食物過敏的膳食危險因素及其調控作用機制研究進展
2024-04-08 02:28:30孫佳晨張巧智李華韜傅玲琳
食品科學 2024年5期
關鍵詞:研究
孫佳晨,張巧智,*,李華韜,張 巖,傅玲琳,*
(1.浙江工商大學食品與生物工程學院,浙江 杭州 310018;2.河北省食品檢驗研究院,河北省食品安全重點實驗室,國家市場監管重點實驗室(特殊食品監管技術),特殊食品安全與健康河北省工程研究中心,河北 石家莊 050000)
食物過敏是指由機體免疫系統對某些食物蛋白質的超常反應,發病時可累及皮膚、呼吸道、胃腸道和神經系統等周身多個系統,嚴重時可發生休克甚至危及生命。在美國、加拿大和新西蘭等西方國家,食物過敏在成人中的發病率約為2%,而在兒童中則高達8%[1];在我國,據不完全統計,食物過敏在嬰幼兒中的發病率約為3.5%~11.1%,且這一發病率近年來呈不斷上升趨勢[2]。食物過敏已經成為全球關注的食品安全和公共衛生問題,被稱為繼哮喘之后的“第二波”過敏流行病。食物過敏不僅會嚴重影響患病個體的身體健康和生活質量,同時也對家庭、醫療和公共衛生管理造成了愈加嚴重的負擔。
研究發現,食物過敏的發生主要與遺傳因素、環境因素以及二者的相互作用有關[3-4]。雖然遺傳因素在食物過敏的發病機制中占有一定作用,但考慮到短時間內人類遺傳基因的改變不足以解釋過敏狀況的大幅上升,環境因素特別是膳食因素被認為是導致食物過敏風險升高的主要驅動力[5-8]。在過去幾十年間,全球城市化與工業化進程加速,人們的飲食結構發生了重大變化。近年來,被證明與食物過敏發生有關的膳食成分改變主要包括致敏原的暴露時間、特定免疫活性成分的攝入不足、有益微生物暴露減少(主要體現為益生菌膳食攝入),以及不健康飲食(如高脂、高糖、低纖維)導致的肥胖等。這些可能增加食物過敏發生可能性的膳食因素統稱為膳食危險因素[4,9]。這其中,腸道菌群在調控機體免疫穩態中具有重要作用,膳食危險因素對食物過敏的影響也與其作用于腸道菌群進而影響宿主的免疫應答密切相關。本文系統綜述了有關食物過敏膳食危險因素的相關研究,從食物過敏的機理、不同危險因素的免疫學機制及其人群隊列研究和干預實驗的結果等方面展開,系統回顧了近年來國內外的相關研究進展,并著重針對膳食和菌群以及免疫系統的相互作用進行了探討,以期為食物過敏的預防和控制,以及以腸道菌群為靶點的食物過敏機制研究和調控策略提供借鑒和新思路。
1 免疫耐受建立與食物過敏
1.1 口服免疫耐受的建立
正常情況下,機體免疫系統能夠正確區分致病性抗原與無害的環境抗原,如食物中的營養物質,這一免疫耐受的建立過程與腸相關淋巴組織(gut-associated lymphatic tissue,GALT)密切相關[10]。在腸道中,GALT中淋巴細胞約為1012個[11],這些細胞間的協同作用實現了對體內常駐細菌和食物蛋白炎性反應的限制,同時對可能導致組織損傷或疾病發生的病原體進行有效識別和響應[12]。
口服免疫耐受的建立在很大程度上是由腸道固有層中的抗原提呈細胞(antigen-presenting cells,APCs)驅動的[20]。人體攝入食物后,通過唾液和胃腸液中的消化酶對食物中的蛋白質進行消化[13]。被消化的食物抗原會以多種方式透過腸上皮屏障,如胞吐作用、M細胞內吞抗原,以及被髓樣細胞(如樹突狀細胞(dendritic cells,DCs)和巨噬細胞)捕獲等[14-16]。抗原透過腸上皮屏障后,經過如DCs等傳統APCs或如庫普弗細胞(Kupffer cell)等非傳統APCs處理后進入腸系膜淋巴結(mesenteric lymph node,MLN),然后經主要組織相容性復合體(major histocompatibility complex,MHC)分子傳遞給T細胞,促進T細胞分化[16-17]。
