不同地面覆蓋方式下新郁葡萄果皮黃酮轉錄組和代謝組聯合分析
2024-02-23 09:30:00戶金鴿白世踐蔡軍社
新疆農業科學 2024年1期
戶金鴿,白世踐,陳 光,蔡軍社
(新疆維吾爾自治區葡萄瓜果研究所,新疆鄯善 838200)
0 引 言
【研究意義】新郁葡萄是以紅地球自然雜交單珠E-42-6為母本,里扎馬特為父本雜交選育而成的鮮食葡萄,橢圓形,果皮紫紅色,平均單粒質量11.6 g,果肉較脆,味酸甜,可溶性固形物16%~19%,耐貯運性較好,適應性較強[1]。在新疆吐魯番市新郁葡萄成熟季節正值夏季高溫,造成果實著色不均或過深,影響了果實的外觀品質。黃酮類化合物作為植物和水果等自身產生的一類次生代謝物質[2-3],能賦予植物紅色、黃色、橙色、紫色以及藍色等多種顏色[4],是園藝植物重要的農藝性狀,是判斷果實成熟、外觀品質的重要指標之一[5]。【前人研究進展】地面覆蓋技術具有改良土壤結構、蓄水保墑、調節微域生態環境等生態功能[6]。地面覆蓋可顯著降低空氣溫濕度及土壤溫度,提高果實總酸含量、可溶性固形物和還原糖含量、花色苷、酚類物質和單寧含量等外在品質和內在品質[7-9],調節果實糖酸比及釀酒葡萄香氣物質的積累[10]。代謝組學是將生物體作為一個動態整體,與轉錄組學、蛋白質組學等數據結合分析更能直接、真實反映機體本身的變化水平[11]。黃酮合成途徑主要包括苯丙氨酸代謝途徑、類黃酮合成代謝途徑以及各種花青素的合成等[12],所涉及的結構基因主要有苯丙氨酸解氨酶(PAL,14個成員)、查爾酮合成酶(CHS,35個成員)、黃烷酮3′羥化酶(F3′H,6個成員)、黃烷酮3′5′羥化酶(F3′5′H,13個成員),調節基因主要有MYB、bHLH、WD40、WKRY、bZIP、BBX[13]。與類黃酮合成關系密切的13個基因中,肉桂酸-4-羥化酶(C4H)、查爾酮合成酶(CHS)、花青素合成酶(ANS)、花青素還原酶(ANR)和類黃酮O-甲基轉移酶(FOMT)基因的表達量較高,類黃酮 3′-羥化酶(F3′H)、類黃酮 3′,5′-羥化酶(F3′5′H) 和黃酮醇合成酶(FLS)基因的表達量則相對較低[14],而在黃秋葵花和果莢轉錄發現,3-黃烷酮羥化酶(F3H)、二氫黃酮醇-4-還原酶(DFR)在黃秋葵花中表現上調效應,類黃酮 3′,5′-羥化酶(F3′5′H)、花青素還原酶(ANR)、 無色花色素還原酶(LAR)在黃秋葵果莢中表現顯著上調效應,花青素合酶(ANS)、花青素3-O-葡糖基轉移酶(GT)則分別在花和果莢中均有上調或下調效應[15],與葡萄黃酮代謝相關的許多基因及轉錄因子在冬季果和夏季果中表達不一致,冬季果中上調表達[16]。【本研究切入點】關于黃酮類化合物轉錄組和代謝組的研究很多,但關于不同地面覆蓋方式下鮮食葡萄果皮黃酮轉錄組和代謝組的相關研究報道較少。需分析不同地面覆蓋方式下新郁葡萄果皮黃酮轉錄組和代謝物含量的差異。【擬解決的關鍵問題】以7年生新郁葡萄為試材,以清耕栽培為對照,分別進行行間鋪設園藝地布和行間生草栽培兩種地面覆蓋方式處理,從分子生物學和代謝組學水平研究不同地面覆蓋方式下為黃酮的合成調控機制,為新郁葡萄高效優質栽培提供新模式。
1 材料與方法
1.1 材 料
1.1.1 葡 萄
試驗于2021年在新疆維吾爾自治區葡萄瓜果研究所進行。以2015年種植生長勢一致的鮮食葡萄新郁為試材,東西行向,株行距1.5 m×5.0 m,順行龍干+(V+水平)葉幕管理,葉幕高度160 cm,果穗結果高度140 cm,單面葉幕長180 cm,新梢間距統一設置為15 cm,單穗果粒數為80粒左右,田間統一水肥管理及病蟲害綜合防控。
試驗共設有2種地面覆蓋方式:(1)行間鋪設園藝地布覆蓋(F),(2)生草栽培覆蓋(S),以清耕栽培(Clean tillage)為對照(Q)。2021年5月20日在新郁葡萄行間種植馬齒莧(S),待馬齒莧完全覆蓋地面時(2021年6月16日),在新郁葡萄行間鋪設黑色園藝地布(F),每處理約300 m2,3次重復。待果實達到該品種生理成熟時,從果穗的上、中、下三個部位(清耕Q-1、Q-2、Q-3,園藝地布F-1、F-2、F-3,生草栽培S-1、S-2、S-3)采集樣品約1 000 g,立即帶回實驗室,迅速剝取3 g果皮,每處理重復3次,共9份樣品。用錫紙包裹后置于液氮中,隨后將樣品置于-80℃冰箱內冷凍保存,用于轉錄組和代謝組測定。
1.1.2 試 劑
甲醇(色譜純),Merck;乙腈(色譜純),Merck;甲酸(色譜純),Sigma-Aldrich;標準品大于98%,MCE(70%甲醇配制,10 mmol/L)。
1.2 方 法
1.2.1 代謝物提取
稱取20 mg樣本粉末加入10 μL濃度為4 000 nmol/L的內標混合工作液和500 μL 70%的甲醇溶液,超聲提取30 min,4℃條件下12 000 r/min離心5 min,吸取上清液,用0.22 μm濾膜過濾樣品,并保存與進樣瓶中用于LC-MS/MS分析。試驗委托武漢邁特維爾生物科技有限公司完成。
1.2.2 LC-MS/MS進行代謝組分及數據處理
數據采集儀器系統包括超高效液相色譜(Ultra Performance Liquid Chromatography, UPLC)和串聯質譜(Tandem Mass Spectrometry,MS/MS)。
液相條件:(1)色譜柱:Waters ACQUITY UPLC HSS C18柱(1.8 μm,100 nm×2.1 mm i.d.);(2)流動性:A相為超純水(加入0.05%的甲酸),B相為乙腈(加入0.05%的甲酸);(3)流速0.35 mL/min;柱溫40℃;進樣量2 μL。(4)洗脫梯度:0 min A/B為90∶10(V/V), 1 min A/B為80∶20(V/V),9 min A/B為30∶70(V/V),12.5 min A/B為5∶95(V/V),13.5 min A/B為5∶95(V/V),13.6 min A/B為90∶10(V/V),15 min A/B為90∶10(V/V)。
色譜條件:點噴霧離子源(Electrospray Ionization,ESI)溫度550℃,正離子模式下質譜電壓5 500 V,負離子模式下質譜電壓-4 500 V,氣簾氣(Curtain Gas,CUR)35 psi。在Q-Trap6500+中,每個離子對時根據優化的去簇電壓(Declustering Potential,DP)和碰撞能(Collision Energy,CE)掃描檢測。
采用Analyst 16.3軟件處理質譜數據。橫坐標為檢測的保留時間(Time,min),縱坐標為離子檢測的離子流強度(Intensity,cps)。將檢測到的所有樣本的積分峰面積比值代入標準曲線線性方程進行計算,得到實際樣本中該物質含量。
樣本中黃酮的含量(nmol/g)
=C×V/1 000 000/m.
