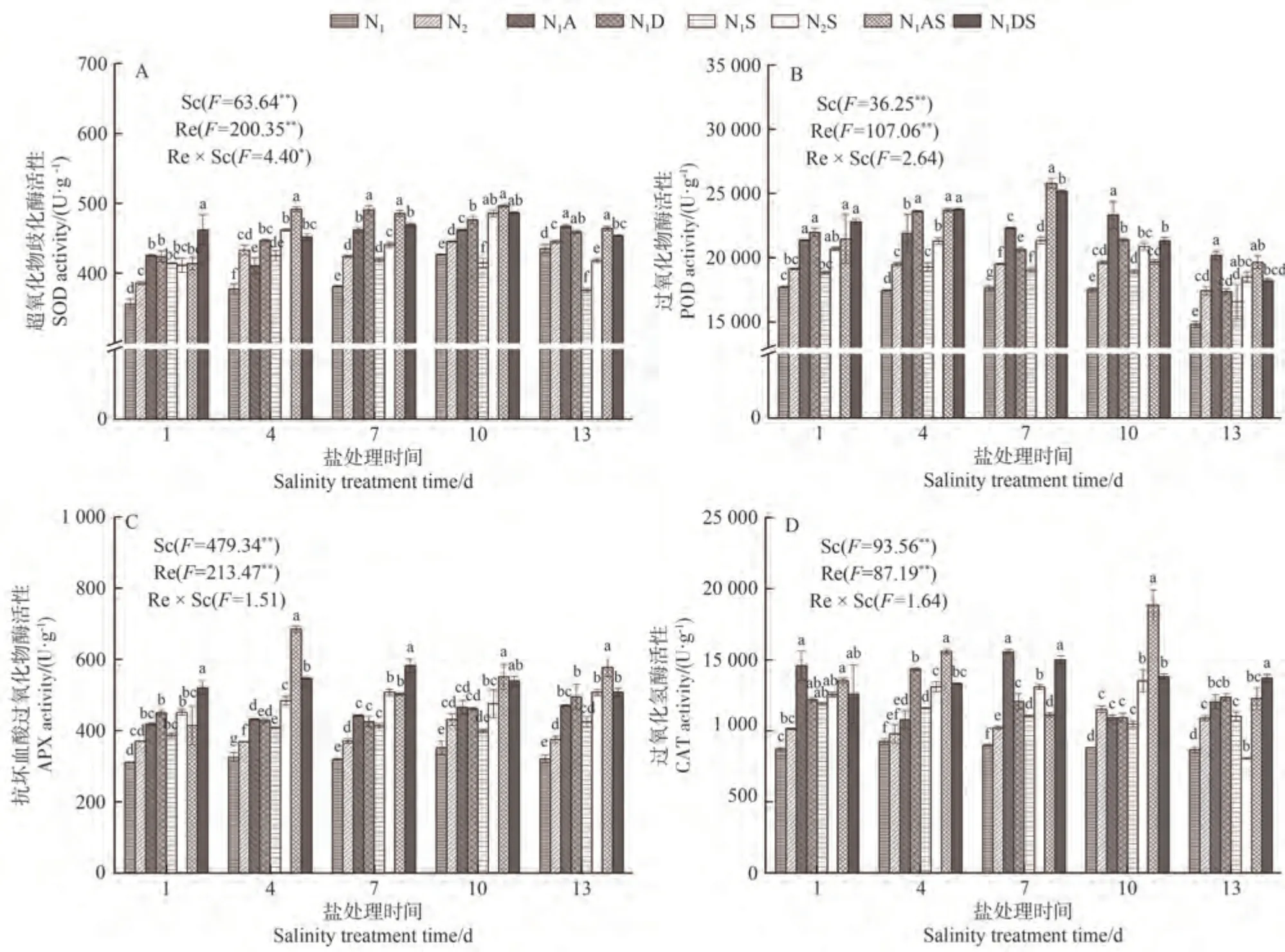
圖2 PGRs和施氮量對水稻葉片抗氧化物質的影響Fig.2 Effects of PGRs and nitrogen application on antioxidant substances in rice leaves
2.4.2 對水稻葉片POD 活性的影響 由圖2-B 可知,NaCl 脅迫使葉片POD 活性增加。NaCl 脅迫處理第1~第13 天,N1S 處理POD 活性較N1整體顯著增加6.31%~11.94%;N2S 處理POD 活性較N2增加6.15%~9.39%。表明在NaCl 脅迫下,水稻先產生應激反應,POD 活性增加。增施氮肥可以增加POD 活性,尤其在NaCl脅迫處理4~13 d,N2S處理的POD 活性較N1S處理整體顯著增加10.37%~12.25%,這說明施加氮肥可以提高NaCl 脅迫下水稻葉片的POD活性。從PGRs調控效果分析,N1A和N1D處理POD活性較N1處理分別顯著增加20.63%~36.25%和17.04%~35.27%,N1AS和N1DS處理POD 活性較N1S 處理分別增加了4.07%~35.64%和9.81%~32.42%,說明兩種PGRs 均可以提高NaCl脅迫下水稻葉片的POD 活性。N1AS 和N1DS 處理POD活性較N2S處理在NaCl脅迫7 d時的增幅較大,分別增加20.84%和17.97%。說明兩種PGRs 提高NaCl 脅迫下水稻葉片POD活性的效果優于增施定量氮肥。
2.4.3 對水稻葉片APX活性的影響 由圖2-C可知,NaCl 脅迫使葉片APX 活性增加。NaCl 脅迫處理第1~第13 天,N1S 處理的APX 活性較N1增加13.35%~32.34%;N2S 處理APX 活性較N2增加10.42%~35.25%。表明在NaCl 脅迫下,水稻先產生應激反應,APX 活性增加。從氮肥角度看,增施氮肥可以增加APX 活性,尤其在NaCl 脅迫處理4~13 d,N2S 處理APX活性較N1S 處理顯著增加19.38%~22.85%,這說明施加氮肥可以提高NaCl 脅迫下葉片APX 活性。從PGRs角度看,N1A 和N1D 處理的APX 活性分別較N1處理顯著增加32.40%~46.77%和31.07%~54.41%,N1AS 和N1DS處理APX活性分別較N1S 處理顯著增加21.53%~68.06%和19.79%~40.99%,說明兩種PGRs 均可以提高NaCl 脅迫下葉片的APX 活性,以5-ALA 效果最佳。從綜合角度看,N1AS和N1DS處理APX 活性較N2S處理在NaCl 脅迫4 d 時的增幅較大,分別顯著增加41.63%和13.01%。說明兩種PGRs 對NaCl 脅迫下水稻葉片APX活性的提升效果優于增施定量氮肥。