在免疫耐受建立的過程中,抗原首先被CX3CR1+髓樣細胞捕獲從而穿過腸上皮屏障。CX3CR1+細胞的樹突延伸到腸上皮細胞之間,在腸腔中采集抗原樣本并轉移給CD103+DCs。一部分DCs從固有層遷移到MLN,在這里CD103+DC表達轉化生長因子β(transforming growth factor-β,TGF-β)和視黃酸,從而誘導初始T細胞分化為抗原特異性叉頭狀P3(forkhead box protein 3,Foxp3+)調節性T(regulatory T,Treg)細胞[18-19]。此外,CD103+DC產生的視黃酸也可誘導整合素α4β7的表達,α4β7可誘導Treg細胞歸巢到腸道,進一步抑制免疫反應。與此同時,視黃酸和DCs的相互作用在次級淋巴組織中也可誘導B細胞向分泌免疫球蛋白(immunoglobulin,Ig)A的漿細胞分化,最終達到IgA抗體的類別轉換(圖1A)[20]。
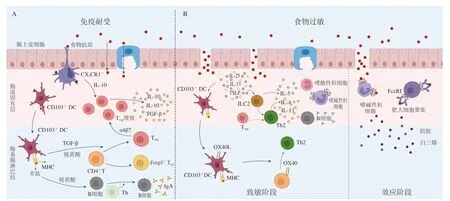
圖1 免疫耐受(A)和食物過敏(B)的機理示意圖Fig.1 Schematic diagram of the mechanisms of immune tolerance (A) and food allergy (B)
1.2 食物過敏的分子機理
在食物過敏患者體內,上述口服耐受的形成過程遭到了破壞。根據發病機制的不同,食物過敏可以分為IgE介導、非IgE介導和混合介導類型,其中由IgE介導的反應是最具特征的食物過敏類型,其免疫學機理也較為清楚[17],主要分為兩個階段:致敏階段和效應階段。個體初次暴露于致敏原時為致敏階段,再次暴露時為效應階段。
在致敏階段,上皮屏障的損傷導致食物蛋白抗原的透過率增加,誘導上皮細胞分泌IL-25、IL-33和胸腺基質淋巴細胞生成素(thymic stromal lymphopoietin,TSLP)等細胞因子,上述因子使DCs中OX40L的表達上調[16],并誘導ILC2增殖,進而激活初始T細胞向Th2細胞分化,Th2細胞分泌IL-4、IL-5和IL-13等細胞因子,其中IL-5可招募嗜酸性粒細胞,IL-4和IL-13可促進B細胞發生類別轉換產生特異性IgE(specific IgE,sIgE),并促進記憶B細胞的產生[21]。B細胞成熟后分化為漿細胞,產生大量致敏原sIgE抗體,這些抗體與肥大細胞和嗜堿性粒細胞表面的高親和力FcεRI受體結合,使機體處于致敏狀態。上述致敏階段可能發生在胃腸道、皮膚和呼吸道等處。當處于致敏階段的機體再次接觸致敏原時,免疫反應進入效應階段,致敏原衍生的表位通過與免疫效應細胞表面的sIgE結合,引發IgE交聯,觸發細胞脫顆粒反應并釋放組胺、白三烯等炎性介質,進而引起局部或全身的過敏癥狀(圖1B)[10,21]。
1.3 腸道微生物調節食物過敏的機理
越來越多的證據表明,腸道微生物在食物過敏的發病機制中起著關鍵作用。已有報道指出,食物過敏與健康人群的腸道微生物組存在明顯不同。如一項前瞻性研究調查了44 名新生兒,發現在過敏兒童腸道中的需氧菌較多,尤其是梭菌和金黃色葡萄球菌數量多,而乳酸桿菌和雙歧桿菌數量少,這表明過敏反應的發生與腸道菌群的結構變化有關[22]。此外研究人員對226 名兒童進行了隊列研究,發現嬰兒在3~6 個月時的腸道微生物構成與8 歲時牛乳過敏的狀況有關,其中腸桿菌和擬桿菌門的富集會導致牛乳過敏反應[23]。另外一項對166 名嬰兒的隊列研究也表明,嬰兒3 個月時的腸道細菌種類每增加25%,1 歲時食物過敏的風險會降低55%[24]。綜上研究結果表明,腸道菌群的失衡很大可能先于過敏癥狀的發生。進一步在動物模型中發現,腸道微生物中的特定菌群可獨立影響菌群移植小鼠的過敏表型,無菌小鼠或菌群多樣性較低的小鼠易發生IgE抗體類別轉換并出現食物過敏癥狀;通過口服特定種類的細菌也可起到緩解食物過敏癥狀的效果,可見腸道微生物是決定機體免疫應答和耐受形成的重要因素,或可作為潛在的食物過敏預防和干預手段[25-29]。