式中,C:樣本的積分峰面積比值代入標準曲線得到的樣本濃度值(nmol/L);V:提取時所用溶液的體積(μL);m:稱取的樣本質量(g)。
1.3 數據處理
1.3.1 轉錄組分析
RNA提取后用瓊脂糖凝膠電泳分析RNA的完整性及是否存在DNA污染,Qubit 2.0熒光儀檢測RNA濃度,Agilent 2100生物分析儀檢測RNA完整性后對文庫的insert size 進行檢測,insert size符合預期后進行下一步試驗。
樣品經文庫構建和文庫質檢合格后,用Illumina HiSep平臺測序,將測序結果與參考基因組(http://plants.ensembl.org/Vitis_vinifera/Info/Index)進行序列比對,進行功能注釋。采用EPKM(Fragments Per Kilobase of transcript per Million fragments mapped)統計分析。將|log2Fold Change|>=1,且FDR(False Discovery Rate)<0.05作為差異基因的篩選條件。
1.3.2 轉錄組和代謝組聯合分析
將代謝組和轉錄組數據結果利用GO注釋、WEGO軟件進行功能分類統計,對篩選出來的差異基因與差異代謝物進行聯合分析。
2 結果與分析
2.1 不同地面覆蓋方式下新郁葡萄果皮轉錄組測序
2.1.1 測序質量
研究表明,共獲得63.26 G測序數據,各樣品的總堿基數均達到6.26 G以上,堿基質量超過Q30的比例均高于91.57%,GC含量均高于44.16%,測序組裝精度高,符合分析要求,可用于后續分析。表1
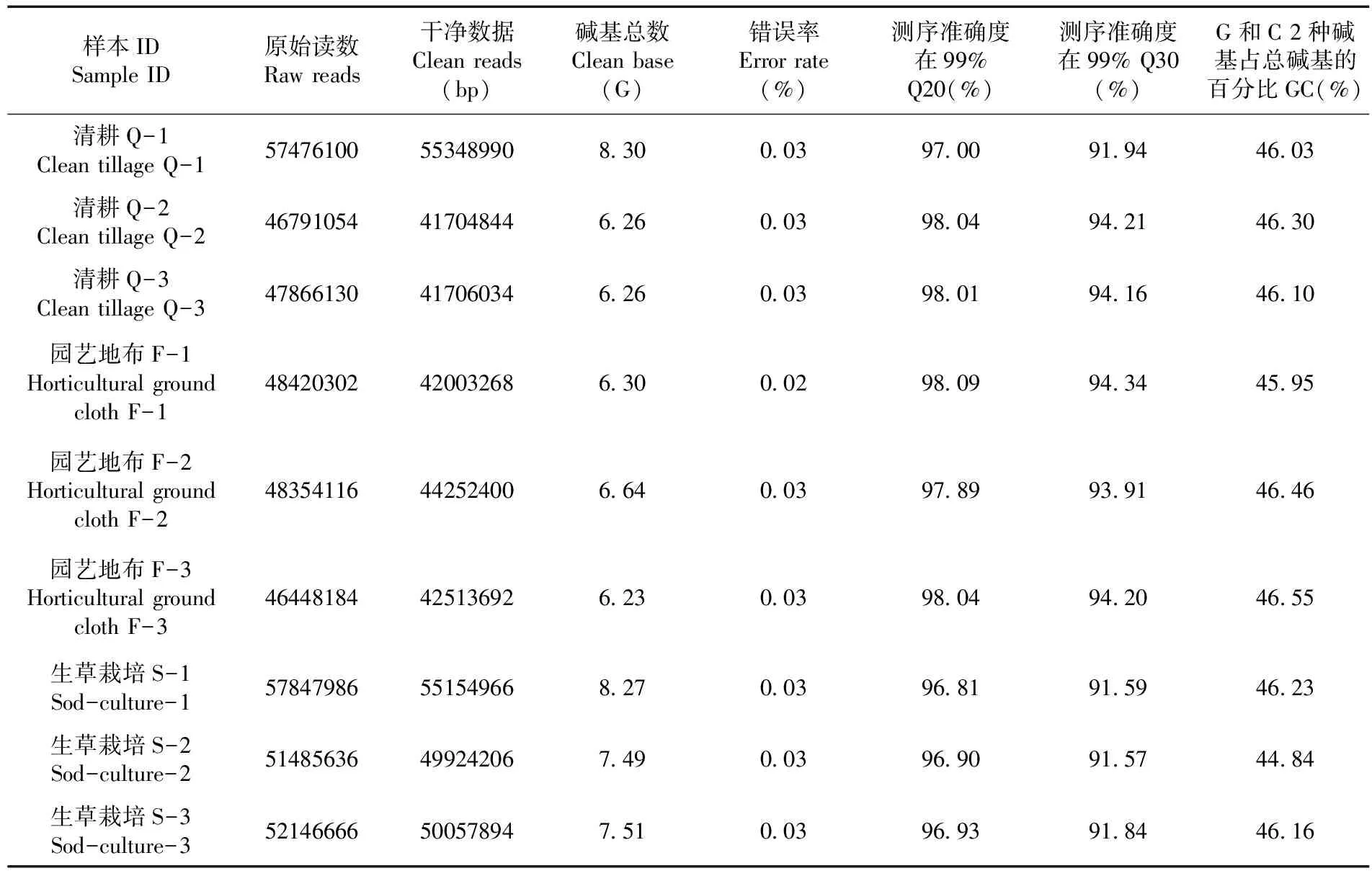
表1 不同地面覆蓋方式轉錄組測序質量變化
2.1.2 不同地面覆蓋方式下葡萄果皮黃酮差異基因
研究表明,新郁葡萄經園藝地布和生草栽培處理后,園藝地布覆蓋處理的葡萄果皮中共找到587個表達差異顯著的基因,其中上調基因317個、下調基因270個;生草栽培的葡萄果皮中共找到177個表達差異顯著的基因,其中上調基因56個、下調基因121個。表2
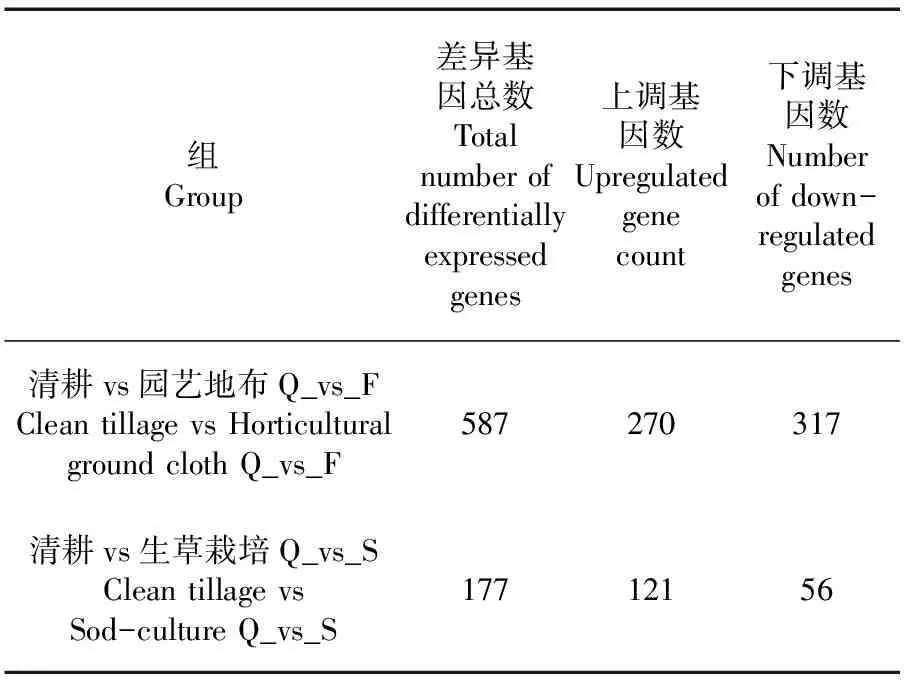
表2 差異基因數量
2.1.3 差異表達基因GO分類和功能富集
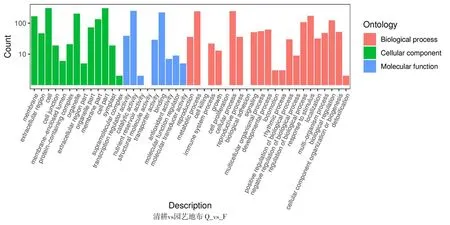
研究表明,16 969個基因得到了功能注釋,其中差異基因得到功能注釋最多的是對照與園藝地布,共364個。園藝地布注釋的基因涉及45個功能組,生草栽培注釋的基因組涉及37個功能組,均主要集中在細胞組分、分子功能和生物學過程等。圖1,表3
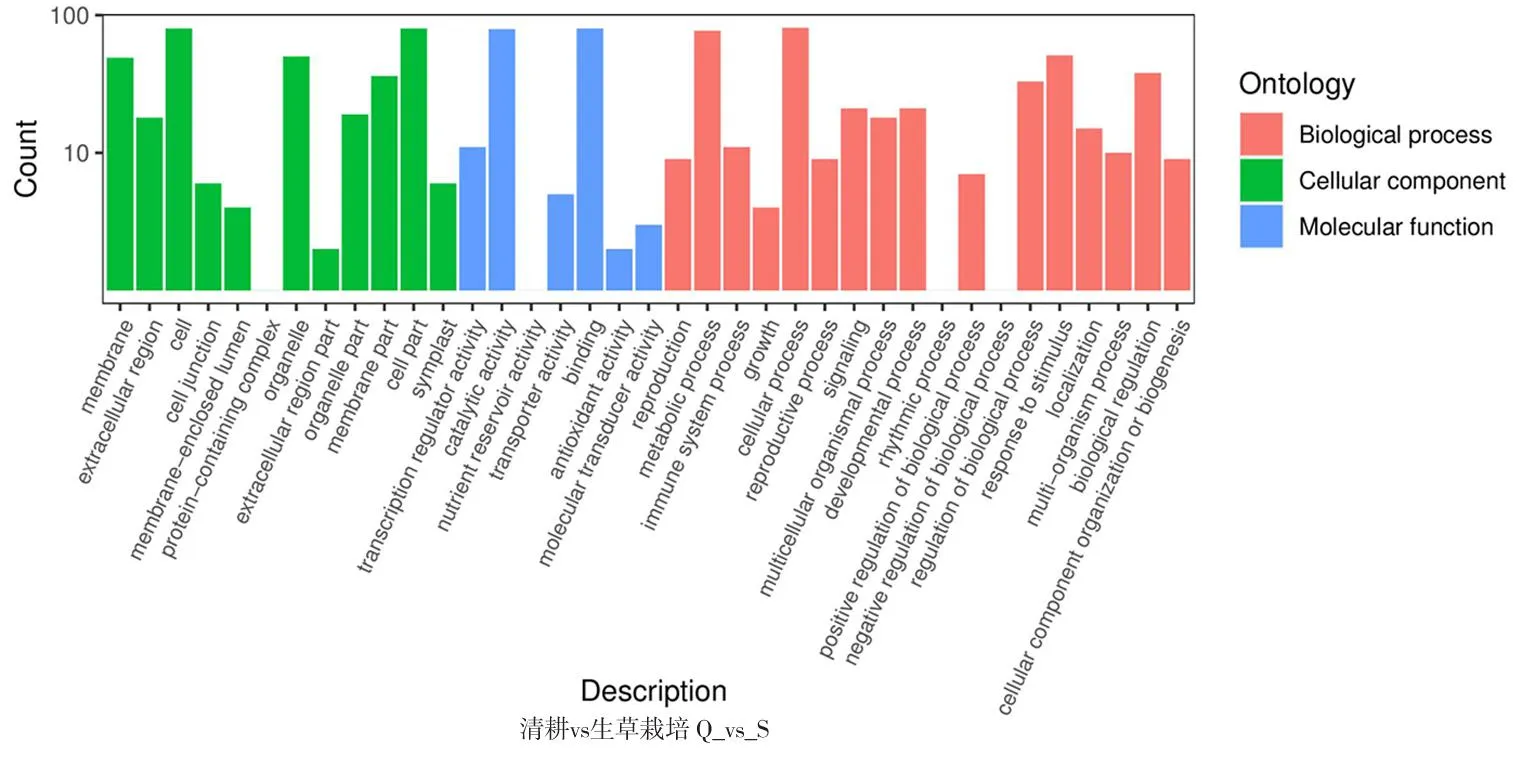
注(Note):生物學過程(Biological process);細胞組分(Cellular component);分子功能(Molecular function);細胞膜(Membrane);胞外區(Extracellular region);細胞(Cell);細胞連接(Cell junction);膜內腔(Membrane-enclosed lumen);蛋白復合體(Protein-containing complex);細胞器(Organelle);細胞外基質部分(Extracellular region part);細胞器組件(Organelle part);細胞膜組件(Membrane part);細胞組件(Cell part);共質體(Symplast);超分子復合物(Supramolecular complex);翻譯調節活性(Transcription regulator activity);催化活性(Catalytic activity);營養儲層活動(Nutrient reservoir activity);結構分子活性(Structural molecule activity);轉運器活動(Transporter activity);綁定(Binding);抗氧化活性(Antioxidant activity);分子功能調節(Moleular