基于牡丹澇害脅迫的轉(zhuǎn)錄組分析及SSR引物開發(fā)
2024-04-08 13:51:10劉慧春許雯婷周江華張加強史小華朱開元
浙江農(nóng)業(yè)學報 2024年3期
關(guān)鍵詞:差異
劉慧春,許雯婷,周江華,張加強,史小華,朱開元
(浙江省園林植物與花卉研究所,浙江 杭州 311251)
牡丹(PaeoniasuffruticosaL.)是原產(chǎn)于中國的著名花灌木,也是一種具有深厚文化內(nèi)涵的中國傳統(tǒng)花卉。牡丹原產(chǎn)于北方,主要種植于黃河流域中下游地區(qū),根系肥厚,對澇害較為敏感。長江以南地處長江中下游,地下水位偏高,雨季土壤水分過飽和,臺風、洪水等災害嚴重影響了長江以南牡丹的種植和應用。隨著人們欣賞水平的提高,迫切需要篩選出適合江南地區(qū)廣泛種植的牡丹品種。耐澇品種的選育一直是牡丹南下栽培的重點和難點。目前,江南牡丹的種植主要采用塑料薄膜防雨,限制了植物的自然光照,從而影響了牡丹的開花質(zhì)量。如何提高牡丹的耐澇性已成為牡丹生產(chǎn)中亟待解決的問題。因此,深入研究牡丹在澇害脅迫下的響應機制,對牡丹生產(chǎn)具有重要的現(xiàn)實意義和應用價值。
在過去的幾年里,轉(zhuǎn)錄組測序技術(shù)已被廣泛應用于功能基因組研究。隨著新一代測序的快速發(fā)展,RNA-Seq已成為一種有效的轉(zhuǎn)錄組數(shù)據(jù)分析方法。此外,它還被用來闡明對各種非生物脅迫的反應機制,包括干旱、低溫、鹽堿和澇害[1-7]。到目前為止,很少有關(guān)于牡丹澇害脅迫相關(guān)分子機制的報道。僅見報道的有,通過全長轉(zhuǎn)錄組測序和第二代轉(zhuǎn)錄組測序的結(jié)合,初步探討了鳳丹牡丹對淹水脅迫反應的分子機制[8]。因此,對牡丹響應澇害脅迫的綜合轉(zhuǎn)錄組學分析還有待進一步研究和提升。
隨著高通量測序技術(shù)的快速發(fā)展,進一步延伸出利用轉(zhuǎn)錄組序列開發(fā)SSR標記,并已被廣泛應用于群體遺傳學、遺傳連鎖圖的構(gòu)建,種質(zhì)與標記輔助選擇育種之間的遺傳關(guān)系分析[9-12]。前人也曾通過RNA-Seq開發(fā)出牡丹的一些SSR標記[13]。例如,前人在滇牡丹(P.delavayi)中開發(fā)了10個SSR標記[14],在紫斑牡丹(P.rockii)中開發(fā)了11個與農(nóng)藝性狀相關(guān)的SSR標記、44個與種子性狀相關(guān)的SSR標記[15-16],在牡丹(P.suffruticosa)中開發(fā)了12個與花色形成有關(guān)的SSR標記[17]。然而,與玉米(Zeamays)和大豆(Glycinemax)等作物相比,牡丹中目前的SSR標記數(shù)量仍然有限,而且關(guān)于牡丹澇害脅迫相關(guān)基因的SSR分子標記的報道很少。
在本研究中,通過RNA-Seq對耐澇牡丹根的轉(zhuǎn)錄組進行了分析,以闡明其對澇害脅迫反應的分子機制。基于構(gòu)建的高通量RNA-Seq數(shù)據(jù)庫,在牡丹中開發(fā)了調(diào)控澇害脅迫性狀的SSR標記。該研究結(jié)果對牡丹進行了大規(guī)模轉(zhuǎn)錄組注釋,將為未來耐澇牡丹品種的育種研究提供寶貴的基因組資源,并為進一步研究其遺傳多樣性、功能基因的發(fā)現(xiàn)和定位、種質(zhì)資源鑒定和標記輔助選擇提供科學依據(jù)。
1 材料與方法
1.1 植物材料及處理
實驗材料為3年生鳳丹牡丹的實生苗,由浙江省園林植物與花卉研究所提供。選擇50株在人工澇害脅迫處理下能夠存活下來的植株,在含有泥炭、珍珠巖和園土(體積比2∶1∶1)的培養(yǎng)基質(zhì)中培養(yǎng)。培養(yǎng)條件為:光照14 h,溫度25 ℃,光照強度為160 μmol·m-2·s-1光子,相對濕度為50%~60%。將其中25株幼苗作為處理組,放置在土壤表面上方含有3 cm水深的泡沫箱中。將另外25株未經(jīng)處理的幼苗置于正常培養(yǎng)條件下作為對照。處理48 h后,從兩個處理(對照和處理的植株)中采集幼根,然后在液氮中快速冷凍并在-80 ℃下儲存?zhèn)溆谩?/p>
1.2 RNA提取