以上研究有力證明了食物過敏與腸道微生態間的因果關聯。針對微生態參與調節食物過敏的機理,已有報道主要包括以下幾個方面(圖2):1)腸道菌群通過與DCs、先天淋巴細胞、嗜堿性粒細胞和巨噬細胞等相互作用,誘導IL-22等細胞因子的分泌,刺激上皮再生,維護腸道屏障功能的完整性;2)腸道菌群通過其代謝產物間接作用于腸黏膜免疫系統,如短鏈脂肪酸(short-chain fatty acids,SCFAs)可作用于腸上皮細胞激活炎癥小體產生IL-18和抗菌肽,強化腸上皮屏障功能;這些代謝產物也可能直接作用于初始T細胞,促使其分化為Foxp3+Treg細胞,進而抑制炎癥反應并促進免疫耐受;3)微生物直接或者通過DCs傳導信號作用于初始T細胞,使其分化為Th1或Treg細胞。Th1細胞通過分泌干擾素γ(interferon-γ,IFN-γ)抑制Th2的分化,維持腸道內的Th1/Th2細胞平衡,Treg細胞能通過分泌TGF-β、IL-10等抗炎因子,并且刺激B細胞產生IgA和IgG4,從而維持免疫耐受[30-32]。
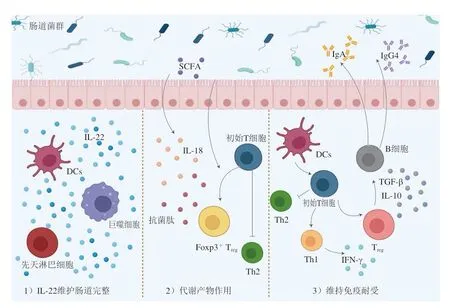
圖2 腸道微生物調節食物過敏的機理Fig.2 Mechanism of action of intestinal microorganisms in regulating food allergy
2 致敏原早期暴露
過敏性食物引入的時間是影響食物過敏發生的重要因素。以往人們認為,1 歲以下的嬰兒由于胃腸功能發育不完全,應盡可能推遲引入花生、雞蛋、魚類等容易過敏的食物。但近年來,越來越多的證據表明,在嬰兒生命早期引入易過敏的食物反而會降低后期發生食物過敏的風險[33]。這其中比較有力的證據包括:
一項里程碑式的研究——關于花生過敏的早期花生學習(learn early about peanut,LEAP)表明,與避免早期引入花生相比,將花生早期引入食物過敏高危嬰兒可有效減少花生過敏的發生[34]。LEAP-On的后續研究也表明,與對照組相比,在出生后前5 年經常食用花生的兒童在停止食用花生1 年后,對花生持續無反應[35]。其他致敏原也與花生有著類似的調查結果,例如一項綜述系統性地評價了7 項隨機對照實驗,觀察雞蛋、花生和牛乳早期引入對于預防食物過敏的作用,其中2 篇關于花生早期引入的隨機對照實驗結果顯示早期引入花生能降低花生過敏,5 篇研究結果一致顯示早期引入雞蛋可以降低雞蛋過敏的發生率[36],再如英國的一項Enquiring About Tolerance(EAT)研究招募了1303 名嬰兒,并將受試嬰兒分成了早期引入組(即3 個月時引入易過敏食物)和標準引入組(即6 個月時引入易過敏食物),結果發現早期引入組易感嬰兒發生多種食物過敏的幾率均低于標準引入組[37-38]。
盡管如此,也存在著不同的研究結果。一項芬蘭的出生隊列調查了6209 名嬰兒發現了相反的結論,在出生后的前幾天內引入牛乳蛋白與牛乳過敏的風險增加相關[39]。另外也有研究表明致敏原的暴露時間與食物過敏的發生無顯著關聯,如一項新加坡的出生隊列研究招募了1152 名來自中國、馬來和印度族裔的母親,并前瞻性地追蹤了她們的后代,結果表明引入致敏性食物的時間與食物過敏風險之間沒有關聯[40]。最后,另一項系統綜述和薈萃分析對特定食物進行研究,也表明早期引入牛乳與牛乳致敏風險無關[41]。
另外,不只是單一的膳食暴露會影響食物過敏,嬰兒早期攝入的膳食多樣性也會影響食物過敏的發生發展[42]。在一項來自5 個歐洲國家的研究中,Roduit等[42]觀察到嬰兒出生后第一年飲食多樣性的減少與6 歲時食物過敏的風險增加有關。在另一項懷特島的嬰兒出生隊列研究中也發現[40],嬰兒6 個月和12 個月時的飲食多樣性與10 歲前發生食物過敏的幾率間存在顯著關聯。出生后6 個月內每增加一種食物,在10 歲前發生食物過敏的幾率會降低10.8%;出生后12 個月內每增加一種食物,在10 歲前食物過敏的可能性降低33.2%[43]。而對于飲食多樣性影響食物過敏發生的作用機制,D’Auria等[44]認為,飲食多樣性可能會通過增加腸道微生物的種類、調控腸道黏膜內環境的穩態維持免疫耐受,鑒于此,有關飲食多樣性與腸道微生物群的多樣性關聯分析或可作為未來人群隊列和動物模型研究的切入方向。
