滇產單子葉植物的多樣性格局與保護
2024-03-19 08:14:16文妍莉
植物研究 2024年2期
文妍莉 李 嶸
(1.中國科學院昆明植物研究所,東亞植物多樣性與生物地理學重點實驗室,昆明 650201; 2.中國科學院大學,生命科學學院,北京100049)
生物多樣性是指地球上所有生命體與其生存環境相互作用形成的生態復合體[1],它是人類賴以生存和發展的基礎。全球生物多樣性的分布極不均衡,熱帶和山地的生物多樣性顯著高于其他區域[2-3]。然而,由于人類活動、氣候變化和資源過度利用等的影響,生物多樣性正急劇減少[4],因此,揭示生物多樣性的分布格局及其形成原因是阻止和逆轉生物多樣性喪失的重要途徑之一。
物種豐富度是反映生物多樣性豐富程度的重要參考指標,能準確揭示各地區的生物復雜性和多樣性[5],例如,Liu 等[6]利用物種豐富度揭示了秦嶺—大巴山南北坡的生物多樣性聚集度。近年來,高通量DNA 測序技術的發展和區域生命之樹的建立,為從系統發育的角度評估生物多樣性的組成提供了方法和途徑。系統發育多樣性是最早用來標識生物多樣性組成的進化指標[7],它用區域內所有物種在系統發育樹上的最短枝長之和來度量。Zhou 等[8]基于樟科(Lauraceae)植物的系統發育關系,并結合其地理分布,探討了中國樟科植物系統發育多樣性的分布格局,揭示了植被過渡區具有較高的進化組成多樣性。這類方法改變了以往將物種視為獨立個體的做法,強調物種間的演化關系,因而更好地反映生物多樣性在進化維度上的多樣性[9-12]。
特有現象是生物多樣性在進化過程中對不同區域自然環境適應和演化的結果[13],對于理解生物多樣性的形成與保護具有重要意義。Huang等[14]根據中國特有種子植物的地理分布,并結合物種間的系統發育關系,識別出我國生物多樣性保護的19個熱點和空白區。加權特有性和系統發育特有性是常用的量化分類學和進化歷史特有現象的指標[15-16]。
近年來,區域物種的受威脅狀態在生物多樣性保護領域受到越來越多的關注[17]。Hills等[18]利用進化特異性全球瀕危度指數對南非薯蕷屬(Dioscorea)植物多樣性進行分析,識別了南非重要的生物多樣性保護優先區。進化特異性全球瀕危度指數是將物種進化特異性與物種瀕危等級相結合[19],用以量化生物多樣性的區域優先保護級別[20]。
隨著全球數據的開放共享,綜合生物多樣性、特有性及瀕危度,從多個視角理解生物多樣性的形成與演變,可為生物多樣性的保護提供多維證據。如Zhang 等[21]對中國西南地區的維管植物多樣性進行研究發現,植物的多樣性、特有性和瀕危度的地理分布格局之間的重合度較低,這使得植物多樣性的綜合保護變得尤為困難。為更好地理解生物多樣性的分布格局和形成原因,有必要從不同類群、不同維度進行綜合研究,以便更精準地制定有效的生物多樣性保護策略。
云南地跨喜馬拉雅、印度—緬甸、中國西南山地3個全球生物多樣性熱點地區[22],富含豐富的珍稀瀕危和特有生物物種[14,23-26]。植物譜系組成多樣,包含木蘭分支、單子葉植物、真雙子葉植物等多個被子植物演化支系[27]。其中,單子葉植物是云南植物區系的重要組成部分,約占云南被子植物的15%[28]。因生活史短、進化速率高,其演化歷史有別于其他被子植物分支[29],也因其適應性強,分布極為廣泛,多是林下或林中樹上的伴生植物,或是高山、亞高山草甸的主要建群植物[30]。本研究以云南單子葉植物為例,基于物種的地理分布及其系統發育關系,從分類學和系統發育的角度探討單子葉植物的多樣性、特有性及受威脅程度的分布格局,并整合云南自然保護地的分布,辨識單子葉植物的保護空缺,為云南生物多樣性保護提供科學指導。
1 材料與方法
1.1 研究地區