將對照樣品(CK1、CK2和CK3)和澇害處理的樣品(W1、W2、W3)的根作為3個生物學重復。根據(jù)產(chǎn)品說明書,使用植物總RNA純化試劑盒(GeneMark TR02)分離總RNA。通過變性瓊脂糖凝膠電泳檢測RNA質(zhì)量,并使用NanoDrop 2000分光光度計(Thermo Scientific,USA)測定RNA濃度。
1.3 RNA序列文庫的構(gòu)建
從經(jīng)過正常條件(對照樣品CK1、CK2、CK3)和澇害條件(澇害處理的樣品W1、W2、W3)的牡丹根中分離的6個高質(zhì)量RNA樣品,分別用于cDNA文庫的構(gòu)建,然后將文庫加入高通量測序平臺(Illumina HiSeq 2500,USA)進行測序。cDNA文庫的構(gòu)建方法和Illumina深度測序的過程參考Li等[18]的方法。RNA-Seq讀數(shù)通過Illumina數(shù)據(jù)處理軟件(Version 1.8)生成。
1.4 數(shù)據(jù)組裝、讀取和基因注釋
按照Li等[18]描述的方法進行轉(zhuǎn)錄組數(shù)據(jù)組裝,采用NR、Swissport、KOG和KEGG數(shù)據(jù)庫對基因功能進行注釋,這些數(shù)據(jù)庫通過BLASTA進行比對,截斷E值≤1.0-5。
1.5 差異表達基因分析
基于上述獲得的RNA-Seq數(shù)據(jù),采用在線工具degust (http://vicbioinformatics.com/degust/) 生成主成分分析(PCA)圖。通過軟件包RPKM評估和量化基因表達水平,并使用Cufflinks軟件的Cuffdiff程序(v2.0.2,http://cole-trapnell-lab.github.io/cufflinks/),使用FDR調(diào)整基因的多次測試和分析中的P閾值。FDR調(diào)整后的P≤0.05和log2R倍數(shù)變化的絕對值≥1(倍數(shù)變化≥2)被設定為顯著差異表達的閾值。基于Wallenius非中心超基因組分布的GOseq R軟件包對DEG進行GO富集分析,該軟件包可以調(diào)整DEG中的基因長度偏差。采用KOBAS軟件測試KEGG途徑中DEG的統(tǒng)計富集。
1.6 qRT-PCR分析
隨機選擇14個來自所有DEG的單基因,使用qPCR驗證基于RNA-Seq的表達水平分析的可靠性和準確性。利用Oligo 7.57軟件設計單基因的引物,按照Li等[18]描述的方法進行qRT-PCR驗證。PCR反應體系為20 μL,包括:1 μL cDNA模板、1 μL引物混合物、10 μL 2×SYBR Green qPCR混合物和8 μL ddH2O。用Mastercycler?ep realplex4進行qPCR反應,反應程序如下:95 ℃預變性3 min;95 ℃變性5 s,60 ℃退火10 s,40個循環(huán);最后95 ℃延伸15 s。選擇18S rRNA作為參考基因,并使用2-ΔΔCT方法對基因的相對表達水平進行測算。
1.7 SSR鑒定、引物設計和PCR擴增
根據(jù)RNA-Seq數(shù)據(jù),使用SSRIT軟件(http://archive/gramene.org/db/markers/ssrtool)鑒定SSR位點[17]。已鑒定的基因位點包含二、三、四、五和六個核苷酸,最小重復數(shù)分別為4、3、3、4和2。使用Primer Premier 5.0從差異基因表達數(shù)據(jù)中選擇并設計這些SSR基因位點。SSR引物的參數(shù)設定為:GC含量40%~60%,引物長度20~25 bp,預期片段大小100~300 bp,退火溫度55~65 ℃,上下游引物差異在2 ℃以內(nèi)。
PCR擴增的反應體系為20 μL,包括:dd H2O 14.8 μL,dNTP 0.4 μL,緩沖液2 μL,20 μmol·L-1上下游引物各0.3 μL,DNA模板2 μL和Taq0.2 μL。PCR擴增的反應程序為:94 ℃預變性5 min;94 ℃變性30 s,54 ℃復性35 s,72 ℃延伸40 s,共35個循環(huán);最后,72 ℃延伸3 min。通過1.5%瓊脂糖電泳檢測PCR擴增產(chǎn)物,點樣量為3 μL。將3 μL可產(chǎn)生特定條帶的PCR擴增產(chǎn)物與5 μL加載緩沖液混合。離心后,在94 ℃變性5 min,在冰箱中冷凍1 min,然后采用6%變性聚丙烯酰胺凝膠進行垂直電泳分析,銀染后觀察結(jié)果。能擴增出清晰條帶的引物被視為有效引物,不能擴增出清晰條帶或擴增條帶不在范圍內(nèi)的引物被認為是無效引物。