2.4.4 對水稻葉片CAT 活性的影響 由圖2-D 可知,NaCl脅迫下的CAT 活性有所增加。NaCl脅迫處理第1~第10 天,N1S 處理的CAT 活性較N1顯著增加18.44%~36.15%;N2S 處理CAT 活性較N2顯著增加17.33%~33.71%。表明在NaCl脅迫下,水稻產生應激反應,CAT 活性增加。氮肥方面,增施氮肥可以增加CAT 活性,尤其在NaCl 脅迫處理1~10 d,N2S 處理CAT活性較N1S處理增加5.70%~29.00%,這說明施加氮肥可以提高NaCl脅迫下葉片CAT 活性。PGRs方面,N1A和N1D處理CAT活性較N1處理分別顯著增加16.60%~73.31%和23.72%~55.05%,N1AS和N1DS處理CAT 活性分別較N1S 處理增加0.95%~79.95% 和5.84%~31.94%,說明兩種PGRs 均可提高NaCl 脅迫下葉片的APX 活性。整體來看,N1AS 和N1DS 處理CAT 活性較N2S 處理在NaCl 脅迫10 d 時的增幅較大,分別增加39.49%和2.27%。說明兩種PGRs 對NaCl 脅迫下水稻葉片CAT活性的提升效果優于增施定量氮肥。
2.5 PGRs 和施氮量對水稻葉片可溶性蛋白和GSH含量的影響
由圖3-A可知,Sc、Re主效應和Sc×Re交互作用均對葉片的可溶性蛋白含量有極顯著影響(P<0.01)。NaCl 脅迫下,與N1和N2相比,N1S 和N2S 處理的可溶性蛋白含量均呈上升趨勢。與N1相比,N1S 處理可溶性蛋白含量在NaCl 脅迫處理第1~第13 天增加1.82%~10.11%;在NaCl脅迫處理第1~第13天,N2S處理可溶性蛋白含量較N2增加2.16%~9.76%。表明NaCl 脅迫使水稻產生應激反應,可溶性蛋白含量增加。從氮肥角度看,增施氮肥可以增加可溶性蛋白含量,在NaCl脅迫處理1~13 d,N2S 處理較N1S 處理增加2.25%~4.38%。從PGRs角度看,N1A和N1D處理的可溶性蛋白含量較N1處理分別顯著增加5.75%~13.24%和8.74%~10.66%,N1AS 和N1DS 處理可溶性蛋白含量較N1S 處理分別增加4.58%~7.61%和4.68%~8.09%,說明兩種PGRs 均能促進葉片滲透調節物質的產生,維持細胞穩態。
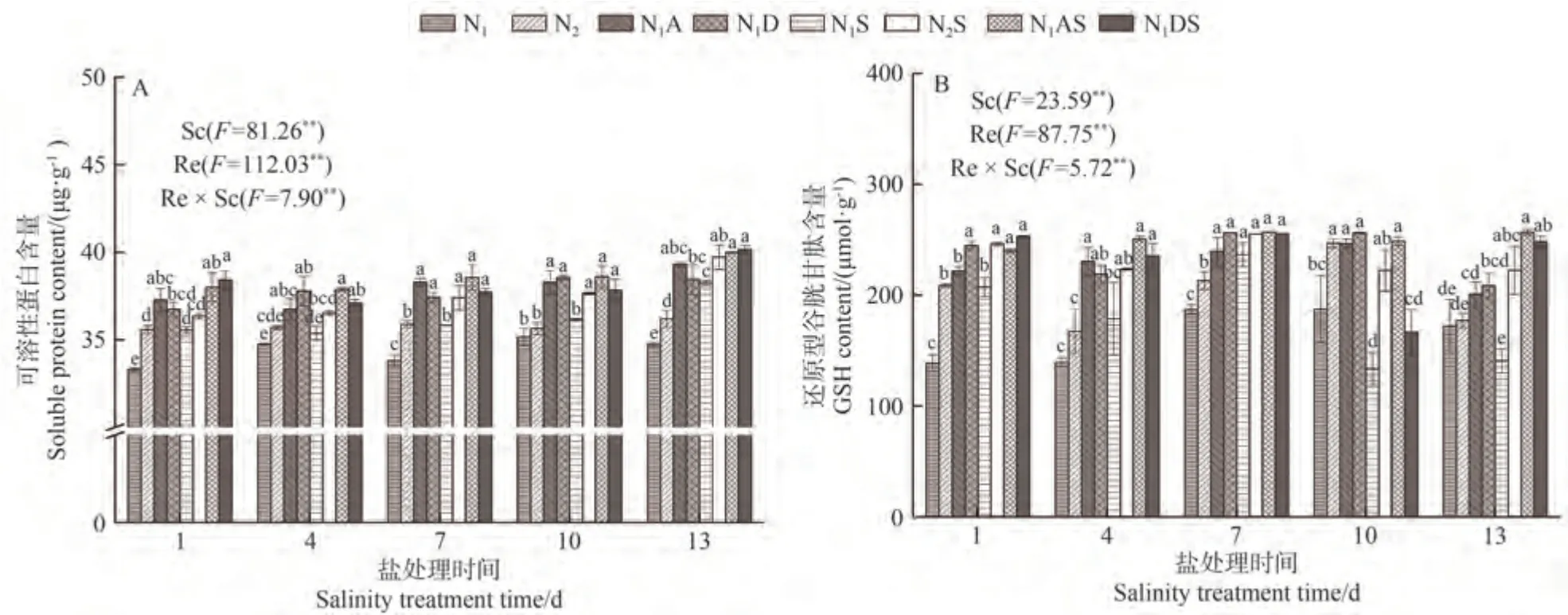
圖3 PGRs和施氮量對水稻葉片可溶性蛋白含量和GSH含量的影響Fig.3 Effects of PGRs and nitrogen application on soluble protein and GSH content of rice leaves