鑒于以上證據,近年來,各地國家和機構相繼頒布了有關致敏性食物引入時間的指南,匯總于表1。總體上,各指南建議嬰兒在出生后4~6 個月可以開始攝入固體食物,而晚于6 個月引入易致敏食物反而會增加食物過敏的風險。

表1 各機構關于致敏性食物引入時間的指南Table 1 Guidelines on the introduction time of allergic foods from various institutions
3 膳食免疫調節活性成分與食物過敏
近年來,飲食結構的顯著變化使得研究人員猜測,除了能夠作為抗原的致敏蛋白以外,飲食中的其他特定成分,如維生素、脂質和抗氧化物質等,這些成分具有調節機體免疫系統的潛力,它們可能通過與變應原相關或無關的途徑影響食物過敏的發生。
3.1 VD
早在1983年,研究人員在淋巴細胞中發現了VD受體,這使得人們首次認識到了VD的免疫學意義[56]。進一步研究發現,VD對固有免疫和適應性免疫具有多重調控作用,可以促進形成免疫耐受、減輕炎癥和保護腸上皮屏障的完整性。目前有關VD促進免疫耐受的機制,研究人員提出了多種可能,包括調節免疫細胞的比例或功能,如誘導耐受性DCs和Treg細胞以及抑制B細胞產生IgE;誘導天然免疫細胞合成抗菌肽從而抑制過度炎癥;改變腸道微生物組的組成和腸道上皮的完整性等[57]。其中,就腸道菌群而言,VD扮演著免疫系統和微生物群落間的中介角色,因VD或VD受體的改變與微生物組失調有關,而微生物組失調又與腸道炎癥和特異性疾病有關[58]。研究發現,VD缺乏或VD受體失活與微生物群落多態性失調有關,這一失調可導致厭氧桿菌屬和變形桿菌屬的增加和炎癥性疾病[59]。例如,克羅恩病患者補充VD會促進腸道微生物組中某些細菌物種的生長,如另枝菌屬(Alistipes)、巴氏桿菌屬(Barnesiella)、羅斯氏菌屬(Roseburia)、厭氧棍狀菌屬(Anaerotruncus)和罕見小球菌屬(Subdoligranulum)[58]。再如,在多發性硬化癥患者中,VD與大量促進免疫耐受的阿克曼菌屬(Akkermansia)以及糞桿菌屬和球菌屬有關,這幾種細菌都能產生丁酸鹽,而丁酸鹽可起到抗炎的作用[60]。
近年來,愈來愈多的證據表明,人群中VD的不足和食物過敏的發病率上升有關;據此,研究人員提出了VD缺乏假說,認為VD不足是食物過敏等過敏性疾病增加的原因之一[61]。支持這一假說的早期證據是在光照、緯度和出生季節與食物過敏風險之間發現的聯系,例如在遠離赤道的地區兒童食物過敏的發生更為普遍[62],又如冬季出生的嬰兒患食物過敏的風險更高等[63]。這些證據表明了由光照暴露不足導致的VD缺乏在食物過敏發病中的潛在作用[64]。此外,Nwaru等對931 對母子進行了為期5 年的隊列研究,發現母親在懷孕期間平均每日補充1.1 μg VD與后代5 歲時較低的食物過敏風險有關[65]。另外,Allen等在對5276 名1 歲嬰兒展開調查后發現,血清中VD不足(25-羥基VD<50 nmol/L)的嬰兒與VD充足(25-羥基VD>50 nmol/L)的嬰兒相比,1 歲時發生花生過敏的可能性高11 倍,發生雞蛋過敏可能性高3 倍,對兩種或兩種以上食物過敏的幾率增加10 倍[66];進一步對該人群中54 名雞蛋過敏的兒童進行長期追蹤,發現兒童1 歲時血清中25-羥基VD的水平與4 歲時建立雞蛋免疫耐受的幾率呈正相關[67]。