2 結(jié)果與分析
2.1 轉(zhuǎn)錄組測序和組裝
為了獲得牡丹耐澇性的總體情況,使用高通量測序平臺構(gòu)建并測序了來自CK1、CK2、CK3(處理)和W1、W2、W3(對照)的6個cDNA文庫。總共產(chǎn)生了444 382 372個Illumina原始讀數(shù),其長度范圍從低于50 bp到151 bp。在去除接頭序列和低質(zhì)量讀數(shù)后,獲得了438 315 223個干凈讀數(shù)。每個樣本中的表達基因數(shù)量在57 024到61 583之間。從組裝轉(zhuǎn)錄物中共獲得74 756個參考unigene,平均長度為743 bp,長度范圍為201~12 252 bp。基于長度的unigene分布如圖1所示。
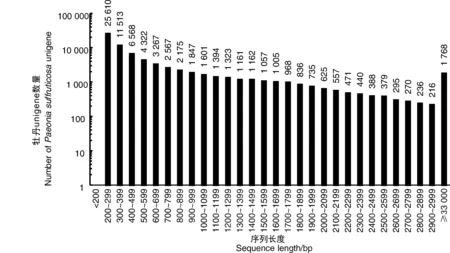
圖1 牡丹轉(zhuǎn)錄組unigene的長度分布和豐度Fig.1 Length distribution and abundance of unigenes from the P. suffruticosa transcriptome
2.2 功能注釋
利用4個公共數(shù)據(jù)庫(NR、Swissprot、KOG和KEGG)對unigene進行注釋。注釋結(jié)果顯示,在NR數(shù)據(jù)庫中有33 105個unigene(44.28%)匹配,在Swissprot數(shù)據(jù)庫中有22 715個(30.38%)匹配,KOG數(shù)據(jù)庫中有19 613個(26.24%)匹配,KEGG數(shù)據(jù)庫中有12 763個(17.07%)匹配。共有33 278個unigene(44.51%)與4個數(shù)據(jù)庫中的一個或多個數(shù)據(jù)庫匹配,其中9 761個unigene在所有4個數(shù)據(jù)庫中匹配(13.06%)。同時,這些結(jié)果表明,Illumina測序測試了牡丹中廣泛表達的基因,其中6 106、1 986、1 964和1 921個分別與葡萄、核桃、可可和土瓶草的表達基因高度相似(圖2)。
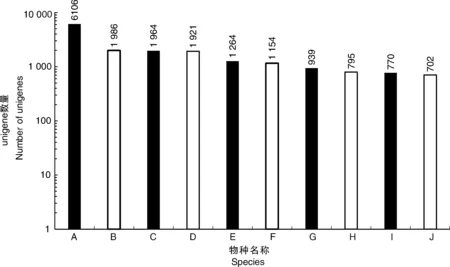
A,葡萄;B,核桃;C,可可;D,土瓶草; E,蓮藕;F,酸棗;G,梅花;H,麻瘋樹;I,柑橘; J,木豆。A, Vitis vinifera; B, Juglans regia; C, Theobroma cacao; D, Cephalotus follicularis; E, Nelumbo nucifera; F, Ziziphus jujuba; G, Prunus mume; H, Jatropha curcas; I, Citrus sinensis; J, Cajanus cajan.圖2 unigene BLASTX比對結(jié)果的物種分布圖Fig.2 Species distribution of unigene BLASTX results
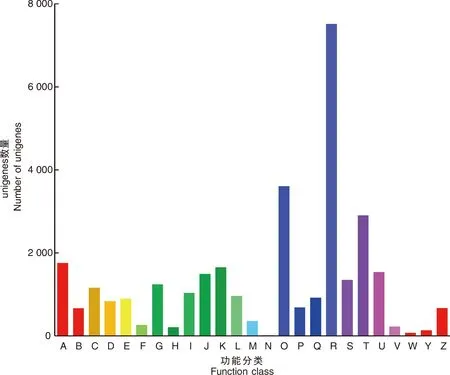
利用GO分析,將預測的牡丹unigene的功能分為3類:生物過程、細胞成分和分子功能。每個類別分別被進一步聚類為17、15和11個子類別,共包含43個子類別(圖3)。對于生物過程類別,“細胞過程”(2 687)、“代謝過程”(3 297)和“單體過程”(2 163)是前3個富集組。細胞成分類別主要包含涉及“細胞”(1 588)、“細胞部分”(1 584)和“細胞器”(1 132)的基因。在分子功能類別中,“結(jié)合”(2 309)、“催化活性”(3 454)和“轉(zhuǎn)運蛋白活性”(330)子類別的代表性很高。