由圖3-B可知,Sc、Re主效應和Sc×Re交互作用均對葉片的GSH 含量有極顯著影響(P<0.01)。NaCl 脅迫下,與N1相比,N1S處理GSH 含量呈先增加再降低趨勢。與N1和N2相比,N1S 和N2S 處理CSH 含量在NaCl脅迫處理第1~第7天分別相應增加26.47%~49.63%和18.10%~33.41%;與N1相比,N1S 處理CSH 含量在NaCl脅迫第10~第13天降低18.18%~28.81%。這表明NaCl 脅迫使水稻先產生應激反應,抗氧化劑GSH 含量增加,隨著脅迫時間增加,脅迫程度加劇,GSH 含量降低。從氮肥角度看,增施氮肥可以促進GSH 合成,在NaCl脅迫處理1~13 d,N2S處理GSH 含量較N1S處理增加7.77%~58.14%。PGRs 調控效果顯示,N1A 和N1D處理的GSH 含量較N1處理分別增加16.82%~65.44%和21.22%~76.68%,N1AS 和N1DS 處理較N1S 處理分別增加8.58%~86.42%和8.02%~76.52%,表明兩種PGRs 均能提高水稻抗氧化脅迫能力。從綜合角度分析,N1AS 和N1DS 處理GSH 含量較N2S 處理在NaCl 脅迫13 d 時的增幅較大,分別提高15.50%和11.62%。說明在NaCl 脅迫下,兩種PGRs 對水稻葉片GSH 合成的促進效果均優于增施定量氮肥。
2.6 各指標間的相關性
本研究對17 個指標的相關性進行分析(圖4),結果表明,水稻的形態指標(株高、莖基寬、地上干重、充實度及壯苗指數)均與葉綠素a、葉綠素b、可溶性蛋白和GSH含量呈顯著或極顯著正相關,與MDA含量呈顯著或極顯著負相關。GSH 含量與SOD、POD、APX 及CAT 活性呈極顯著正相關,MDA 含量與POD 活性呈極顯著負相關。
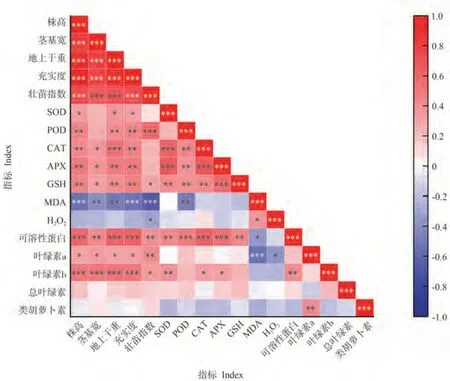
圖4 所測指標間的相關性分析Fig.4 Correlation analysis between measured indicators
3 討論
3.1 PGRs和施氮量對水稻幼苗建成的影響
NaCl 脅迫會抑制植物生長,降低植物生物量,在甜高粱[32]、小麥[33]和水稻[34-35]等多種作物中均有報道。本研究表明,隨著NaCl 脅迫時間的延長,幼苗形態指標均呈現下降趨勢,說明NaCl 脅迫抑制幼苗生長。在NaCl 脅迫下,氮肥和葉面噴施兩種PGRs 均能提高幼苗的耐鹽性,增加莖基寬、壯苗指數和地上干重,達到壯苗效果,一定程度上緩解NaCl 脅迫。這與前人研究氮肥、5-ALA 和DTA-6 均可促進逆境中植物生長的結果一致[36]。總體看來,在低氮基礎上葉噴兩種GPRs 對水稻幼苗形態的影響均達到在低氮基礎上每盆增施0.05 g純氮效果。