function regulator);分子傳感器活性(Molecular transducer activity);生殖(Reproduction);代謝過程(Metabolic process);生長(Growth);細胞增殖(Cell proliferation);細胞進程(Cellular process);生殖過程(Reproductive process);生物黏附(Biological adhesion);信號(Signaling);多細胞有機體進程(Multicellular organismal process);發育進程(Developmental process);運動力(Locomotion);節律進程(Rhythmic process);生物過程的正調控(Positive regulation of biological process);生物過程的負調控(Negative regulation of biological process);應激反應(Response to stimulus);定位(Localization);多有機體進程(Multi-organism process);生物調節(Biological regulation);細胞成分組織或生物合成(Cellular component organization or biogenesis);解毒(Detoxification);橫坐標表示二級GO條目,縱坐標表示GO條目的差異基因的數量(The abscissa represents the second-level GO entry,and the ordinate represents the number of differential genes in the GO entry)

表3 GO數據庫三大功能主類DEG數
2.1.4 差異表達基因KOG注釋
研究表明,分別將園藝地布和生草栽培注釋到KOG數據庫的175個和86個DEGs進行直系同源分類,各自獲得22個和16個功能分類。在園藝地布覆蓋方式下,R(一般功能預測)獲得46個注釋結果,占16.73%,Q(次級代謝物的生物合成、轉運和代謝),獲得38個注釋結果,占13.82%;在生草栽培模式下,Q(次級代謝物的生物合成、轉運和代謝)獲得21個注釋結果,占24.42%,R(一般功能預測)獲得17個注釋結果,占19.77%。圖2
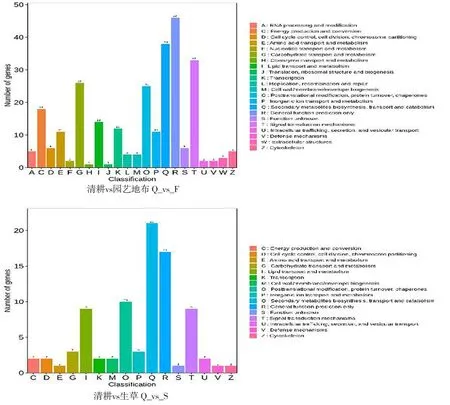
注:(Note)A,RNA的加工與修飾(RNA processing and modification);C,能源的產生與轉化(Energy production and conversion);D,細胞周期調控,細胞分裂,染色體分配(Cell cycle control,cell division,chromosome partitioning);E,(氨基酸轉運與代謝)Amino acid transport and metabolism;F,(核苷酸轉運與代謝)Nucleotide transport and metabolism;G,碳水化合物轉運與代謝(Carbohydrate transport and metabolism);H,輔酶轉運與代謝(Coenzyme transport and metabolism);I,脂質轉運與代謝(Lipid transport and metabolism);J,翻譯、核糖體結構和生物合成(Translation,ribosome structure and biosynthesis); K,轉錄(transcription);L,復制、重組和修復(Replication,recombination and repair);M,細胞壁/細胞膜的生物發生(Cell wall/membrane/envelope biogenesis);O,次生代謝物合成、轉運和代謝(Posttranslational modification,protein