a-q,生物過程(a,再生產(chǎn);b,免疫系統(tǒng)過程;c,代謝過程;d,細胞過程;e,再生過程;f,生物黏附;g,信號傳導;h,多細胞生物過程;i,發(fā)展過程;j,增長;k,單體過程;l,韻律過程;m,對刺激的反應;n,本地化;o,多生物過程;p,生物調(diào)控;q,細胞成分組織或生物發(fā)生); r-z,A-F,細胞成分 (r,細胞外區(qū)域;s,細胞;t,膜;u,病毒粒子;v,細胞連接;w,細胞外基質(zhì);x,膜封閉管腔;y,大分子復合物;z,細胞器;A,細胞外基質(zhì)成分;B,細胞器部分;C,病毒粒子部分;D,膜部分;E,細胞部分;F,超分子纖維); G-Q,分子功能 (G,轉(zhuǎn)錄因子活性、蛋白質(zhì)結(jié)合;H,核酸結(jié)合轉(zhuǎn)錄因子活性;I,催化活性;J,信號轉(zhuǎn)換器活動;K,結(jié)構(gòu)分子活性;L,轉(zhuǎn)運蛋白活性;M,綁定;N,電子載體活性;O,抗氧化活性;P,分子轉(zhuǎn)導活性;Q,分子功能調(diào)節(jié)劑)。a-q, Biological process (a, reproduction; b, immune system process; c, metabolic process; d, cellular process; e, reproductive process; f, biological adhesion; g, signaling; h, multicellular organismal process; I, developmental process; j, growth; k, single-organism process; l, rhythmic process; m, response to stimulus; n, localization; o, multi-organism process; p, biological regulation; q, cellular component organization or biogenesis) ; r-z, A-F, Cellular component (r, extracellular region; s, cell; t, membrane; u, virion; v, cell junction; w, extracellular matrix; x, membrane-enclosed lumen; y, macromolecular complex; z, organelle; A, extracellular matrix component; B, organelle part; C, virion part; D, membrane part; E, cell part; F, supramolecular fiber) ; G-Q, Molecular function (G, transcription factor activity, protein binding; H, nucleic acid binding transcription factor activity; I, catalytic activity; J, signal transducer activity; K, structural molecule activity; L, transporter activity; M, binding; N, electron carrier activity; O, antioxidant activity; P, molecular transducer activity; Q, molecular function regulator).圖3 澇害脅迫下牡丹差異表達基因的GO分類圖Fig.3 GO classification map of differentially expressed genes during waterlogging stress in P. suffruticosa
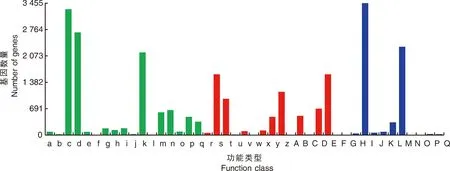
此外,共有32 059個unigene被分配到KOG分類中,并分為25個特定類別(圖4)。最大的一組屬于“僅一般功能預測”(7 512),其次是“翻譯后修飾、蛋白質(zhì)周轉(zhuǎn)、伴侶蛋白”(3 604)和“信號轉(zhuǎn)導機制”(2 901)。只有少數(shù)unigene被分為“細胞外結(jié)構(gòu)”(68)和“細胞運動性”(13)。