然而,也有研究發現,過量補充VD可能會引起食物過敏。Milner等進行了一項超8000 人的隊列研究發現,出生后6 個月內補充VD的嬰兒在3 歲時患食物過敏的幾率反而有所增加[68]。類似地,Rosendahl等研究指出,嬰兒臍帶血中25-羥基VD的水平與1 歲時發生食物過敏等相關疾病的風險呈正相關[69]。鑒于補充VD與食物過敏風險之間的差異結果,研究人員提出了U形曲線假設,認為VD與食物過敏的發病率之間可能存在U型關系,即VD過少或過多均會增加食物過敏的風險[70]。Hypp?nen等對英國7288 名志愿者進行生物醫學評估,發現血清中的25-羥基VD的濃度與IgE水平呈非線性關系[71],這也支持了該假說。但是,目前尚沒有充足數據進一步證實上述關系,為了進一步了解VD在食物過敏中的確切作用,一方面,需要破譯VD在先天免疫系統和適應性免疫系統中的細胞和分子機制;另一方面,食物過敏中VD和腸道菌群間的相互作用和關聯也值得進一步研究。另外,為了闡明VD在食物過敏預防和治療中的潛力,應在特征明確的隊列中補充VD的隨機對照實驗。
3.2 多不飽和脂肪酸(polyunsaturated fatty acids,PUFA)
在過去幾十年間,人類的飲食結構發生了巨大改變,尤其是PUFA的攝入水平發生了顯著變化。PUFA中理想的膳食ω-3∶ω-6平衡為1∶3~1∶5,然而在西方飲食中,由于人們對含有ω-6 PUFA的人造黃油、植物油(如葵花籽油、大豆油和玉米油)等食物的消耗量增大,而對含有ω-3 PUFA的深海魚、堅果等食物的消耗量減小,導致膳食中ω-3∶ω-6達到了1∶10至1∶30甚至更高,比值嚴重失衡[72]。據此,研究人員提出假設,近年來食物過敏患病率的增加可能與飲食中ω-6與ω-3 PUFA攝入量的不平衡有關[72]。該假設認為,PUFA能參與調節機體免疫系統和炎癥反應,例如在常見的PUFA中,ω-6 PUFA(如亞油酸、花生四烯酸(arachidonic acid,AA))通常具有促炎作用,而ω-3 PUFA(如亞麻酸、硬脂酸、長鏈二十碳五烯酸、二十二碳五烯酸(eicosapentaenoic acid,EPA)、二十二碳六烯酸(docosahexaenoic acid,DHA)等)則被認為可以減輕炎癥和過敏反應。這其中,亞油酸和AA可導致前列腺素E2(prostaglandin E2,PGE2)的合成和分泌,PGE2作用于T淋巴細胞抑制IFN-γ的產生,進而誘導B淋巴細胞成熟并分泌IgE;在上述過程中,ω-3 PUFA中如EPA和DHA可與AA產生競爭,減少PGE2等促炎因子的形成,發揮抗炎作用[72]。
盡管尚無定論,但大多數針對母親在產前或是嬰兒在產后攝入ω-3 PUFA與嬰兒食物過敏之間聯系的研究發現,ω-3 PUFA具有保護人體免受食物過敏的有益作用。例如Dunstan等開展了一項隨機對照實驗,招募的孕婦從妊娠20 周至分娩期間服用魚油(3.7 gω-3 PUFA/d)或安慰劑,發現服用魚油母親的嬰兒在1 歲時發生雞蛋過敏的可能性是對照組的1/3[73]。Palmer等的實驗發現了類似現象,孕婦從妊娠第21周至生產,期間每天補充0.1 g二十碳五烯酸和0.8 g DHA或安慰劑,發現補充ω-3組嬰兒的食物過敏發生率顯著低于安慰劑組[74]。為了研究產后嬰兒的魚油攝入與食物過敏的關聯,Clausen等對冰島兒童進行了一項從出生到2.5 歲的前瞻性隨訪,結果顯示嬰兒出生后前6 個月食用魚油可降低發生食物敏感的風險[75]。Vahdaninia等[76]針對7 個國家10 項隨機對照實驗進行了系統回顧和薈萃分析,發現懷孕期間開始攝入ω-3 PUFA可能會降低雞蛋和花生過敏的發生率。
然而也存在著不一致的結論,如在一項針對420 名高風險嬰兒進行的雙盲隨機對照實驗中,與對照組(補充橄欖油)相比,每日補充魚油雖然增加了嬰兒體內ω-3 PUFA(DHA和EPA)的水平,但并不能起到預防過敏性疾病的作用[77]。Gunaratne等[78]結合8 項不同調查的結果,其中涵蓋了3366 名女性和3175 名兒童,得出結論認為,孕期補充ω-3 PUFA對預防后代發生食物致敏和其他過敏性疾病的證據比較有限。