云 南 地 處 中 國 西 南,位 于21°8′32″~29°15′8″N,97°31′39″~106°11′47″E。由于新近紀以來的造山運動,使得境內河谷深切、地勢陡峭,整體自東南向西北抬升[31]。由于受太平洋季風和印度洋季風的雙重影響,云南氣候表現出年溫差小、日溫差大、雨量充沛、干濕季分明等特點[32]。古南大陸成分、古北大陸成分、古地中海成分及東亞成分是云南植物區系的主體[33]。植被類型多樣,包括熱帶雨林、亞熱帶常綠闊葉林、針闊葉混交林、高山和亞高山灌叢和草甸等多種類型[30]。
1.2 數據來源
根據《云南植物志》[28]《云南省生物物種名錄(2016版)》[34]等資料,對云南單子葉植物物種數據庫進行補充完善,該數據庫包括植物學名、中文名、科屬位置、地理分布等信息。利用Flora of China(http://www.efloras.org/,FOC)、Plants of the World Online(https://powo.science.kew.org/,POWO)、World Flora Online(https://www.worldfloraonline.org/,WFO)、中國數字植物標本館(https://www.cvh.ac.cn/,CVH)、中國國家標本資源平臺(http://www.nsii.org.cn/,NSII)等線上數據庫對植物學名、地理分布信息進行修訂。科屬范疇以被子植物分類系統為依據[27],物種學名以Flora of China 為標準,地理分布以行政縣為單元。將亞種、變種等種下等級歸并到相應的種內,同時排除栽培、歸化和外來入侵物種。此外,根據《IUCN 物種紅色名錄的瀕危等級和標準:3.1 版》[35],結合《中國高等植物受威脅物種名錄》[36],將云南單子葉植物的受威脅狀態歸為易危、瀕危和極危。
為減少縣域面積大小對生物多樣性度量的影響,使用地理信息系統ArcGIS 10.2 將云南劃分為4 374個10 km×10 km 的等面積柵格。以云南省地圖做底圖,將植物縣域分布記錄根據經緯度匹配到相應的等面積柵格中,最終獲得929 114 條物種柵格分布記錄。底圖矢量數據來源于國家基礎地理信息中心(http://www.ngcc.cn/ngcc/)1∶100 萬矢量地圖數據,地理坐標參考系統為WGS_1984_UTM_Zone_47N。
1.3 系統發育樹構建
基于GBOTB 系統發育樹[37],利用R 3.6.2 中的V.PhyloMaker 程序包[38],根據云南單子葉植物物種數據庫構建滇產單子葉植物系統發育樹。參考V.PhyloMaker 中的Scenario 3 方法,對于骨架樹中缺失的物種,根據其所在科或屬的系統位置進行添加[38]。
1.4 多樣性度量指標
采用物種豐富度、系統發育多樣性、標準化系統發育多樣性、加權特有性、系統發育特有性度量云南單子葉植物多樣性。
物種豐富度(RS)為每個柵格內所有物種之和,計算公式如下:
式中:K表示柵格單元中的物種總數量。
系統發育多樣性(DP)是指連接系統發育樹根部到支端的枝長之和[7],計算公式如下:
式中:C是一組分支,表示一個柵格單元中所有類群到系統發育樹根部的最短生成路徑;c是一個分類單元的分支;Lc是分支c所代表的分支長度。
研究表明,物種豐富度和系統發育多樣性顯著相關[39],為消除物種豐富度對系統發育多樣性的影響,采用標準化的方法量化每個柵格的系統發育組成,也即標準化系統發育多樣性(DP,s)[40],計算公式如下:
式中:DP,o表示每個柵格內系統發育多樣性的觀測值,DP,r表示隨機化每個柵格的系統發育多樣性,σDP,r表示隨機化后的每個柵格的系統發育多樣性標準差。若DP,s為正,表明柵格系統發育組成多樣性相對較高;反之,柵格系統發育組成多樣性相對較低[41]。
加權特有性(EW)是指限定在特定范圍的物種數量[15],根據物種的分布范圍,為每個物種設置權重,分布范圍越小,權重越高,計算公式如下:
式中:n表示柵格的物種數量;Wi表示i物種在該柵格內的權重。
系統發育特有性(EP)是柵格內每個分支分布范圍的倒數加權的分支長度之和[16],計算公式如下:
式中:C是分類群到系統發育樹根部的最短路徑的分支集合;Lc是分支c的長度;Rc是分支c所有后代物種的全局范圍大小,其范圍大小用所占據的柵格單元總數來衡量,而不重復計算柵格。
使用受威脅指數(IT)和進化特異性全球瀕危度指數(IEDGE)度量云南不同柵格單子葉植物的受威脅程度。
受威脅指數(IT)是每個柵格內受威脅物種數量占柵格內全部物種的比例,計算公式如下:
式中:RT為柵格內受威脅物種數量,即極危、瀕危和易危物種的總和,RS為柵格內物種總數。
進化特異性全球瀕危度指數是將各柵格內每一物種的進化特異性(ED)與全球受威脅等級(GE)進行綜合分析得到的指標[19],計算公式如下:
式中:ED是基于GBOTB 骨架樹構建的云南單子葉植物系統發育樹計算獲得的進化特異性值;GE是受威脅物種的分值(易危1分、瀕危2分、極危3 分)。最后,根據柵格內每個物種的IEDGE,計算各柵格所有物種IEDGE的平均值作為各柵格單子葉植物的受威脅程度。
物種豐富度與系統發育多樣性(標準化系統發育多樣性)、加權特有性與系統發育特有性、受威脅指數與進化特異性全球瀕危度指數的相關性分析采用Pearson 指標。上述所有分析利用R 3.6.2 軟件中的函數cor()、picante 程序包[42]和Biodiverse 3.2軟件完成。
1.5 保護空缺分析
將云南單子葉植物的多樣性、特有性及受威脅程度的前30%和5%定義為熱點區域,表示云南單子葉植物的重點保護區域。使用ArcGIS 10.2軟件,在同一圖層文件夾下依次打開物種豐富度熱點區域、系統發育多樣性熱點區域、物種特有性熱點區域、系統發育特有性熱點區域、受威脅物種熱點區域、進化特異性及全球受威脅度熱點區域和云南自然保護地圖層,熱點區域圖層中未被自然保護地所覆蓋的區域即為云南單子葉植物的保護空缺。
2 結果與分析
2.1 云南單子葉植物的多樣性格局
據統計,云南有單子葉植物35 科459 屬2 267種及38 亞種和101 變種。其中,局限分布于云南的特有種有317種,受威脅物種436種(極危52種、瀕危166種、易危218種)。
云南單子葉植物的物種豐富度與系統發育多樣性顯著正相關(R=0.99,P<0.001;見圖1A),與標準化系統發育多樣性顯著負相關(R=-0.42,P<0.001;見圖1B);云南單子葉植物的加權特有性與系統發育特有性顯著正相關(R=0.98,P<0.001;見圖1C);云南單子葉植物的受威脅指數與進化特異性全球瀕危度指數顯著正相關(R=0.64,P<0.001;見圖1D)。云南西北部、南部和東南部的物種組成和系統發育組成多樣性和特有性均較高(見圖2A~D);然而,當消除物種豐富度對系統發育多樣性的影響后,除西北部和東南部的標準化系統發育多樣性較低外,其他區域的標準化系統發育多樣性均較高(見圖3)。
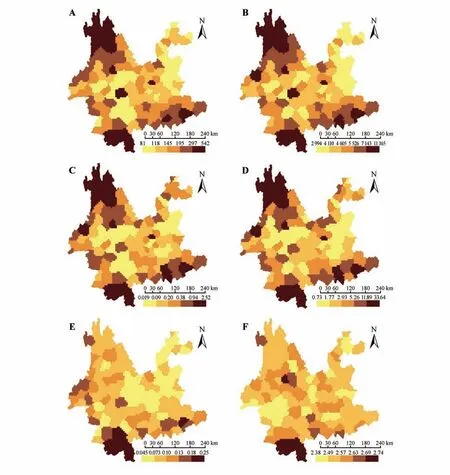
圖2 云南單子葉植物的多樣性格局A.物種豐富度;B.系統發育多樣性;C.加權特有性;D.系統發育特有性;E.受威脅指數;F.進化特異性全球瀕危度指數。Fig.2 Diversity patterns of Yunnan monocotyledons A.Species richness;B.Phylogenetic diversity;C.Weighted endemism;D.Phylogenetic endemism;E.Threatened index;F.Evolutionary distinctiveness and globally endangered index.