A,RNA加工和修飾;B,染色質(zhì)結(jié)構(gòu)和動力學;C,能源產(chǎn)生與轉(zhuǎn)換;D,細胞周期控制、細胞分裂、染色體分割;E,氨基酸運輸和代謝;F,核苷酸運輸和代謝;G,碳水化合物運輸和代謝;H,輔酶運輸和代謝;I,脂質(zhì)運輸和代謝;J,翻譯、核糖體結(jié)構(gòu)與生物發(fā)生;K,轉(zhuǎn)錄;L,復制、重組和修復;M,細胞壁/膜/包膜生物發(fā)生;N,細胞運動性;O,翻譯后修飾、蛋白質(zhì)轉(zhuǎn)運蛋白、伴侶蛋白;P,無機離子運輸和代謝;Q,次生代謝產(chǎn)物的生物合成、運輸和分解代謝;R,僅通用功能預測;S,功能未知;T,信號轉(zhuǎn)導機制;U,細胞內(nèi)運輸、分泌和囊泡運輸;V,防御機制;W,細胞外結(jié)構(gòu);Y,核結(jié)構(gòu);Z,細胞骨架。A, RNA processing and modification; B, Chromatin structure and dynamics; C, Energy production and conversion; D, Cell cycle control, cell division, chromosome partitioning; E, Amino acid transport and metabolism; F, Nucleotide transport and metabolism; G, Carbohydrate transport and metabolism; H, Coenzyme transport and metabolism; I, Lipid transport and metabolism; J, Translation, ribosomal structure and biogenesis; K, Transcription; L, Replication, recombination and repair; M, Cell wall/membrane/envelope biogenesis; N, Cell motility; O, Posttranslational modification, protein turnover, chaperones; P, Inorganic ion transport and metabolism; Q, Secondary metabolites biosynthesis, transport and catabolism; R, General function prediction only; S, Function unknown; T, Signal transduction mechanisms; U, Intracellular trafficking, secretion, and vesicular transport; V, Defense mechanisms; W, Extracellular structures; Y, Nuclear structure; Z, Cytoskeletion.圖4 澇害脅迫下牡丹差異表達基因的KOG分類和分析Fig.4 KOG classification and analysis of differentially expressed genes during waterlogging stress in P. suffruticosa
KEGG系統(tǒng)還對與澇害脅迫相關(guān)的unigene生物途徑進行了注釋。總共發(fā)現(xiàn)13 881個unigene參與了135個KEGG途徑。涉及最多unigene的途徑是“代謝途徑”(2 733,39.22%),其次是“次級代謝產(chǎn)物的生物合成”(1 453,20.85%)和“抗生素的生物合成”(669,9.6%)。
2.3 澇害脅迫下的差異表達基因
為了可視化6個RNA-Seq樣本之間的差異表達基因,進行了PCA繪圖。該圖顯示,3個對照樣品(CK1、CK2、CK3)和3個澇害處理的樣品(W1、W2、W3)分別沿維度1和維度2清晰區(qū)分(圖5)。為了鑒定在淹水脅迫下積累的牡丹的差異表達轉(zhuǎn)錄物,使用RPKM方法測量基因表達水平。使用FDR≤0.05和log2R≥1(R≥2)的值作為閾值來估計差異基因表達的顯著性。在W組和CK組之間共檢測到780個表達顯著改變的基因,包括155個上調(diào)基因和625個下調(diào)基因(圖6)。此外,在W和CK的文庫中分別有71 187和70 697個轉(zhuǎn)錄本特異性表達。基于這些結(jié)果,我們發(fā)現(xiàn)牡丹的大多數(shù)基因在對淹水脅迫的反應中都有差異表達。在這些差異表達基因中,大量基因可能在淹水脅迫中發(fā)揮關(guān)鍵作用。