鑒于以上證據,有關ω-3和ω-6 PUFA是否具有預防和改善食物過敏的效果尚未得到確切的結論[79];目前,在世界不同國家和機構頒布的食物過敏預防指南中尚沒有關于PUFA的指導意見。以預防食物過敏為目的補充ω-3 PUFA的時間、劑量、受益個體的遺傳差異以及ω-3 PUFA發揮作用的具體途徑還需進一步深入研究。
3.3 抗氧化物質
近幾十年來,因受到西方飲食的影響,人們攝入動物性食物的比例升高而果蔬類食物的攝入嚴重不足,這會導致人體從食物中獲得的抗氧化物質減少。眾所周知,天然的抗氧化物質在果蔬中含量較為豐富,如葡萄、橙子、西紅柿等;而非天然抗氧化劑大多是以食品添加劑的形式被人體攝入的,如亞硫酸鈉和苯甲酸等食品防腐劑[80]。
人體對富含抗氧化劑的新鮮果蔬的攝入量減少可能與食物過敏的發生有關。提出這一設想的原因是基于體內過量的活性氧可誘發過敏性炎癥,而抗氧化劑則可通過防止氧化應激和減輕活性氧誘導的損傷對人體發揮有益作用[81]。此外,在患炎癥性腸病的動物模型中發現天然抗氧化劑白藜蘆醇會導致腸道中的雙歧桿菌和乳酸桿菌數量增加、腸壁纖維化減輕以及中性粒細胞浸潤和促炎細胞因子釋放減少,這也可能是抗氧化劑起到預防食物過敏的原因之一[82]。
有關抗氧化劑與食物過敏之間的關聯研究不多,如一項前瞻性研究調查了母親孕期攝入抗氧化劑(如β-胡蘿卜素、VC、VE、銅和鋅)的含量與后代嬰兒1 歲時患各類過敏性疾病之間的關系。結果發現銅和VC的攝入量與后代發生食物過敏的概率呈負相關,而其他抗氧化劑的攝入量則與過敏性疾病無顯著關聯[83]。然而也有人持反對意見,認為攝入富含抗氧化劑的食物可能會增加食物過敏的易感性。雖然抗氧化劑可以中和促氧化物質,但過量的抗氧化劑介導的抗氧化應激也會對免疫系統產生不利影響,例如抑制Th1型免疫反應并激活Th2反應等[84]。在一項芬蘭的前瞻性出生隊列研究中發現,母親孕期攝入β-胡蘿卜素的水平與子代牛乳過敏的風險增加相關[85]。然而目前有關抗氧化劑與食物過敏之間關聯的流行病學調查研究較為有限,可能是因為設定不含抗氧化劑的食物對照組比較困難。在未來的研究中可以著重于動物實驗,研究攝入特定抗氧化物質對食物過敏的影響。
綜上所述,盡管一些研究報道了母親在懷孕期間攝入抗氧化劑與后代發生食物過敏之間的潛在聯系,但現有證據并不充分,因此在向孕婦提出建議之前,需要在動物模型中重復流行病學研究,在此基礎上開展大規模的臨床隊列研究,以確認抗氧化劑對食物過敏干預的有效性和功效持續性并提出有效的膳食建議。
4 膳食益生菌和益生元的暴露與食物過敏
腸道微生物群的結構和功能是影響食物過敏發生發展的重要因素[86]。在飲食成分中,膳食纖維、母乳低聚糖、益生菌和益生元等已被證明會影響腸道微生物群的結構和功能。這其中,益生菌作為一類對宿主有益的活性微生物,服用后定植于人體腸道,可起到改善機體微生態失衡、調控和緩解食物過敏的作用[87]。采用益生菌干預對食物過敏進行預防或治療的人群實驗研究中,最常被作為膳食補充劑攝入的益生菌多數為乳酸桿菌屬(Lactobacillusspp.)和雙歧桿菌屬(Bifidobacteriumspp.)菌株。關于益生菌和食物過敏的研究大多集中于牛乳過敏的人群,如Canani等對153 名1~12 個月的牛乳過敏兒童補充鼠李糖乳桿菌GG(L.rhamnosusGG,LGG),進行了為期12 個月的臨床試驗,發現補充LGG可以治療牛乳過敏的癥狀并縮短耐受性獲得的時間[88],Cukrowska等給151 名2 歲以下患有牛乳過敏兒童補充由鼠李糖乳桿菌LOCK 0900、鼠李糖乳桿菌EOCK 0908和干酪乳桿菌LOCK 0918組成的益生菌制劑或安慰劑,進行3 個月的干預隨后隨訪9 個月,發現益生菌組對牛乳蛋白的耐受性明顯高于安慰劑組[89]。然而Hol等對119 名1~6 個月的牛乳過敏兒童補充干酪乳桿菌和乳酸雙歧桿菌,發現水解配方奶粉中添加這兩種益生菌對牛乳過敏患兒的牛乳耐受性無促進作用[90]。可見,益生菌對牛乳過敏的緩解作用可能具有菌株特異性。益生菌對其他食物過敏類型的干預研究也有報道,如Tang等進行了一項花生過敏兒童的臨床試驗,采用LGG干預聯合花生口服免疫療法治療18 個月,發現與安慰劑相比,接受益生菌聯合治療的受試者對花生的脫敏率更高[91]。