turnover,chaperones);P,無機離子轉運與代謝(Inorganic ion transport and metabolism);Q,次級代謝物的生物合成、轉運和代謝(Secondary metabolites biosynthesis,transport and cetabolism);R,一般功能預測(General function prediction only);S,未知功能(Function Unknown);T,信號轉導機制(Signal transduction mechanisms);U,胞質運輸、分泌和囊泡運動(intracellular trafficking,secretion and vesicle transport);V,防御機制(Defense mechanisms); W,細胞外結構(xtracellular structures);Z, 細胞骨架(cytoskeleton)
2.1.5 差異表達基因KEGG注釋與富集
研究表明,園藝地布覆蓋的193個差異基因被注釋到KEGG通路中,與黃酮物質生物合成的相關富集途徑有次級代謝物生物合成、苯丙醇生物合成、類黃酮生物合成、苯丙氨酸生物合成、植物晝夜節律、二苯乙烯類二芳庚類和姜辣素類。生草栽培的61個差異基因被注釋到KEGG通路中,與黃酮相關富集途徑有次級代謝物生物合成。圖3、圖4
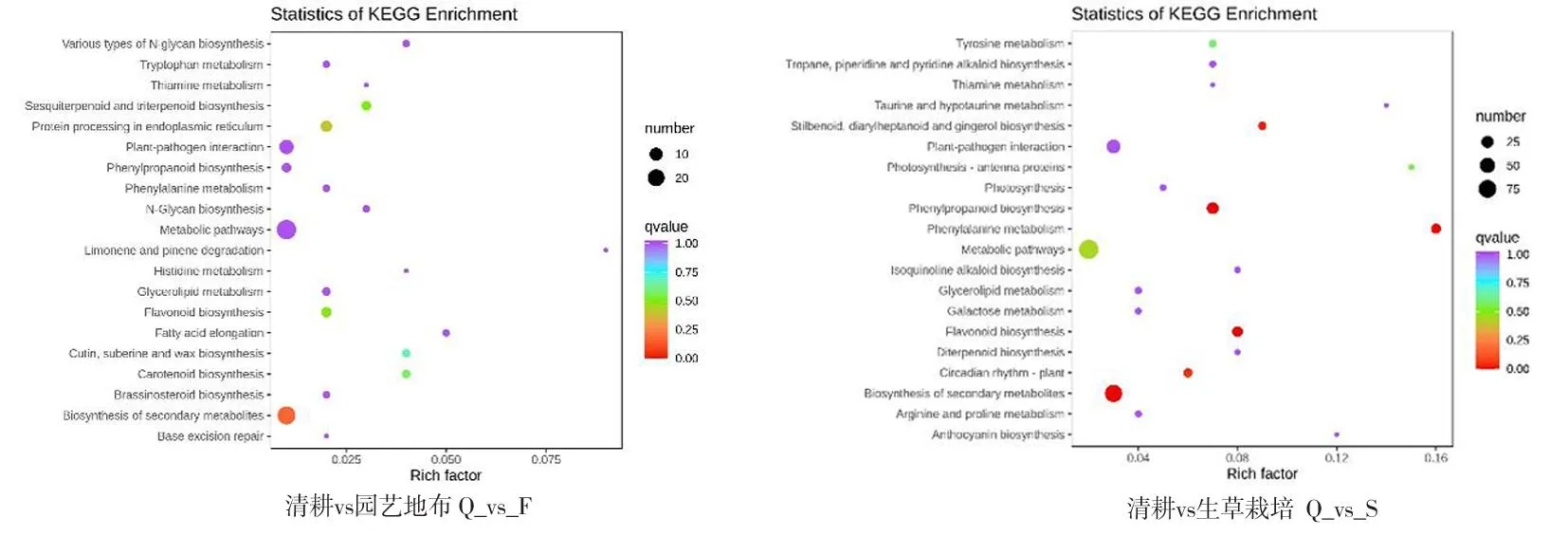
注(Note):縱坐標表示KEGG通路。橫坐標表示Rich factor。Rich factor越大,富集的程度越大。點越大,通路富集的差異基因的數量越多。點的顏色越紅,代表富集越顯著(The ordinate represents the KEGG pathway. Abscissa represents the Rich Factor. The larger the Rich factor,the greater the degree of enrichment. Larger point indicates more number of differential genes enriched by pathway. Color shows significance with most significant in red);酪氨酸代謝(Tyrosine metabolism);莨菪烷、哌啶和吡啶生物堿的生物合成(Tropane,piperidine and pyridine alkaloid biosynthesis);硫胺素新陳代謝(Thiamine metabolism);牛磺酸和次牛磺酸代謝(Taurine and hypotaurine metabolism);二芳基庚烷和姜醇的生物合成(Stilbenoid,diarylheptanoid and gingerol biosynthesis);植物-病原互作(Plant-pathogen interaction);光合作用-天線蛋白(Photosynthesis-antenna proteins);光合作用(Photosynthesis);苯丙烷生物合成途徑(Phenylpropanoid biosynthesis);苯丙烷類的代謝途徑(Phenylalanine metabolism);代謝途徑(Metabolic pathways);異黃酮生物合成途徑(Isoquinoline alkaloid biosynthesis);甘油酯代謝(Glycerolipid metabolism);半乳糖代謝(Galactose metabolism);類黃酮生物合成途徑(Flavonoid biosynthesis);二萜生物合成(Diterpenoid biosynthesis);晝夜節律-植物(Circadian rhythm-plant);次級代謝物生物合成(Biosynthesis of secondary metabolites);精氨酸和脯氨酸代謝(Arginine and proline metabolism);花青素生物合成(anthocyanin biosynthesis);各種類型的N -聚糖生物合成(Various types of N-glycan biosynthesis);色氨酸代謝(Tryptophan metabolism);酪氨酸代謝(Thiamine metabolism);類倍半萜烯和三萜(Sesquiterpenoid and triterpenoid);內質網中的蛋白質加工(Protein processing in endoplasmic reticulum);N-寡糖生物合成(N-Glycan biosynthesis);檸檬烯和蒎烯的降解(Limonene and pinene degradation);組氨酸代謝(Histidine metabolism);甘油酯代謝(Glycerolipid metabolism);脂肪酸延伸(Fatty acid elongation);角質、亞伯堿和蠟的生物合成(Cutin,suberine and wax biosynthesis);類胡蘿卜素合成(Carotenoid biosynthesis);油菜素類固醇生物合成(Brassinosteroid biosynthesis);堿基切除修復(Base excision repair)。
2.2 不同地面覆蓋方式下黃酮代謝組變化
2.2.1 黃酮代謝組
研究表明,園藝地布和生草栽培模式下的新郁葡萄果皮共檢測到60種代謝物,共分為12類,其中花青素1種,查爾酮類6種,黃烷醇類6種,二氫黃酮類4種,二氫黃酮醇類6種,黃酮類11種,黃酮醇類17種,異黃酮類3種,酚酸類3種,異芒果苷1種,茶黃素1種,黃酮碳糖苷1種。
黃烷醇類含量最高,對照、園藝地布和生草栽培的黃烷醇含量占總黃酮含量的比例高達52.41%~63.70%,其次是黃酮醇,占總黃酮含量的24.32%~29.40%,再次是二氫黃酮醇含量,占總黃酮含量的5.93%~8.03%,黃酮碳糖苷所占比例最低,幾乎可以忽略不計。生草栽培的黃烷醇類含量顯著高于對照(P<0.05),比對照提高了19.42%,園藝地布的黃烷醇類含量雖較對照有所增加,但無顯著差異;園藝地布的黃酮醇含量較對照有所提高,生草栽培降低了黃酮醇含量,但和對照間均無顯著差異;園藝地布的二氫黃酮醇含量高于對照,生草栽培的二氫黃酮醇含量低于對照,但和對照間無顯著差異性。
園藝地布覆蓋差異代謝物7種,下調2種,包括異黃酮類如毛蕊異黃酮(Flavonoid_96),其他1種(Flavonoid_154),對照的黃酮類如毛蕊異黃酮含量極低,幾乎為0.000 mg/kg,對照的含量為0.005 mg/kg,Flavonoid_154的含量比對照降低了99.70%;上調5種,包括黃酮醇類4種,如楊梅素類(Flavonoid_92、Flavonoid_178)、異鼠李蘇(Flavonoid_58)、西伯利亞落葉松黃酮(Flavonoid_188),查爾酮1種(Flavonoid_177)。園藝地布覆蓋栽培的楊梅素(Flavonoid_92)類是對照的3.10倍、楊梅(Flavonoid_178)是對照的2.67倍、異鼠李蘇含量是對照的2.25倍、Phloretin(Flavonoid_177)是對照的2.25倍。園藝地布覆蓋栽培的西伯利亞落葉松黃酮含量是0.058 mg/kg,而對照的含量幾乎為0.000 mg/kg。
生草栽培覆蓋差異代謝物11種,且全部下調,查爾酮1種如新橙皮甙二氫查爾酮(Flavonoid_22),二氫黃酮醇2種如二氫楊梅素(Flavonoid_40)和異水飛薊賓(Flavonoid_04),黃酮1種Nicotiflorin(Flavonoid_42),黃酮醇6種如楊梅素(Flavonoid_92)、紫云英苷(Flavonoid_119)、異鼠李素-3-O-新橙皮苷(Flavonoid_54)、Baimaside(Flavonoid_138)、Afzelin(Flavonoid_175)和堪非醇 3-新橙皮糖苷(Flavonoid_52),異黃酮1種,毛蕊異黃酮苷(Flavonoid_96)。新橙皮甙二氫查爾酮、二氫楊梅素、Nicotiflorin、楊梅素、紫云英苷、異鼠李素-3-O-新橙皮苷、Baimaside、Afzelin和堪非醇 3-新橙皮糖苷分別比對照增加了82.61%、51.21%、55.13%、62.275%、52.91%、57.82%、62.26%、68.38%和58.82%;生草栽培的異水飛薊賓和毛蕊異黃酮苷含量極低,而對照的含量分別為0.008和0.005 mg/kg。表4