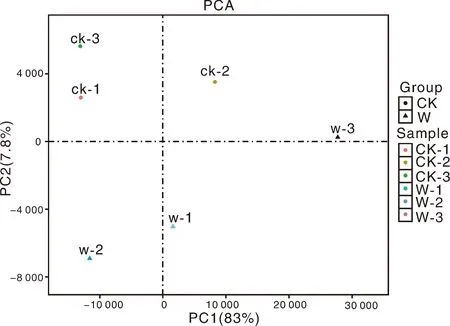
圖5 六個RNA-Seq樣本的主成分分析(PCA)圖Fig.5 Principal component analysis (PCA) plot of 6 RNA-Seq samples
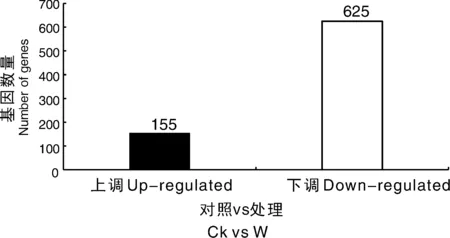
圖6 澇漬脅迫下牡丹差異表達基因統(tǒng)計圖Fig.6 Changes in gene expression profiles between the waterlogging stress and control libraries of P. suffruticosa
2.4 差異表達基因的功能分析
進行GO富集分析,對澇害處理組和對照組之間差異表達基因的推定功能進行分類。如上所述,差異表達的基因被分為3大類,包括生物進程、分子功能和細胞成分。在這3個類別下,前30個富集的功能組如表1所示,具體包括核小體組織、蛋白質(zhì)-DNA復合物亞單位組織、染色體組織、蛋白質(zhì)復合物亞單位組織等10個生物進程類功能組,蛋白質(zhì)二聚化活性、蛋白質(zhì)結(jié)合、核酸結(jié)合、雜環(huán)化合物結(jié)合等10個分子功能類功能組,以及無膜細胞器、細胞內(nèi)無膜細胞器、核腔、細胞器腔等10個細胞成分類的功能組。

表1 GO類別下最富集的30個功能組Table 1 Top 30 most enriched functional groups under GO categories
為了進一步闡明差異表達的unigene在響應澇害脅迫中的功能,將差異表達的轉(zhuǎn)錄物與KEGG數(shù)據(jù)庫類別進行匹配。結(jié)果顯示,共有74條途徑與澇害脅迫顯著相關(guān)。前11個富集的功能途徑如表2所示。在這11條KEGG途徑中,3條最顯著富集的途徑分別是DNA復制、核糖體和嘧啶代謝。此外,還發(fā)現(xiàn)了與吞噬體、堿基切除修復、內(nèi)質(zhì)網(wǎng)蛋白質(zhì)加工、嘌呤代謝等相關(guān)的其他重要途徑,推測是牡丹響應澇害脅迫的主要途徑。

表2 最富集的11個KEGG途徑Table 2 Top 11 most enriched KEGG pathways
2.5 響應澇害脅迫的轉(zhuǎn)錄因子的鑒定
在澇害處理和對照條件下鑒定出多種差異表達的轉(zhuǎn)錄因子。在57個轉(zhuǎn)錄因子(TF)家族中,前10個TF家族如圖7所示。在這10個TF家族中,大多數(shù)TF屬于bHLH(92個unigene)、ERF(74個unigene)和MYB_related(65個unigene)。bHLH家族是在澇害脅迫條件下顯著差異表達數(shù)量最多的一類轉(zhuǎn)錄因子,其他7個轉(zhuǎn)錄因子家族分別屬于C2H2(62個unigenes)、FAR1(62個unigenes)、MYB(53個unigenes)、bZIP(51個unigenes)、C3H(50個unigenes)、NAC(44個unigenes)和WRKY(40個unigenes)家族。有25個、14個和12個差異表達的unigenes分別被分為HD-ZIP、ARF和AP2家族。

圖7 前10個轉(zhuǎn)錄因子家族的基因數(shù)量Fig.7 Gene number of top 10 transcription factor families
2.6 qRT-PCR驗證
為了驗證RNA-Seq數(shù)據(jù)的準確性和可靠性,從差異表達基因中隨機選擇14個unigenes,使用特異性引物進行qRT-PCR分析。在14個被檢測的unigenes中,11個基因的qRT-PCR表達模式與RNA-Seq數(shù)據(jù)(圖8)顯示出高度相似性。這一結(jié)果反映了基于RNA-Seq的差異表達分析是可靠和準確的。此外,使用倍數(shù)變化測量來計算RNA-Seq和qRT-PCR數(shù)據(jù)之間的相關(guān)性。隨后,在兩個數(shù)據(jù)組之間發(fā)現(xiàn)了良好的一致性(R2=0.933 8)(圖8),證實了RNA-Seq方法獲得的結(jié)果是有效和可靠的。
