喀斯特地區不同海拔對油茶根際土壤叢枝菌根真菌物種群落的影響
2024-04-01 08:13:36曾維軍劉燕秦維羅文敏
山東農業科學 2024年1期
曾維軍 劉燕 秦維 羅文敏
摘要:本研究旨在了解喀斯特地區不同海拔油茶根際土壤叢枝菌根真菌(arbuscular mycorrhizal fungi,AMF)的種群多樣性及物種分布特征,以提高AMF 在喀斯特地區生態修復中的應用基礎。 以貴州喀斯特高、低海拔地區油茶根際土壤為材料,利用Illumina MiSeq 測序平臺進行高通量測序,分析AMF 物種多樣性及差異。 結果表明,喀斯特地區油茶根際土壤AMF 中球囊霉屬(Glomus)占主要地位,整體豐度為87.28%;低海拔油茶根際土壤AMF 群落特異OTU 數和Shannon、Chao1 指數顯著高于高海拔,物種多樣性較為豐富,且組間AMF 物種聚類較遠;油茶根際土壤有機質和有效磷含量與球囊霉屬真菌豐度呈顯著正相關,相關系數分別為0.58、0.65,表明喀斯特的低磷土壤促進油茶根際球囊霉屬的生長,增加土壤碳庫,促進油茶對磷的吸收利用。其作用機理待進一步研究驗證。
關鍵詞:油茶;喀斯特地區;根際土壤;叢枝菌根真菌;多樣性
中圖分類號:S154.3文獻標識號:A文章編號:1001-4942(2024)01-0105-08
貴州省62%的國土面積為喀斯特地貌,總面積達10.91 萬km2,居全國之首,是全球喀斯特發育最典型、最復雜、景觀類型最豐富的地區之一[1] 。 喀斯特地貌地表干旱缺水,容易造成石漠化等生態退化問題[2] 。 通過植被恢復緩解石漠化是該地區生態修復的關鍵,但土壤的缺水和貧瘠導致植物難以定居、生長緩慢[3] 。 油茶是具有重要經濟和生態價值的油料植物,貴州引種面積逐年增加,栽培過程中也面臨著土壤保水保肥能力弱等問題,制約著油茶果產量和質量的提升。尋找一種提高油茶抗逆性的高效方法成為解決該問題的關鍵[4-5] 。
近年來研究發現,廣泛定植于植物根部的叢枝菌根真菌(AMF) 具有促進宿主營養吸收、抗病、抗旱和改善土壤微生態等功能[6-9] 。 世界上許多國家和地區也規定在特定環境下造林,必須采用菌根化的苗木,以增加移栽的存活率[10-11] 。劉燕等[12] 前期研究也發現,耐旱且油茶果產量穩定的貴州野生油茶,其根系普遍被AMF 定殖。 因此,對AMF 的開發應用是未來研究的熱點和趨勢。
AMF 的開發利用過程中存在兩個主要問題,一是不同AMF 對環境適應性存在差異,二是其高效培養和接種技術有待突破[13-14] 。 貴州喀斯特高、低海拔地區都有野生油茶林分布,不同海拔的油茶可能形成了自己特有的根際AMF 群落。 本研究通過Illumina MiSeq 平臺對喀斯特高、低海拔地區野生油茶根際土樣進行高通量測序,探究AMF 的物種多樣性及其在土壤營養形成中的作用,為AMF 在貴州喀斯特地區油茶綠色種植中的應用提供理論依據。