續表 4 不同地面覆蓋方式下葡萄果皮黃酮代謝物組分和含量變化
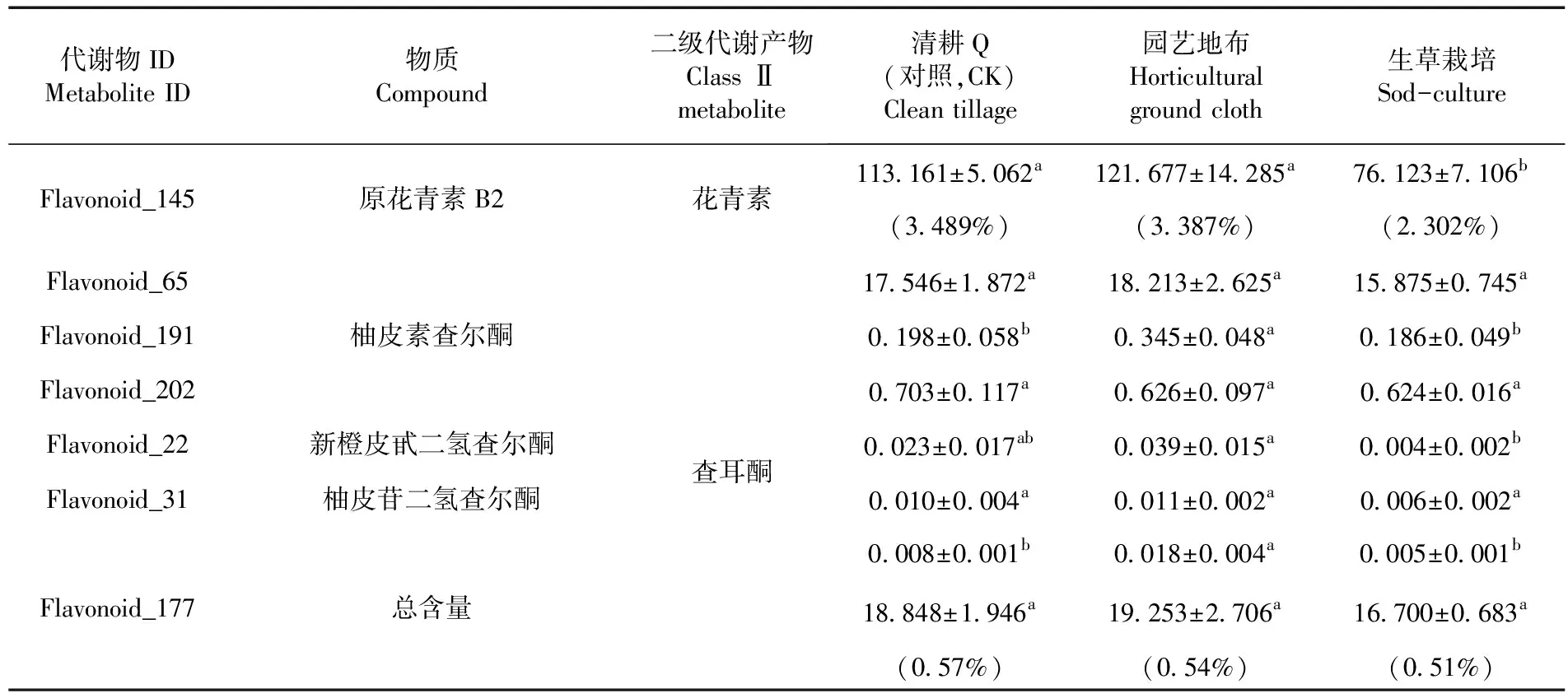
表4 不同地面覆蓋方式下葡萄果皮黃酮代謝物組分和含量變化
2.2.2 不同地面覆蓋方式下黃酮差異代謝物KEGG功能注釋及富集
研究表明,園藝地布覆蓋的8個代謝物被注釋到代謝過程,黃酮和黃酮醇生物合成是主要的,其次是類黃酮生物合成,再次是次級代謝產物的生物合成。生草覆蓋栽培的9個代謝物被注釋到代謝過程,黃酮和黃烷醇代謝物生物合成是主要的,其次是次生代謝產物的生物合成,再次是類黃酮的生物合成。2種地面覆蓋方式下與黃酮類物質合成最重要的途徑是黃酮和黃酮醇生物合成途徑。圖5
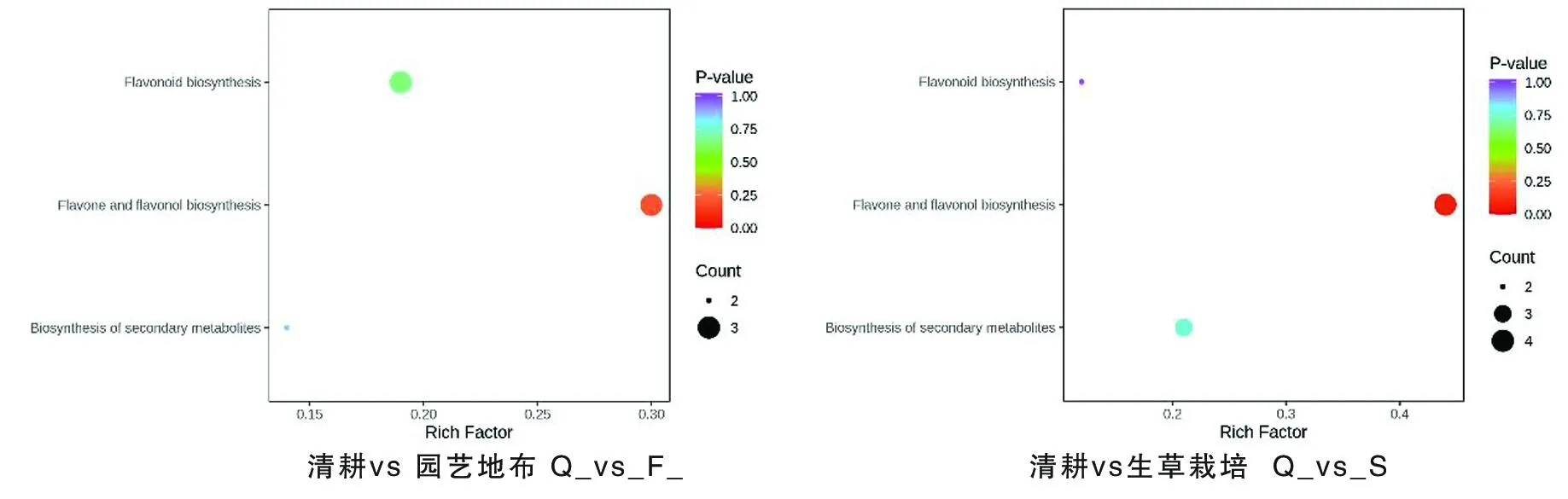
注(Note):縱坐標表示每個通路對應的Rich factor,縱坐標為通路名稱,點的顏色為P value,越紅表示富集越顯著。點的大小代表富集到的差異代謝物的個數多少(The ordinate represents the Rich factor corresponding to each pathway,the ordinate represents the pathway name,and the color of the point is P value. The redder indicates the more significant enrichment. The size of the dot represents the number of enriched differential metabolites);類黃酮生物合成(Flavonoid biosynthesis); 黃酮和黃烷醇代謝物生物合成(Flavone and flavonol biosynthesis); 次級代謝物生物合成(Biosynthesis of secondary metabolites)
Q-F組的5種差異代謝物中,楊梅素(Flavonoid_92)、楊梅素(Flavonoid_178)、查爾酮(Flavonoid_177)和異鼠李素(Flavonoid_58)地布高于對照,差異倍數為1.16~1.63,黃酮碳糖苷(Flavonoid_154)含量地布低于對照,差異倍數為1.47。Q-S組的9種差異代謝物中,二氫楊梅素(Flavonoid_40)、紫云英苷(Flavonoid_119)、黃酮(Flavonoid_42)、異鼠李素-3-O-新橙皮苷(Flavonoid_54)、堪非醇3-新橙皮糖苷(Flavonoid_52)、黃酮醇(Flavonoid_138)、楊梅素(Flavonoid_92)、黃酮醇(Flavonoid_175)和新橙皮甙二氫查爾酮(Flavonoid_22)含量生草低于對照,差異倍數為1.04~2.68。圖6
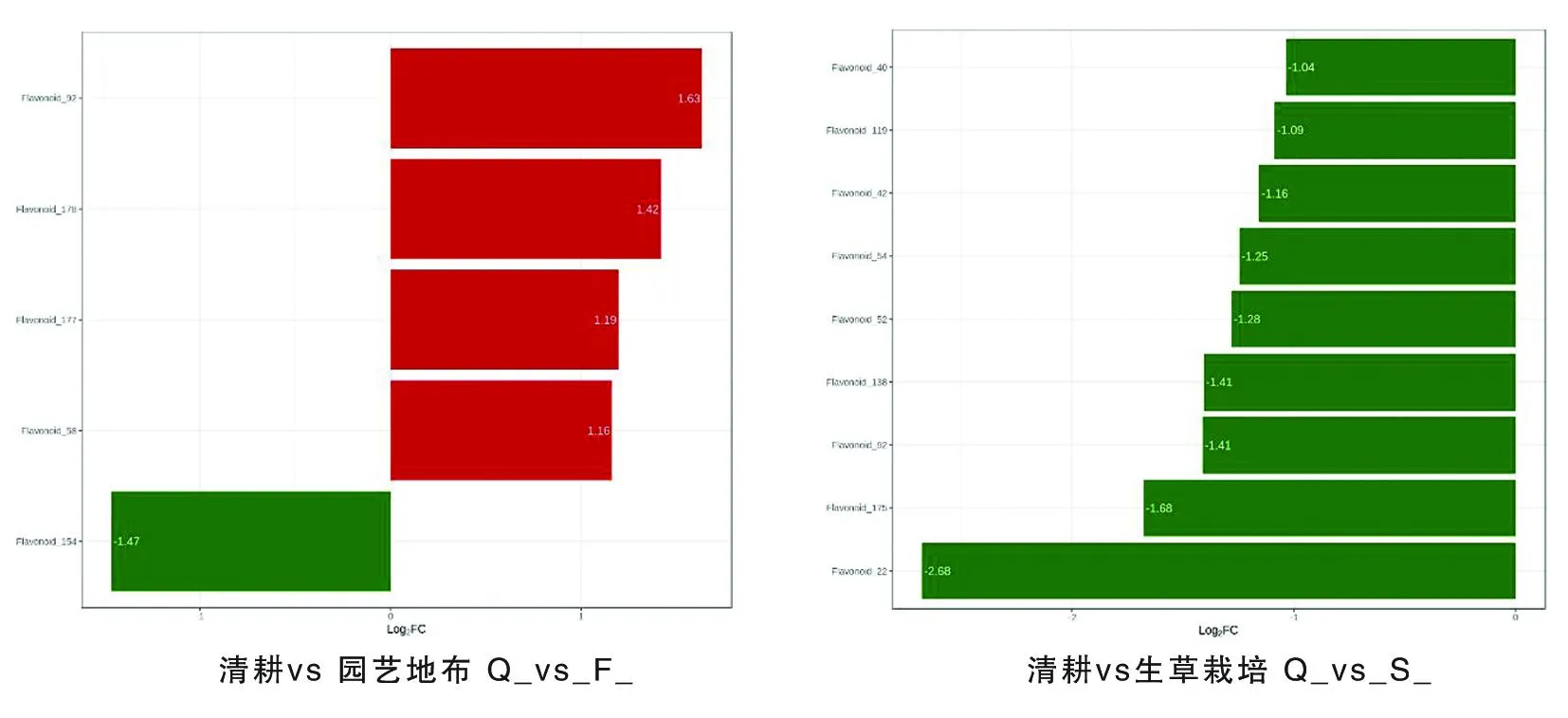
圖6 不同地面覆蓋方式下葡萄果皮差異代謝物差異倍數
2.3 不同地面覆蓋方式下黃酮定量轉錄組和代謝組聯合
研究表明,與黃酮合成相關富集途徑主要集中在黃酮和黃酮醇、類黃酮生物合成。參與園藝地布覆蓋類黃酮合成的基因有1個C4H、6個CHS、6個F3H、3個LAR、4個HCT、4個FLS和2個C12RT1,參與黃酮和黃酮醇生物合成的有2個C12RT1;參與生草覆蓋類黃酮生物合成有6個CHS、6個F3H、8個F3′5′H、3個FLS和3個LAR,參與黃酮和黃酮醇生物合成的有3個F3′5′H。表5,圖7
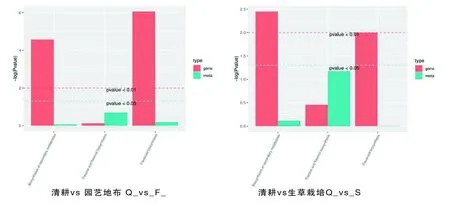