有關益生菌調控食物過敏的免疫學機制,研究人員指出,益生菌的抗炎作用可以預防食物過敏性反應[92],這些有益微生物也可通過作用于腸道免疫系統發揮非免疫性保護作用。首先,益生菌可以直接阻斷腸道病原微生物的生長繁殖:益生菌可通過產生抗菌物質直接抑制病原體的生長,如細菌素(如抗生素)和多種SCFAs(乙酸、乳酸和丙酸)[93],它們還可通過黏附腸上皮細胞進而阻斷病原體與腸上皮細胞的結合[94];其次,益生菌可以通過不同的途徑增強腸道屏障的完整性:如植物乳桿菌可通過上調緊密連接蛋白claudin-1、occludin、JAM-1和ZO-1的表達水平提高腸上皮細胞的屏障完整性,它們還可通過激活抗凋亡通路和抑制促凋亡通路[95-96],促進腸上皮細胞的存活并抑制細胞因子誘導的上皮損傷;最后,益生菌可以通過調節腸道微生態的結構組成進而發揮有益作用:如益生菌干預后小鼠的腸道菌群分析表明,益生菌可通過增加腸道中糞球菌屬和理研菌屬的相對豐度發揮保護作用[97],再如人群研究顯示,產前補充LGG可調節嬰兒腸道雙歧桿菌的定植[98],使其更接近母乳喂養健康嬰兒的腸道菌群構成,此外Canani等對2 歲以下的牛乳過敏嬰兒進行LGG和深度水解配方奶粉聯合干預,發現干預后嬰兒對牛乳的耐受性提高且其糞便中布勞特氏菌屬、羅斯氏菌屬和糞球菌屬得到顯著富集,由此得出結論,深度水解奶粉和LGG促進牛乳過敏嬰兒耐受的部分原因是通過影響嬰兒腸道的細菌群落結構達到的[99]。
另一方面,益生元是一類“通過選擇性刺激存在于結腸中細菌的活性,從而改善宿主健康,對宿主產生有益影響的不易消化的食物成分”[100]。典型的益生元如膳食纖維類,常見的有菊糖、低聚半乳糖、低聚果糖和低聚木糖等。這些食品補充劑對腸道微生物群和機體免疫系統有許多復雜且有益的影響,可以用于緩解腸道免疫系統中的某些疾病[101]。一項動物研究把可可作為食用的益生元[102],在這項研究中,與標準飲食組的大鼠(無論是口服致敏還是非致敏大鼠)相比,給予益生元飲食的大鼠具有口服耐受力,其腸道中厚壁菌門和變形菌門的相對豐度較低,軟壁菌門和藍藻菌門的相對豐度較高,進一步發現可可作為益生元可通過調節腸道菌群,增加腸系膜淋巴結中T細胞受體δ+(T-cell receptor δ+,TCRδ+)、CD103+和CD8+T細胞的比例,降低CD62L+CD4+和CD62L+CD8+細胞比例并調節Treg細胞功能,減少IgA產生,從而對過敏性反應起到抑制。然而,其他利用益生元干預食物過敏的人群實驗證據較為有限,一項系統性綜述指出,補充益生元可能預防兩歲以下的嬰兒發生濕疹,但是并不能起到預防食物過敏的作用[103]。總之,有關益生元在過敏性疾病(包括食物過敏)中的干預作用需要通過對更多的實驗結果進行評估。
表2總結了采用益生菌干預對食物過敏進行預防或治療的人群實驗研究。總體上,由于已有研究的異質性較高,尚沒有充足的證據表明在母親孕期、哺乳期或嬰童期使用益生菌、益生元或合生元可以起到預防食物過敏的作用。據此,國際上不同地區發布的食物過敏預防指南尚未針對益生菌、益生元或合生元的使用提出官方建議。例如EAACI頒布的食物過敏預防指南中指出,對于在懷孕、母乳喂養或嬰兒生命早期補充益生菌、益生元或合生元沒有明確的建議[104]。類似地,AAAAI、美國變態反應和臨床免疫學學會以及加拿大變態反應和臨床免疫學學會聯合發布的食物過敏預防共識,以及ASCIA發布的聲明中也指出,鑒于目前有限的臨床證據,尚不足以對使用益生元和益生菌預防食物過敏的可行性提出明確建議[105]。因此,為了進一步明確益生菌/益生元對腸道菌群和免疫系統的潛在影響,亟需更多關于食物過敏兒童腸道菌群失調的數據以及益生菌/益生元對腸道菌群平衡作用的系統研究,同時需要明確不同益生菌對食物過敏作用的機制,以期確定有效干預的最佳劑量、時機和藥物組合等。