3.2 PGRs和施氮量對水稻幼苗光合色素含量的影響
NaCl 脅迫會破壞葉綠體結構,降低葉綠素含量,導致光合作用減弱,影響植物生命活動[32]。光合色素可以穩定類囊體膜以減少光氧化損傷[37]。本研究顯示,在NaCl 脅迫下,水稻葉片的葉綠素和類胡蘿卜素含量隨著鹽脅迫時間的延長呈下降趨勢,這可能是因為NaCl脅迫減弱光合色素合成酶活性,損壞葉綠體結構,從而加速光合色素的分解。5-ALA、DTA-6和氮肥均調節了NaCl脅迫下光合色素的含量,維持葉片光合作用光反應過程,這與前人有關5-ALA、DTA-6 和氮肥提高葉綠素含量和光合能量轉換效率的結果相一致[38-40]。本研究表明,NaCl 脅迫下,葉噴兩種調節劑處理光合色素含量高于增施定量氮肥,其中5-ALA 效果優于DTA-6,這可能是因為DTA-6是通過提高δ-氨基乙酰丙酸(δ-aminolevulinic acid,ALA)向膽色素原(porphobilinogen,PBG)及PBG 向尿卟啉原Ⅲ的轉化效率來促進葉綠素合成,而5-ALA 是所有卟啉化合物生物合成的共同前體,是植物體內的非蛋白型氨基酸,直接參與葉綠素合成,顯著改善光合色素積累[41]。
3.3 PGRs和施氮量對水稻活性氧代謝的影響
NaCl 脅迫使植物體內活性氧增加,活性氧過度積累造成氧化損傷[42-43]。MDA 是膜脂過氧化的重要指標,其含量可以反映逆境條件下的膜損傷程度[44]。H2O2在植物代謝和信號通路中發揮重要作用,并影響作物生長發育[45]。NaCl 脅迫下,抗氧化系統通過清除植物體內過量ROS 來維持氧化還原穩態[45-46]。超氧化物歧化酶和過氧化物酶的活性常被作為衡量植物抗逆性的兩大指標[47]。本研究發現,NaCl 脅迫下,鹽處理使MDA和H2O2含量增加,SOD和CAT活性隨著脅迫時間的延長先增加后減小,POD 和APX 活性則逐漸升高,與前人研究結果一致[38,48]。這可能是因為水稻幼苗主動調節自身保護酶活性來消除活性氧自由基帶來的傷害。氮肥、5-ALA和DTA-6均能降低NaCl脅迫引起的超氧自由基積累,從而減輕幼苗氧化損傷,維持其正常生長。這與前人有關氮肥、5-ALA 和DTA-6 可以緩解逆境引起的膜損傷的結果一致[40]。其中DTA-6對膜損傷的緩解效果優于5-ALA,5-ALA 提高植物保護酶活性效果最佳,這可能是由于5-ALA 作為抗氧化酶活性的啟動子[49],刺激抗氧化酶活性增加。