1 材料與方法
1.1 研究區概況
設置高海拔和低海拔2 個研究區,海拔高度300~2 421 m。 研究區屬亞熱帶季風性和季風性濕潤氣候,年平均氣溫11.5 ~ 19.2 ℃,年降水量926.2~1 340.7 mm,年日照時數為1 328 ~ 1 812h,無霜期180~345 d。 樣區為野生油茶林,林齡為40~50 年,經度在104.08° E~109.07° E,緯度在24.93° N~27.47° N。 每樣地設3 個野生油茶根際土壤采樣點,樣地詳情見表1。
1.2 土壤樣品采集
2021 年8 月,在貴州省威寧、冊亨、從江、玉屏縣的野生油茶分布區內各設置規格為400 m2的3 個相鄰樣方(20 m×20 m),在每個樣方內沿“S”形路徑分別選擇10 株胸徑和樹高相近的油茶樹,鏟除落葉層并挖開上層覆土,剪下細根分枝并輕輕抖動,用毛刷將黏在細根上的土壤刷落到自封袋中。 每個樣方內取10 份根際土壤,混勻后4 ℃冰箱保存,用于測定AMF 多樣性。 樣品的采集與預處理參照?土壤微生物研究原理與方法?[15] 。
1.3 土壤樣品測序
根據soil DNA kit(Omega Bio-tek, Norcross,GA,U.S.)使用說明進行總DNA 提取。 1%的瓊脂糖凝膠電泳檢測基因組DNA 質量,使用Nano ̄Drop2000(Thermo Scientific 公司,美國)測定DNA濃度和純度。 用兩對特異性引物AML1/ AML2(5′ - ATCAACTTTCGATGGTAGGATAGA - 3′;5′-GAACCCAAACACTTTGGTTTCC-3′)和AMV4-5NF/AMDGR(5′-AGCTCGTAGTTGAATTTCG-3′;5′-CCCAACTATCCCTATTAATCAT - 3′) 進行巢式PCR 擴增[16] 。 PCR 反應體系為: 5 × TransStartFastPfu 緩沖液4 mL,2.5 mmol/ L dNTPs 2 mL,上、下游引物(5 mmol/ L)各0.8 mL,TransStart FastPfuDNA 聚合酶0.4 mL,DNA 模板10 mL,超純水補足至20 mL。 擴增程序如下:95 ℃ 預變性3 min;95 ℃變性30 s,55 ℃退火30 s, 72 ℃延伸45 s,