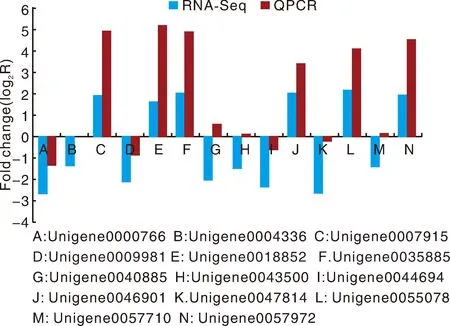
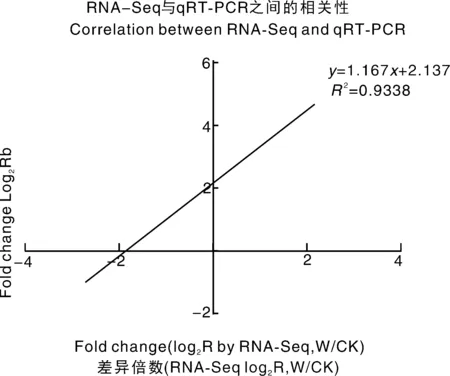
圖8 qRT-PCR對基因相對表達水平的驗證Fig.8 Verification of the relative expression levels of genes by qRT-PCR
2.7 SSR標記的分布和特征
從牡丹RNA-Seq數(shù)據(jù)中,在74 756個unigenes中發(fā)現(xiàn)了4 596個unigenes,包含5 204個SSR位點,頻率為6.15%(包含SSR位點的unigene占總unigene的比例)。每個unigene至少有1個SSR位點,302個unigenes包含多個SSR位點。在5 204個SSR基因位點中,二核苷酸重復序列(3 025個,58.13%)占優(yōu)勢,其次是三核苷酸(1 411個,27.11%)、四核苷酸(377個,7.24%)、五核苷酸(128個,2.46%)和六核苷酸重復(263個,5.05%)(表3)。
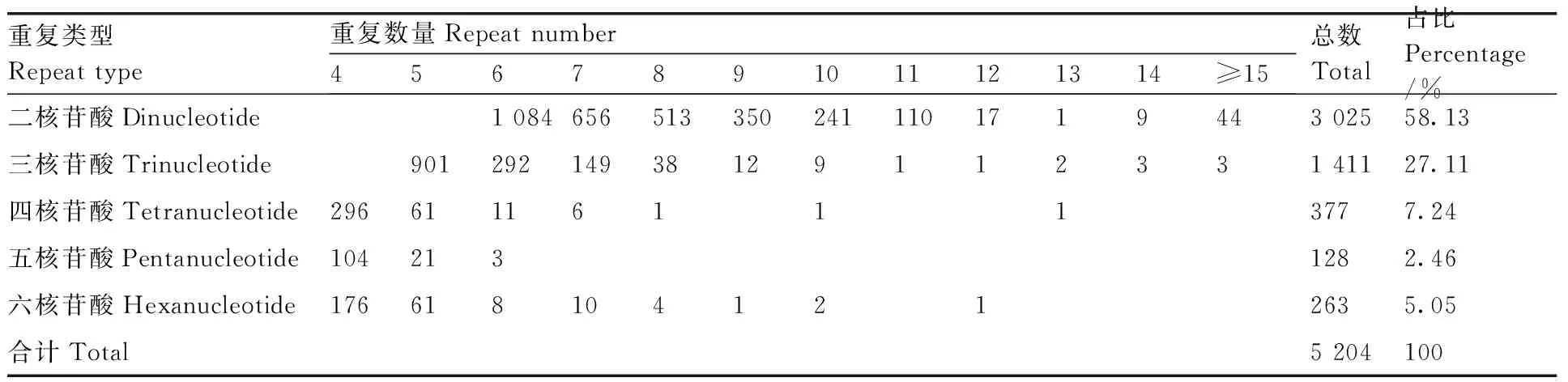
表3 牡丹耐澇相關(guān)unigene中SSR的類型、數(shù)量和分布頻率Table 3 Type, number and distribution frequency of SSRs in Paeonia suffruticosa waterlogging related unigenes
對轉(zhuǎn)錄組中SSR的核苷酸類型進行分析,結(jié)果表明,在5 204個SSR位點中有212個重復基序。在二核苷酸重復基序中,最明顯的顯性基序是AG/CT(2 153,41.4%),其次是AT/AT(513,9.9%)和AC/CT(352,6.8%)。在三核苷酸重復基序中,最常見的基序是AAG/CTT(362,7%)、ACC/GGT(277,5.3%)和ATC/ATG(196,3.8%)。重復基序AAAT/ATT(171,3.3%)僅存在于四核苷酸重復基序中。然而,在其他類型中沒有明顯的顯性基序(表4、圖9)。
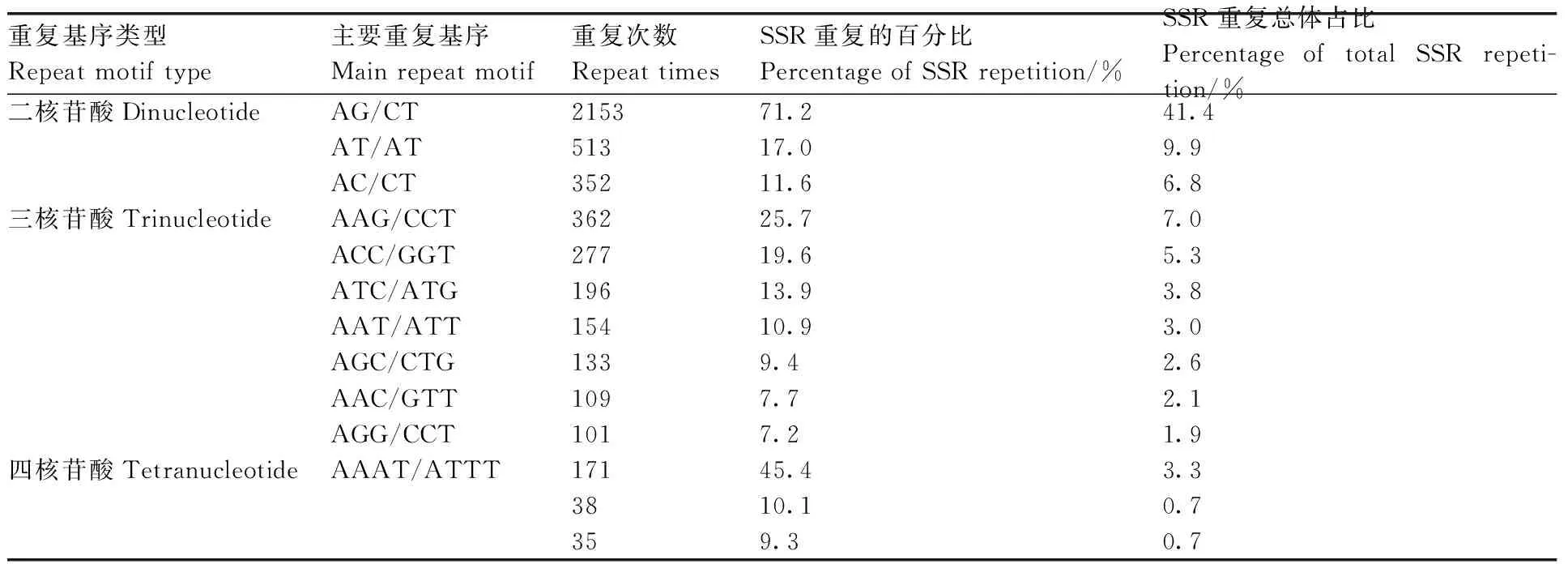
表4 牡丹澇害轉(zhuǎn)錄組SSR重復基序的類型和頻率分布Table 4 Types and frequency distribution of SSR repeat motifs in the waterlogging transcriptome of Paeonia suffruticosa
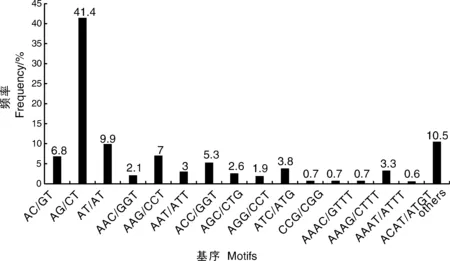
圖9 二核苷酸、三核苷酸、四核苷酸和其他類型重復基序的頻率分布Fig.9 Frequency distribution of di-, tri-, tetra-nucleotide and other type repeat motifs
2.8 澇害脅迫下SSR標記開發(fā)
本研究以與澇害脅迫調(diào)控相關(guān)的基因為候選基因,在葉片中發(fā)現(xiàn)了780個與澇害脅迫反應相關(guān)的差異表達基因。通過與SSR基因位點的比較,從差異表達的基因中發(fā)現(xiàn)55個含有SSR基因位點。設計了110對引物,篩選得到45對引物(表5)。以牡丹嫩葉DNA為模板,初步測試了引物的有效性,并對12對引物進行了二次篩選,在9個品種(紅芙蓉、玉樓春、大富貴、一品紅、島錦、島輝、鳳丹和2個新品種)中擴增出清晰穩(wěn)定的靶帶(圖10),平均多態(tài)性為83%。
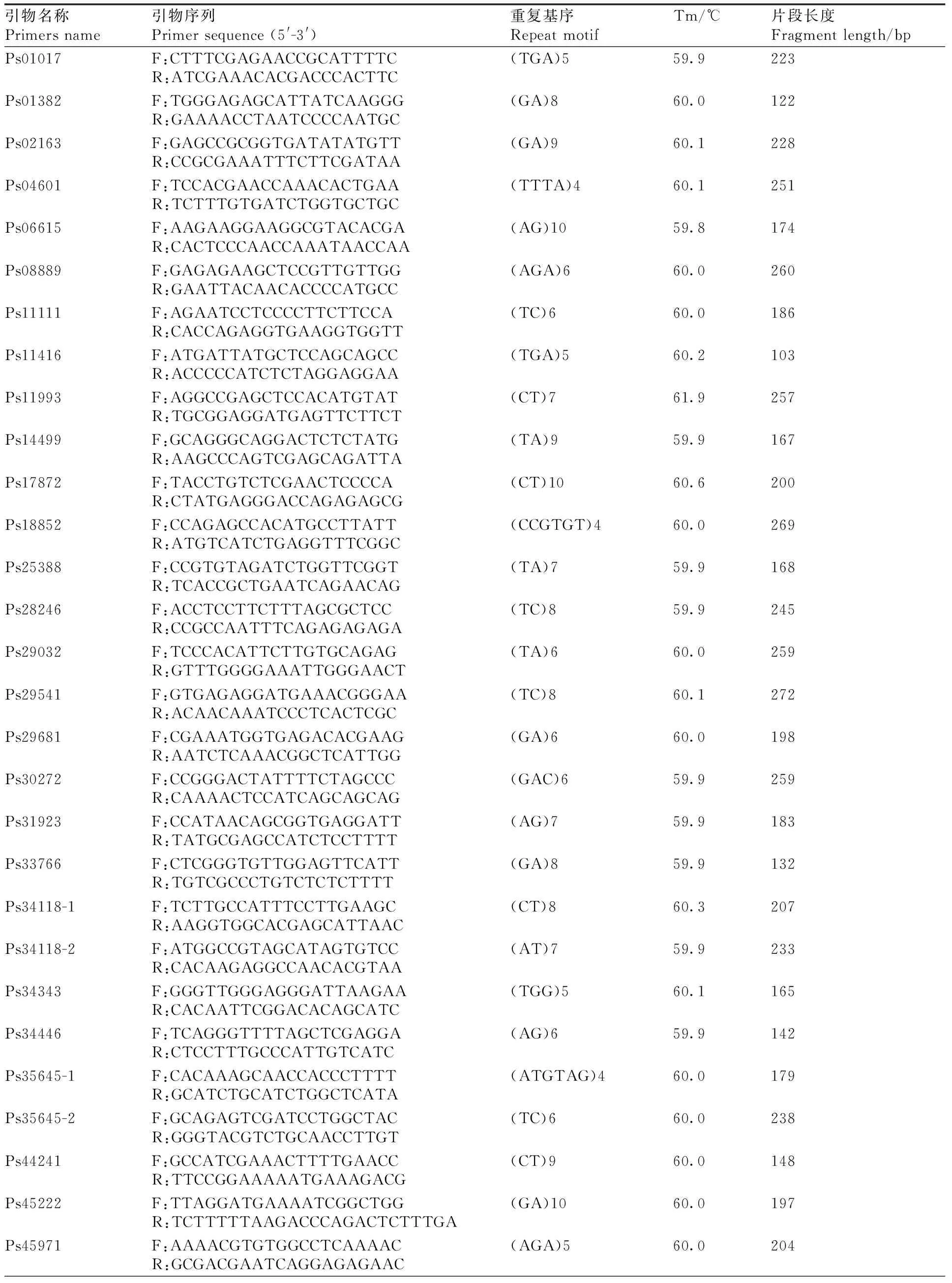
表5 牡丹澇害脅迫相關(guān)的SSR引物開發(fā)Table 5 SSR primers developed from genes related to waterlogging stress in Paeonia suffruticosa
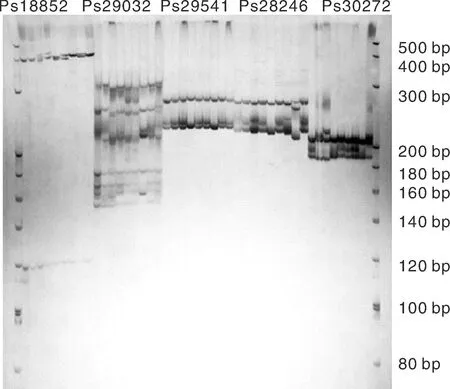