圖3 云南單子葉植物的標準化系統發育多樣性Fig.3 Standardized phylogenetic diversity patterns of Yunnan monocotyledons
2.2 區域受威脅狀況與重點保護區域
Pearson相關性分析表明受威脅指數與進化特異性全球瀕危度指數呈顯著正相關(R=0.64,P<0.001),且二者的分布格局均顯示云南西北部和南部是單子葉植物受威脅程度最為嚴重的區域(見圖2E~F)。云南單子葉植物的多樣性、特有性、受威脅程度與自然保護地的圖層疊加顯示,云南西北部、南部和東南部是單子葉植物的重點保護區域(見圖4)。
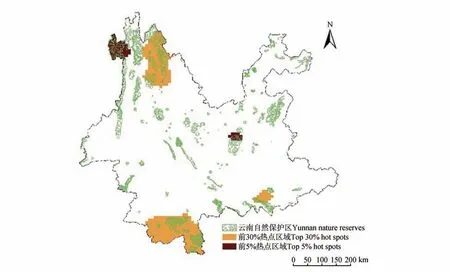
圖4 云南單子葉植物的重點保護區域Fig.4 Key protected areas of Yunnan monocotyledons
3 討論
3.1 云南單子葉植物多樣性的演化歷史
研究表明云南單子葉植物的物種豐富度與系統發育多樣性、加權特有性與系統發育特有性均呈顯著正相關(見圖1A,C),云南的西北部、南部和東南部是植物多樣性和特有現象最為豐富的地區(見圖2A~D),這與前期不同類群的研究結果一致,如云南被子植物[43]、云南被子植物薔薇分支[25]、云南被子植物菊類分支[26]。然而,使用零模型去除物種豐富度對系統發育多樣性的影響后,發現云南單子葉植物的譜系多樣性在西北部和東南部較低,而在其他區域較高(見圖3)。新生代以來,地質歷史事件如印度板塊與歐亞板塊的碰撞,對云南西北部地理環境的塑造有著深遠的影響,加之中新世以來印度洋西南季風的增強,更是為植物提供了多樣的生存環境,成為百合屬(Lilium)、鹿藥屬(Maianthemum)、豹子花屬(Nomocharis)、貝母屬(Fritillaria)等類群的分化中心[44],而這些類群大都來源于相近的譜系分支如百合科(Liliaceae),導致該區域單子葉植物的物種多樣性豐富,而系統發育多樣性卻較低。云南南部和東南部地理位置特殊,地質環境和氣候條件比較穩定,且未受到第四紀冰期—間冰期的影響[45],是諸多喜熱喜濕類群,如豆蔻屬(Amomum)、石斛屬(Dendrobium)、蜘蛛報蛋屬(Aspidistra)、芭蕉屬(Musa)、萬代蘭屬(Vanda)、水玉簪屬(Burmannia)等的重要聚集地[46],因此,這些地區的物種組成較為多樣。然而,云南東南部主要為喀斯特地貌,由于環境的過濾作用和植物自身的系統發育保守性[47],組成該區域的植物主要來自相同的進化支系,如天南星科(Araceae)的芋頭屬(Colocasia)、崖角藤屬(Rhaphidophora)、石柑子屬(Pothos)、犁頭尖屬(Typhonium)等,導致云南東南部單子葉植物的物種組成多樣,但系統發育組成卻較單一;云南南部地形復雜,為不同植物類群的生存和繁衍提供了良好的條件,這些植物不但種類豐富,如海芋屬(Alocasia)、兜蘭屬(Paphiopedilum)、開口箭屬(Campylandra)、姜屬(Zingiber)、省藤屬(Calamus)、龍竹屬(Dendrocalamus),而且來源于不同的進化譜系,如天南星科(Araceae)、蘭科(Orchidaceae)、百合科(Liliaceae)、姜科(Zingiberaceae)、棕櫚科(Arecaceae)、禾本科(Poaceae)等,從而導致云南南部具有較高的單子葉植物物種組成和系統發育組成多樣性。
3.2 云南單子葉植物多樣性的保護