27 個循環;72 ℃延伸10 min;4 ℃保存。 每樣本3個重復。 將同一樣本的PCR 產物混合后使用2%瓊脂糖凝膠回收,利用AxyPrep DNA Gel Extrac ̄tion Kit(Axygen Biosciences,Union City,CA,USA)純化,2%瓊脂糖凝膠電泳檢測其純度,并用Qua ̄ntusTM Fluorometer(Promega, USA)對回收產物進行檢測定量。 使用NEXTFLEX Rapid DNA-SeqKit 對純化后的PCR 產物進行建庫。 委托上海美吉生物醫藥科技有限公司利用Illumina MiSeq 平臺進行測序。
1.4 土壤理化性質檢測
土壤全氮、全磷、全鉀、有效磷、速效鉀含量及pH 值分別參照NY/ T 53—1987、GB/ T 9837—1988、NY/ T 87—1988、NY/ T 1121.7—2014、NY/ T1849—2010、NY/ T - 1241—1999 中方法進行測定;堿解氮采用堿解擴散法測定[17] 。
1.5 數據統計與分析
使用Flash1.2.11 進行pair-end 雙端序列拼接;RDP Classifier2.13 進行序列分類注釋;Use ̄arch11 進行OTU 統計;Mothur1.30.2 進行Alpha多樣性分析,并對樣本間的多樣性指數進行t 檢驗;Mega7 和IQ-TREE 進行進化樹分析和構建;R語言(version 3.3.1)進行物種Venn 圖分析、制作群落柱形圖以及NMDS 分析和作圖。 最后使用Adobe Illustrator2022 修飾圖形。 其中聚類方式采用USEARCH7-uparse 算法,物種分類數據庫為maarjam081/ AMOTU。
2 結果與分析
2.1 不同海拔野生油茶根際土壤AMF 群落OTU組成及結構
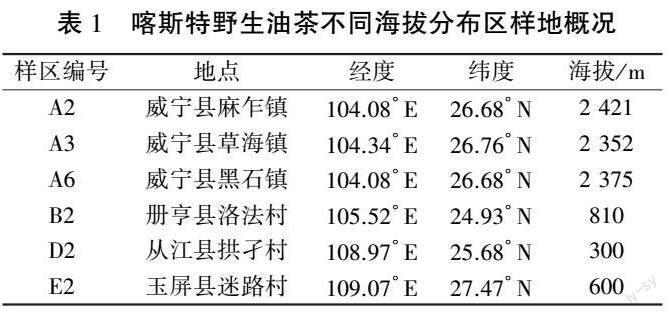
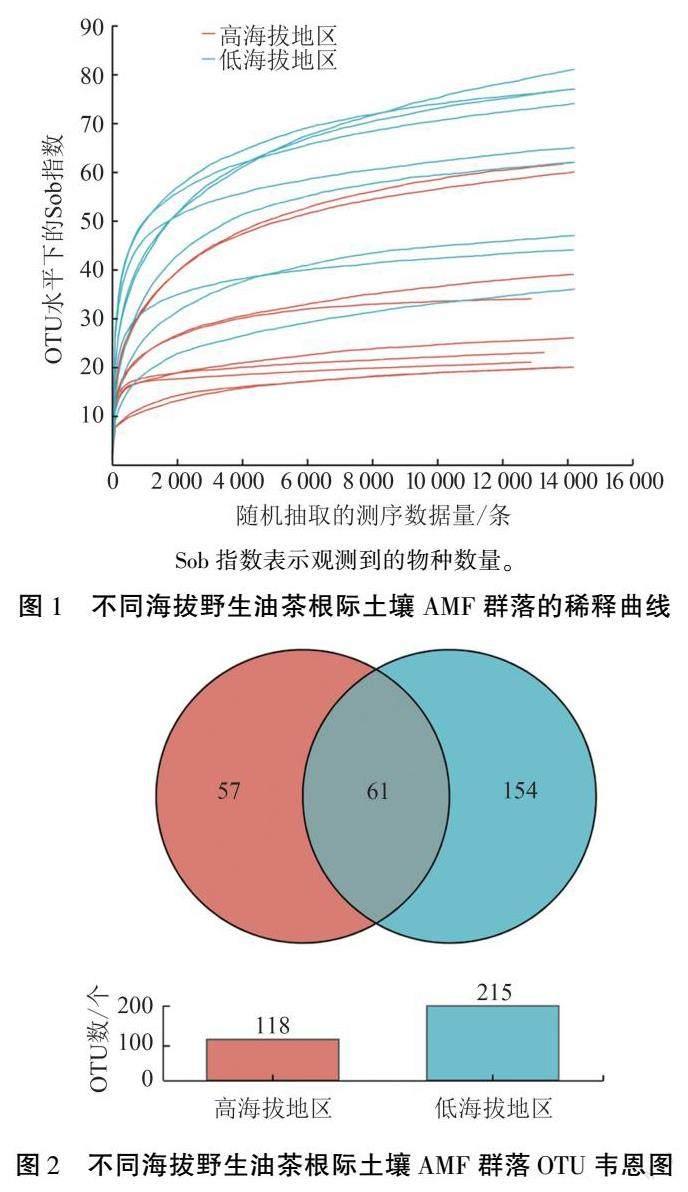
不同海拔野生油茶根際土壤中共獲得367 148條高質量序列,樣品平均序列為20 397條,總堿基數為79 338 214 bp,平均序列長度為216 bp,序列相似度為0.97,OTU 分類置信度為0.7。由稀釋曲線(圖1)可知,抽取的序列條數達到14 223條以上,曲線趨于平坦,表明不同處理的土樣樣本測序數量足夠,測序數據合理。 由Venn圖(圖2)可知,喀斯特高、低海拔地區AMF 群落共有的OTU 數為61 個,占總數的18.32%;高海拔地區特異OTU 數為57 個,占總數的17.12%;低海拔地區特異OTU 數為154 個, 占總數的46.25%。
2.2 不同海拔野生油茶根際土壤AMF 群落結構及多樣性
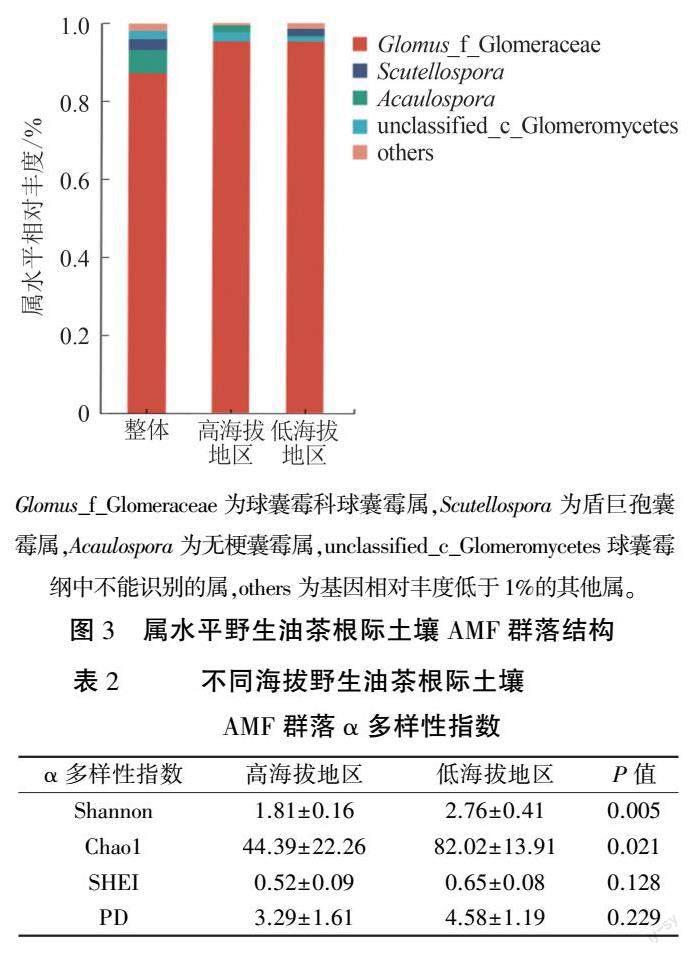