表2 采用益生菌干預對食物過敏進行預防或治療的人群實驗研究Table 2 Human studies on the prevention or treatment of food allergy by probiotics intervention
5 肥胖與食物過敏的潛在關聯性
肥胖是由環境(如飲食)、遺傳和激素之間相互作用而導致的疾病。近幾十年間,受西方飲食和生活方式的影響,全球范圍內尤其是西方國家中飲食性肥胖的人群數量逐步攀升,鑒于部分地區食物過敏和肥胖癥患病率的同步增加,研究人員推測二者之間可能存在一定關聯[118]。支持這一假說的證據主要是基于:首先,肥胖常伴隨著慢性低密度炎癥狀態和免疫系統的非特異性激活,部分來自脂肪組織的促炎因子循環水平增加,這可能間接導致腸上皮屏障功能的損傷[119]。其次,在肥胖患者中瘦素的分泌量增加,瘦素可以促進巨噬細胞功能,抑制Treg細胞的增殖[120]。最后,肥胖引起的腸道微生物群失衡也可能會增加人體對食物致敏原的易感性[121]。
目前,針對肥胖與食物過敏之間的關聯研究大多為橫斷面研究。如Vieira等發現,18~41 歲女性肥胖患者的sIgE陽性率比正常人高出3 倍[122]。Visness等[123]也發現了類似結論,在2~19 歲兒童中,肥胖和超重兒童血清總IgE的水平顯著高于正常體重兒童,這表明肥胖和超重兒童更有可能出現食物過敏反應。此外,Luo Xiao等[124]發現,肥胖與成年人發生過敏性皮炎和鼻炎的比例呈顯著正相關。Irei等[125]調查了2027 名來自日本、越南和中國臺灣的9~13 歲兒童,發現在不同地區,肥胖與食物過敏之間存在的聯系不同。可見,肥胖在食物過敏中發揮作用的證據尚未得到一致結論。
在動物模型中,Gu Yanjun[126]、Gao Jingxin[127]等發現高脂飲食誘導的Balb/c肥胖小鼠較正常體重小鼠有更嚴重的食物過敏反應,進一步研究發現,肥胖可通過促進小鼠的腸道炎癥、破壞其腸道物理屏障進而導致食物過敏的發生。另外,對肥胖小鼠的后代進行腸通透性實驗,發現母體肥胖會導致子代小鼠的腸道通透性增加并誘發腸道炎性反應[128];此外也發現肥胖母鼠后代患哮喘和過敏性炎癥反應的可能性更高等[129]。
盡管肥胖與食物過敏的潛在關聯已在動物模型中得到初步驗證,但鑒于人群中較少的前瞻性研究和干預性研究,肥胖在食物過敏發生發展中的作用有待進一步探索,今后需開展觀察性隊列研究并系統評估肥胖和食物過敏臨床診斷的人體測量指標確定這兩種癥狀之間的因果關系。
6 結語
人體對食物建立口服耐受的過程會受基因、環境以及飲食等多方面因素的影響,涉及多種器官、組織、細胞及其產物的共同調控作用。在膳食方面,非致敏性物質的攝入會對食物過敏產生重要的影響,其中一部分是通過直接影響機體免疫應答發揮作用,另一部分則是通過調節腸道菌群的平衡進而對機體免疫系統的功能造成影響。這其中,就益生菌干預而言,雖然尚未有明確的食用建議,但已有較多的證據表明益生菌能夠通過改變腸道菌群的結構對食物過敏的預防產生有益效果;此外,西方飲食造成的肥胖可能通過多種路徑誘發食物過敏的發生發展,這在動物模型實驗中已經得到了初步驗證,但由于變量的多樣性,基于人群的前瞻性和干預性報道較少,考慮到當前西方飲食的普遍性以及肥胖與過敏患病率的同步增加,有關西方飲食誘導食物過敏發生的機制或可成為未來研究食物過敏危險因素的重要方向之一,此外,現代食品加工引入的外源性添加劑和內源性加工產物對機體免疫應答的潛在影響也值得關注。目前對于非致敏性食物成分在食物過敏進程中的作用研究仍處初級階段,未來這需要科研工作者大量的研究和驗證,以期達到通過飲食調理干預或預防食物過敏發生的目的。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