對受威脅指數、進化特異性全球瀕危度指數的聯合分析,發現云南西北部和南部是受威脅物種數量最多且受威脅程度最高的區域(見圖2E~F)。已有研究顯示,由于社會發展、人為干擾、外來物種入侵,尤其大型工程帶來的生態環境破壞是植物受威脅的重要原因[48]。此外,Zhang 等[49]的研究表明,由于高額經濟利益驅使,一些重要的野生植物自然資源,如石斛屬(Dendrobium)和兜蘭屬(Paphiopedilum)的種群數量稀少且分布零星,人為采挖及棲息地破碎化是這些類群面臨的最大威脅。除外界客觀因素外,植物自身的特性,如生長緩慢、遺傳變異、萌芽率低等[50]也是導致植物受威脅的主要原因。我國東北、華北分布的大花杓蘭(Cypripedium macranthos)由于基因交流受限,群體內遺傳多樣性較低[51],導致種群瀕臨滅絕。另外,Yildiz 等[52]對土耳其珍稀特有貝母屬植物(Fritillaria aurea)的繁殖生物學研究表明,高度自交不親和及傳粉者依賴性使其生存受到嚴重威脅。
區域性物種豐富度、系統發育多樣性及植物受威脅程度與自然保護地的疊加分析表明,云南西北部、南部及東南部是滇產單子葉植物的重點保護區域(見圖4)。這些地區不但植物的種類、特有化程度及系統發育組成較高,而且受威脅物種的種類組成和區域受威脅程度(東南部除外)也較高。為應對人類活動、氣候極端事件等引起的第六次生物大滅絕,加強對這些地區的生物多樣性保護勢在必行。具體而言,云南南部的單子葉植物譜系多樣性較高,即其現代植物區系構成含有較為多樣的演化支系[53]。通過加強云南南部單子葉植物多樣性的保護,不但可以保護物種、特有種及受威脅物種的多樣性,還可以保護復雜多樣的植物區系演化歷史。而對于云南西北部和東南部來說,雖然單子葉植物的譜系多樣性較低,但這些譜系或主要由新近起源的類群構成(如西北部)[54],或是古老植物類群的孑遺(如東南部)[55],新興類群是未來演化的基礎,古老孑遺類群是植物進化密碼的載體。加強云南西北部、東南部植物多樣性的保護,不僅可以保護植物的種類組成、特有化水平及受威脅物種的多樣性,還可以最大化保護植物區系的演化密碼和進化潛能。盡管目前自然保護地的規劃尚不能涵蓋所有重點保護區(見圖4),但通過提升現有自然保護地的連通性、規劃布局的全能性,仍可以對植物多樣性實施多維度、綜合性的保護(包括演化歷史、進化潛能和演化密碼的保護)。
3.3 演化歷史在生物多樣性保護中的作用
生物多樣性是從宏觀(生態系統多樣性)到微觀(遺傳多樣性)、從群體到個體的自然綜合體[1],生物多樣性保護是多層次、多維度的綜合性保護[56]。然而,由于技術、方法、數據等因素的制約,早期生物多樣性的保護主要關注物種維度和宏觀尺度的保護,忽視了演化歷史(遺傳多樣性)維度的保護。近年來,隨著區域性生命之樹的構建、全球數據的開放共享、機器學習、深度學習、人工智能等新技術、新方法的涌現[37,57],生物多樣性保護既面臨時代的機遇,也面對時代的挑戰,不同保護策略的選擇對保護結果具有根本性影響,基于演化歷史確立的保護優先級和空白區域識別,有助于提高保護策略的效率[58]。如Shan等[59]利用系統發育指標,補充完善了國家重點保護野生藥用物種名錄;Yang等[60]結合系統發育方法,確立了我國種子植物多樣性優先保護區域;因此,未來生物多樣性的保護應在不摒棄物種層面的基礎上,從演化歷史的維度,加強大數據、新技術、新方法在生物多樣性保護中的應用,開展綜合性保護。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
航空世界(2020年10期)2020-01-19 14:36:20
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