對喀斯特野生油茶根際土壤AMF 群落進行物種注釋得到1 綱5 目8 科14 屬75 種。 由圖3可知,在屬水平上,相對豐度排前4 位的屬分別為球囊霉屬(Glomus,87.28%)、無梗囊霉屬(Acau ̄lospora,5. 97%)、盾巨孢囊霉屬( Scutellospora,2.78%)、球囊菌綱中不能注釋的菌屬(unclassified_c_Glomeromycetes,2.12%),球囊霉屬為優勢屬。喀斯特高、低海拔地區相比,球囊霉屬相對豐度差異不顯著,無梗囊霉屬和球囊菌綱中不能注釋的菌屬在高海拔地區相對豐度較高。 高海拔地區未發現盾巨孢囊霉屬。不同海拔地區野生油茶根際土壤AMF 群落α 多樣性指數如表2 所示,高海拔地區Shannon指數和Chao1 指數均顯著低于低海拔地區,SHEI指數和PD 指數與低海拔地區相比差異不顯著。表明喀斯特高海拔地區的野生油茶根際AMF 群落的多樣性和物種總數均顯著低于低海拔地區。
2.3 不同海拔野生油茶根際土壤AMF 群落PCoA 分析與樣本聚類分析
PCoA 分析可通過降維找出影響野生油茶根際土壤AMF 樣本群落組成差異的潛在主成分。由圖4 可知,在種水平上,主成分1(PC1)與主成分2(PC2)解釋方差分別為51.05%和32.59%;在OTU 水平上,主成分1(PC1)與主成分2(PC2)解釋方差分別為24.60%和22.86%,影響野生油茶根際土壤AMF 群落結構的主導因子明顯(P <0.05)。在種水平上,同一海拔、同一地點根際土壤3 個重復比較聚集,不同地點之間分化明顯,高海拔樣點之間的物種分化較低海拔明顯。 在OTU水平上聚類情況與種水平上相反。
非度量多維尺度分析(NMDS)可將多維空間的研究對象簡化到低維空間進行定位、分析和歸類,揭示復雜背景下的簡單規律。 由圖5 可知,在種水平上,高海拔野生油茶根際土壤樣本組的AMF 群落包含了低海拔土壤樣本組,且Stress<0.05,具有很好的代表性;在OTU 水平上,不同海拔樣本組間存在差異,且Stress<0.2。 PCoA 與NMDS 分析共同驗證了貴州喀斯特高、低海拔間野生油茶根際土壤AMF 群落組成差異明顯,高海拔樣本組涵蓋了低海拔的AMF 分子種。
基于weighted normalized unifrac 距離算法對貴州喀斯特高、低海拔野生油茶根際土壤樣本進行層級聚類分析(圖6)可知,高海拔地區A3 和A6 樣本組的AMF 種群相似,聚為一類。 高、低海拔的AMF 物種組成豐度上有較大差異,說明海拔高低能改變AMF 種群結構。 但是低海拔樣本組E2 與高海拔A2 組的AMF 種群相似,聚為一類。威寧縣草海鎮(A3)與從江縣拱孑村(D2)兩地海拔差為2 052 m,其AMF 群落結構相似性最低。
2.4 AMF 群落與土壤養分因子的相關性分析