可溶性蛋白含量可以反映植物整體代謝水平,非生物脅迫對植物蛋白質合成影響較大[50]。NaCl 脅迫使MDA 和可溶性蛋白含量增加,說明NaCl 脅迫在損傷水稻葉片細胞膜的同時,提高了滲透調節物質的積累以適應NaCl 脅迫。原型谷胱甘肽是一種強非酶抗氧化劑[51],本研究表明,5-ALA、DTA-6 和氮肥均可促進水稻葉片GSH 合成。在NaCl 脅迫下,水稻葉片中GSH 含量隨著NaCl 脅迫時間的延長呈先升高后降低趨勢,可能是由于GSH 消耗降低了植物細胞中的ROS[52]。在NaCl 脅迫下,5-ALA、DTA-6 和氮肥均使GSH 含量增加且增幅相似,說明在氮肥減施的基礎上葉噴兩種PGRs 清除鹽脅迫下水稻葉片細胞內自由基和過氧化物能力與增施氮肥相當。
相關性分析表明,POD 活性與MDA 含量呈顯著負相關,推測NaCl 脅迫下,水稻本身的POD 不能清除過量的活性氧,需要靠調節因子的調控作用緩解氧化損傷。GSH含量與SOD、POD、APX及CAT活性呈極顯著正相關,這可能是由于氮肥和PGRs 提高了抗氧化酶活性,調控GSH 的合成,保護NaCl 脅迫下水稻細胞膜結構的完整性,與前人研究報道中調節劑保護堿脅迫下煙草細胞膜結構完整性的結果相一致[53]。然而,磚紅壤養分含量很低,本試驗用土采用的是貧瘠的磚紅壤與沙3∶1混合,土壤養分含量相對較低,雖然氮水平N2較N1增幅達50%,但有關進一步增加施氮量或改為其他養分含量較高的土壤是否有同樣的試驗效果還有待進一步探索。
4 結論
NaCl 脅迫抑制水稻生長和代謝過程。增施氮肥與葉面噴施5-ALA 和DTA-6 均可改善NaCl 脅迫期間水稻幼苗的形態變化,提高光合色素含量和抗氧化水平,從而減輕活性氧對幼苗的損傷。施用調節劑(5-ALA和DTA-6)對減輕幼苗在NaCl脅迫下受到的氧化應激損傷效果優于增施定量氮肥,以5-ALA 對NaCl脅迫下幼苗的形態和抗氧化酶活性改善效果較好,DTA-6 在緩解NaCl 脅迫下水稻幼苗膜損傷方面的效果較好。說明在緩解NaCl脅迫對水稻的損傷中,調節劑可以達到增施定量氮肥的效果,進而推測在低氮基礎上外源施用兩種PGRs可以減少水稻氮肥施用量。
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中老年保健(2021年12期)2021-11-30 02:58:01
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
攝影之友(影像視覺)(2019年2期)2019-03-05 08:27:14
中華詩詞(2018年11期)2018-03-26 06:41:34
Coco薇(2016年8期)2016-10-09 02:11:50