注(Note):橫坐標代表代謝通路,縱坐標中紅色代表差異基因的富集P value值,綠色代表差異代謝物的富集P value值,用log(P-value)表示,縱坐標越高,代表富集程度越強(The abscissa represents metabolic pathway,the red in the ordinate represents the enriched P value of differential genes,and the green represents the enriched P value of differential metabolites,represented by log(P-value). The higher the ordinate is,the stronger the enrichment degree is);次級代謝物生物合成(Biosynthesis of secondary metabolites);黃酮和黃烷醇代謝物生物合成(Flavone and flavonol biosynthesis);類黃酮生物合成(Flavonoid biosynthesis)
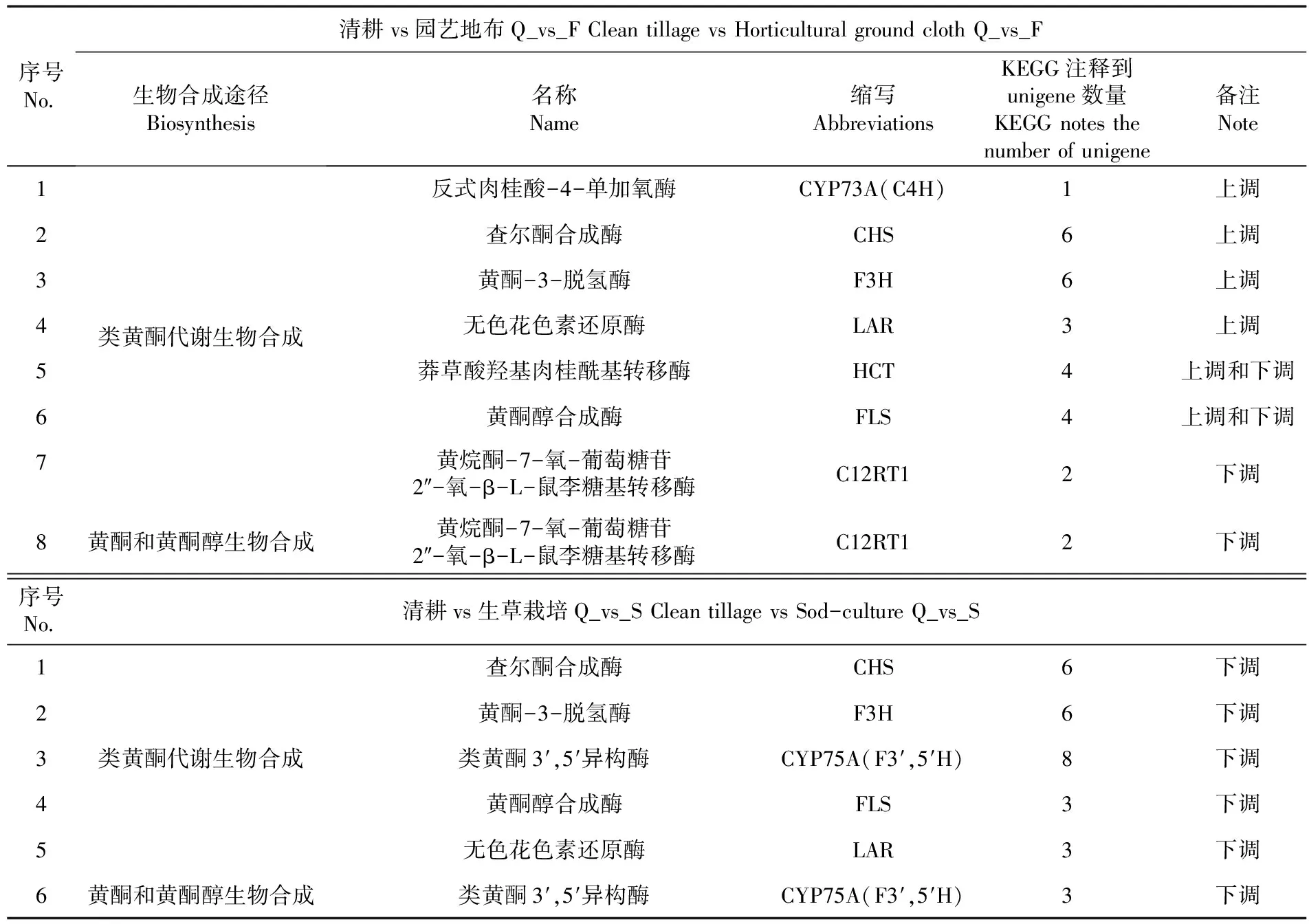
表5 新郁葡萄果皮黃酮生物合成途徑中候選基因
3 討 論
3.1黃酮是葡萄中含量最為豐富的一類次生代謝物,其在細胞質中合成,然后儲存在液泡中[17],這些化合物不僅能保護葡萄免受紫外線、病蟲害等傷害,還對葡萄的風味品質以及營養價值起著決定性作用[18]。
目前已報道的黃酮種類達9 000種[19-20],根據吡喃環的飽和取代狀態,可以分為花青素、黃酮、黃烷醇、黃烷酮、黃酮醇、查爾酮、原花青素和異黃酮等[21]。目前已報道的釀酒葡萄黃酮種類有40種[22]。研究利用UPLC-MS/MS方法測定不同地面覆蓋方式下的黃酮組分,均得到12大類60種黃酮物質,較前人[22]研究相比,極大地豐富了葡萄黃酮組分。在12類黃酮物質中,黃烷醇和黃酮醇所占比例較高,構成了黃酮物質的主要組成部分,與BAYDAR等[23]的研究結果相同。黃烷醇類的比例大于50%,生草栽培的高達63.73%,對照的占54.41%,園藝地布所占比例較低(52.83%),第二大比例的是黃酮醇,對照占總黃酮的比例最高為29.40%,園藝地布的為28.84%,生草栽培的較低為24.32%。在前期的研究結果中發現,園藝地布和生草栽培的花色苷含量顯著高于對照,生草栽培可以提高果實外觀和內在品質[24]。經進一步分析發現地布覆蓋的差異代謝物共有7種(下調2種,上調5種),生草覆蓋差異代謝物12種(全部下調),計算發現園藝地布覆蓋的總黃酮含量(3 592.749 mg/kg)高于對照的總黃酮含量(3 243.242 mg/kg),而Wang等[25]研究卻發現地布覆蓋降低黃酮含量。黃酮種類和含量除受遺傳因素影響外,還與光照、溫濕度和栽培管理措施密切相關,這些因素相互作用,共同調節果實的黃酮代謝[26]。
關于黃酮組分及其合成酶基因一直是近年來研究的熱點,也是目前研究較為清楚的次生代謝途徑之一[27]。該通路上主要存在2個基因群:上游基因群(early biosynthesis genes,EBGs)與下游基因群(later biosynthesis genes,LBGs)[28-29]。上游基因群主要包括查爾酮合成酶(Chalcone synthase,CHS)、查爾酮異構酶(Chalcone isomaerase,CHI)、黃酮-3-脫氫酶(Naringenin 3-dioxygenase,F3H)、類黃酮3′,5′異構酶(Flavonoid 3′,5′-hydroxylase,F3′5′H)等基因[30-31],下游基因群包括二氫黃酮醇4-還原酶(Dihydroflavonol 4-ruductase,DFR)、黃酮醇合成酶(Flavonol synthase,FLS)、花青素苷合成酶(Anthocyanidin,ANS)、花青素還原酶(Anthocyanin reductase,ANR)、糖基轉移酶(Glucosyltransferase,GT)、酰基轉移酶(Acyl transferase,AT)和甲基轉移酶(Methyl transferase,MT)等[32]。