通過方差膨脹因子分析(VIF)對7 個影響樣本AMF 菌群組成的土壤養分因子進行篩選,保留多重共線性較小的因子,進行后續研究。 由表3可知,篩選后的土壤養分因子分別為全磷、全鉀、有效磷、速效鉀和有機質,VIF 值均小于10。
通過計算篩選后的土壤養分因子與AMF 物種之間的Pearson 相關系數,獲得相關性熱圖(圖7)。 結果表明,球囊霉屬與土壤有機質和有效磷呈顯著正相關,相關系數分別為0.58、0.65;多樣孢囊霉屬(Diversispora) 和原囊霉屬(Archaeospo ̄ra)與全磷呈極顯著正相關,相關系數分別為0.91、0.90;盾巨孢囊霉屬與速效鉀呈顯著負相關,相關系數為-0.61;無梗囊霉屬(Acaulospora)與全鉀呈極顯著負相關,相關系數為-0.78。
3 討論
除宿主特異性外,AMF 的分布還受氣候、土壤條件、緯度、空間隔離、干擾等多種非生物因子的影響[18-22] 。 研究發現,隨著海拔的升高,青藏高原草甸AMF 的物種豐度逐漸降低[22] ;在宿主植物和土壤條件相似的情況下,AMF 多樣性隨氣溫升高而增加[23-24] 。 本研究發現,低海拔地區油茶根際AMF 群落特異OTU 數、Shannon 指數、Chao1 指數顯著高于高海拔地區,表明AMF 物種總數和多樣性隨海拔的升高而降低;同時AMF 的群落結構也發生了改變,與低海拔地區相比,高海拔地區油茶根際土壤中未發現盾巨孢囊霉屬真菌。 本研究中,海拔高度的改變直接導致兩地區間較大的年平均氣溫差,達7.7 ℃,說明野生油茶根際土壤AMF 的物種結構和多樣性差異也可能是由緯度、海拔差造成的環境溫差導致。 另外,高、低海拔地區的油茶根際AMF 物種可以通過聚類分析分別聚在一起,同時高、低海拔組間又有一定的物種隔離,說明在喀斯特地區海拔差異越大,油茶根際AMF 物種的親緣關系就越遠。
石灰土是喀斯特地區分布最廣的非地帶性土壤,表現出高鈣鎂、低磷鉀和土壤偏堿性的地球化學特征[25] ,而在一定范圍內,低磷土壤會促進AMF 的生長[26-27] 。 研究表明,球囊霉屬真菌能分泌球囊霉素,作為一種非專性蛋白,其主要作用是穩定和增加土壤有機碳庫、改善土壤團聚體,還可以促進植物對磷的吸收,并能與鉛結合在土壤重金屬形態轉化中具有潛在的重要意義[28-30] 。 本研究中,球囊霉屬在AMF 中占據主要地位,并與油茶根際土壤有機質和有效磷含量呈顯著正相關,說明喀斯特的低磷土壤能提高油茶根際球囊霉屬豐度,使土壤中的球囊霉素含量理論上得到提高,從而改善土壤質量,增加土壤碳庫,促進油茶對磷的吸收。
4 結論
貴州喀斯特高、低海拔地區的野生油茶根際土壤AMF 多樣性、群落結構、種水平上的聚類關系均具有顯著差異,其中低海拔地區的AMF 多樣性較豐富,并與高海拔地區存在較為明顯的種群親緣隔離。 另外,喀斯特地區特有的石灰土中缺乏磷、鉀,促進了油茶根際AMF 的生長,尤其是球囊霉屬,其分泌較多的球囊霉素擴撒到土壤中,可改善土壤質量、增加土壤碳庫、促進油茶對磷的吸收利用,從而使油茶根際土壤有機質和有效磷含量增加。 其相關作用機理有待進一步研究驗證。
參 考 文 獻:
[1] Tang J, Tang X X, Qin Y M, et al. Karst rocky desertificationprogress: soil calcium as a possible driving force[J]. Scienceof the Total Environment, 2019, 649:1250-1259.
[2] 王世杰. 喀斯特石漠化———中國西南最嚴重的生態地質環境問題[J]. 礦物巖石地球化學通報,2003,22(2):120-126.
[3] 魏源,王世杰,劉秀明,等. 叢枝菌根真菌及在石漠化治理中的應用探討[J]. 地球與環境,2012,40(1):84-92.
[4] 黃安香,柏文戀,鄔能英,等. 貴州油茶主產區土壤重金屬環境質量狀況及風險評價[J]. 環境化學, 2023,42(4):1325-1336.
[5] 柏文戀,黃安香,楊守祿,等. 貴州油茶主產區土壤養分特征及綜合肥力評價[J]. 西南農業學報,2022,35(9):2096-2106.
[6] 劉代軍,涂波,施松梅,等. 石漠化地區的生態危機及菌根桑生物修復潛力研究進展[J]. 中國巖溶,2012,31(2):185-190.
[7] Duru M, Cruz P P, Raouda A H K, et al. Relevance of plantfunctional types based on leaf dry matter content for assessingdigestibility of native grass species and species ̄rich grasslandcommunities in spring[J]. Agronomy Journal, 2008, 100(6):1622-1630.
[8] 姜海燕,李敏. 西瓜-辣椒輪作體系AMF 和PGPR 防控線蟲的效果[J]. 山東農業科學,2021,53(4):131-134.
[9] 康佳,梁夕金,楊文龍,等. 叢枝菌根真菌(AMF)對鹽堿地花生根系土壤微生物的影響[J]. 山東農業科學,2022,54(7):77-84.
[10] Skuji?? J, Allen M F. Use of mycorrhizae for land rehabilitation[J]. MIRCEN Journal of Applied Microbiology and Biotechnol ̄ogy, 1986, 2:161-176.