3.2試驗通過Illumina HiSep高通量測序技術,對2種地面覆蓋方式下的葡萄果皮進行RNA-Seq轉錄組測序并進行組裝,共獲得63.26G測序數據,各樣品的總堿基數6.26 G~8.30 G,堿基質量超過Q30的比例91.57%~94.20%,GC含量44.16%~46.55%,該試驗測序組裝精度高,保障試驗的準確性。園藝地布覆蓋下新郁葡萄果皮中有587個表達差異顯著的基因,其中上調基因317個、下調基因270個,生草覆蓋下果皮中共有177個表達差異顯著的基因,其中上調基因56個、下調基因121個。并通過KEGG通路分析,園藝地布覆蓋中與黃酮相關富集途徑有次級代謝物生物合成、苯丙醇生物合成、類黃酮生物合成、苯丙氨酸生物合成、植物晝夜節律、二苯乙烯類二芳庚類和姜辣素類的生物合成,生草栽培中與黃酮相關富集途徑有次級代謝物生物合成。造成園藝地布覆蓋和生草栽培覆蓋下差異基因和黃酮代謝途徑不同的原因可能是由于地面覆蓋方式導致的。
轉錄組和代謝組聯合分析發現,參與園藝地布覆蓋類黃酮合成的候選基因有C4H、CHS、F3H、LAR、HCT、FLS和C12RT1,其中C4H、CHS、F3H、LAR上調,HCT和FLS即有上調也有下調,C12RT1下調,參與園藝地布黃酮和黃酮醇生物合成的候選基因C12RT1下調。參與生草覆蓋類黃酮生物合成的候選基因有CHS、F3H、F3′5′H、FLS和LAR,參與生草黃酮和黃酮醇生物合成的候選基因有F3′5′H,均表現為下調。黃酮物質的合成是一個復雜的生理生化過程,與環境如生物(細菌、真菌、昆蟲等)和非生物脅迫(低溫、高溫、鹽害、干旱、溫度、重金屬等)等息息相關[33]。但研究中,新郁葡萄果皮代謝通路中仍有部分基因未被發現,如查爾酮異構酶、花青素還原酶(Anthocyanin reductase,ANR)和黃酮合成酶(Flavone synthase,FNS),可能是這些基因未在成熟果皮中表達所致。此外造成同一類候選基因在不同地面覆蓋方式中表現為上調或下調的原因可能是地面覆蓋方式不同造成果穗微環境的差異造成的,Gaiotti等[34]、Mori等[35]也發現,CHS、F3H、F3′5′H的表達與溫度有關系。
下一步將開展不同地面覆蓋方式對葡萄果皮基因表達量方面的研究,對該研究結果進行驗證。
4 結 論
4.1獲得了新郁葡萄果皮的轉錄組數據,并通過轉錄組數據的生物信息分析和代謝物組分分析,對新郁葡萄果皮的代謝通路有了初步認識:園藝地布覆蓋與黃酮合成相關富集途徑主要集中在類黃酮生物合成,生草覆蓋栽培與黃酮合成相關富集途徑主要集中在次級代謝物和類黃酮生物合成,園藝地布覆蓋和生草覆蓋栽培的總黃酮含量高于對照。篩選出參與園藝地布覆蓋黃酮合成的候選基因有7個,分別是C4H、CHS、F3H、LAR、HCT、FLS、C12RT1;參與生草覆蓋栽培黃酮生物合成的候選基因有5個分別是是CHS、F3H、F3′5′H、FLS和LAR。
4.2對照園藝地布和生草栽培共檢測到12大類、60類黃酮物質,黃烷醇占總黃酮的比例最高,其中生草栽培黃烷醇類占總黃酮的比例為63.73%,園藝地布的比例最低為52.83%,對照為54.41%;其次是黃酮醇的比例,生草栽培占總黃酮的24.32%,園藝地布占28.84%,對照占29.40%。園藝地布的黃酮物質含量最高3 592.749 mg/kg,其次是生草栽培為3 306.859 mg/kg,對照的總黃酮物質含量最低為3 234.242 mg/kg。在吐魯番極端干旱區鮮食葡萄新郁可選擇生草栽培。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
四川蠶業(2021年3期)2021-02-12 02:38:46
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
中成藥(2018年11期)2018-11-24 02:57:00
中成藥(2017年8期)2017-11-22 03:19:40
中成藥(2017年10期)2017-11-16 00:50:13