[11] 魏源,王世杰,劉秀明,等. 喀斯特地區叢枝菌根真菌遺傳多樣性[J]. 生態學雜志,2011,30(10):2220-2226.
[12] 劉燕,曾維軍,秦維,等. 威寧短柱油茶叢枝菌根真菌和深色有隔內生真菌定殖特征及其與果實經濟性狀的相關性[J]. 江蘇農業科學,2023,51(22):145-153.
[13] 楊佳絨,戴冬,陳俊芳,等. 叢枝菌根真菌多樣性對植物群落構建和稀有種維持的研究進展[ J]. 植物生態學報,2023,47(6):745-755.
[14] 儲薇,郭信來,張晨,等. 叢枝菌根真菌-植物-根際微生物互作研究進展與展望[J]. 中國生態農業學報:中英文,2022,30(11):1709-1721.
[15] 林先貴. 土壤微生物研究原理與方法[M]. 北京:高等教育出版社,2010:112-134.
[16] Van Geel M, Busschaert P, Honnay O, et al. Evaluation of sixprimer pairs targeting the nuclear rRNA operon for characteriza ̄tion of arbuscular mycorrhizal fungal (AMF) communities u ̄sing 454 pyrosequencing[J]. Journal of Microbiological Meth ̄ods, 2014, 106:93-100.
[17] 馬獻發. 土壤學實驗[M]. 北京:中國林業出版社,2020:218-234.[18] Bever J D. Host ̄specificity of AM fungal population growthrates can generate feedback on plant growth [ J]. Plant andSoil, 2002, 244:281-290.
[19] 楊海水. 宿主植物對叢枝菌根真菌的影響———共生功能、地理分布及多樣性[D]. 杭州:浙江大學,2013.
[20] 楊蕊毓,鄧錡璋,田麗平,等. 不同生境下川麥冬根圍土壤叢枝菌根真菌多樣性[J]. 西北植物學報,2022,42(1):145-153.
[21] 朱亮,郭可馨,藍麗英,等. 亞高山森林類型轉換對土壤叢枝菌根真菌多樣性的影響[J]. 生態學雜志,2020,39(12):3943-3951.
[22] Gai J P, Tian H, Yang F Y, et al. Arbuscular mycorrhizal fun ̄gal diversity along a Tibetan elevation gradient[J]. Pedobiolo ̄gia, 2012, 55(3):145-151.
[23] Koske R E. Distribution of VA mycorrhizal fungi along a latitu ̄dinal temperature gradient[J]. Mycologia, 1987, 79:55-68.
[24] 石國璽,王芳萍,馬麗,等. 長期、短期增溫對高寒草甸AM真菌群落結構的影響[J]. 草地學報,2021,29(增刊1):179-189.
[25] 羅緒強,王程媛,楊鴻雁,等. 喀斯特優勢植物種干旱和高鈣適應性機制研究進展[J]. 中國農學通報,2012,28(16):1-5.
[26] Tawaraya K, Saito M, Morioka M, et al. Effect of phosphateapplication to arbuscular mycorrhizal onion on the developmentand succinate dehy ̄drogenase activity of internal hyphae[J].Soil Science and Plant Nutrition, 1994, 40(4):667-673.
[27] 吳強盛,夏仁學,鄒英寧. 柑橘叢枝菌根真菌生長與根際有效磷和磷酸酶活性的相關性[J]. 應用生態學報,2006,17(4):4685-4689.
[28] 張健,徐明,吳麗豐,等. 干擾對黔中地區馬尾松林土壤球囊霉素的影響[J]. 森林與環境學報,2022,42(1):63-70.
[29] 張健,徐明,王陽,等. 黔中地區不同馬尾松群叢土壤球囊霉素分布特征[J]. 生態環境學報,2021,30(12):2303-2308.
[30] 甘佳偉,韓曉增,鄒文秀. 球囊霉素及其在土壤生態系統中的作用[J]. 土壤與作物,2022,11(1):41-53.