圖10 SSR引物Ps18852, Ps28246, Ps29032, Ps29541和 Ps30272在9個牡丹品種中的擴增結(jié)果Fig.10 Polymorphism of Ps18852, Ps28246, Ps29032, Ps29541, and Ps30272 of SSR primers in 9 samples of P. suffruticosa
9個牡丹品種基因組DNA中12對引物的擴增帶均具有多態(tài)性(表6),其中5對引物占100%的多態(tài)性,4對引物占80%以上的多態(tài)性。觀察到的等位基因數(shù)(Na)在1.333 3~2.000 0,平均為1.830 7。有效等位基因數(shù)(Ne)在1.060 4~1.692 6,平均1.388 4。Nei基因多樣性的變異范圍為0.148 1~0.397 2,平均為0.256 1。香農(nóng)信息指數(shù)在0.109 6~0.5581 8,平均值為0.372 0。這些結(jié)果表明,本研究開發(fā)的12對EST-SSR引物都是有效的通用引物,可用于牡丹品種的遺傳多樣性分析和品種鑒定。

表6 十二個SSR引物在9個牡丹品種中的遺傳多樣性指數(shù)Table 6 Genetic diversity index for 12 SSR primers among 9 Paeonia suffruticosa cultivars
3 討論
耐澇性是一個極其復雜的特征,涉及許多代謝途徑和大量基因。在差異表達的基因中,一些具有功能注釋的基因可能參與了耐澇性的調(diào)節(jié),其中一些是未報道的基因。它們差異表達的原因和作用機制尚不清楚,需要進一步研究。對這些基因的進一步研究將有助于我們更好地了解牡丹耐澇性的調(diào)控機制。
本研究篩選出780個差異表達基因,其中37個差異表達最明顯的基因與耐澇性相關(guān),包含10個上調(diào)基因和27個下調(diào)基因。在37個差異表達基因中,有6個基因含有4個Hsp70和2個Hsp80,它們與熱休克同源蛋白(Hsp)有關(guān)。Hsp70是一種熱休克蛋白,長期以來一直被認為是最保守的蛋白家族之一。它可以對外部環(huán)境刺激作出反應,提高身體適應不利環(huán)境的能力。Hsp70蛋白廣泛存在于細菌、植物和人類中[19]。在植物中,它們在應對非生物脅迫方面發(fā)揮著至關(guān)重要的作用,如熱、冷、干旱和鹽度等環(huán)境因素[20]。然而,Hsp對澇害脅迫的反應尚未報道。
通過分析,我們還發(fā)現(xiàn)了一些與轉(zhuǎn)錄因子相關(guān)的基因,如AP2/ERF、MYB和bZIP轉(zhuǎn)錄因子。這些轉(zhuǎn)錄因子均上調(diào),并對其靶基因的表達產(chǎn)生進一步影響,從而決定牡丹的耐澇性。AP2/ERF基因家族成員被報道為與植物澇害脅迫相關(guān)的重要基因[21-23],ERFⅦ亞家族N末端的一個保守基序(NH2-MCGGAI/L)被證實與澇害脅迫適應有關(guān)[24]。MYB轉(zhuǎn)錄因子調(diào)節(jié)包括澇害脅迫在內(nèi)的大量應激反應基因的表達譜[25-26]。例如,對棉花電子表達譜的分析表明,GhMYB基因在棉鈴中特異性表達,并由根和葉中的耐澇脅迫誘導[27]。bZIP轉(zhuǎn)錄因子是近年來研究最多的轉(zhuǎn)錄因子家族之一。bZIP家族在包括擬南芥、煙草、水稻、小麥等在內(nèi)的大量植物中被鑒定,其表達和功能被證明與耐澇性有關(guān)[25,28]。我們目前的研究結(jié)果進一步支持AP2/ERF、MYB和bZIP轉(zhuǎn)錄因子可能在牡丹的耐澇反應中發(fā)揮作用,并對許多耐澇基因的激活起重要作用,從而增強耐澇性。因此,提高一個關(guān)鍵轉(zhuǎn)錄因子的表達可能會提高牡丹的耐澇性。
還有一些其他與脫氫酶相關(guān)的蛋白質(zhì)基因,如MDH、ADH、GDH等,以及一些編碼滲透調(diào)節(jié)物質(zhì)的相關(guān)基因如APX,可能在牡丹耐澇性的調(diào)節(jié)中發(fā)揮重要作用。
SSR標記作為一種能夠區(qū)分顯性和隱性等位基因的共顯性標記,已被廣泛用于研究種質(zhì)之間的遺傳關(guān)系和遺傳多樣性,以及連鎖定位和基因定位[29-30]。新一代高通量測序技術(shù)(RNA-Seq)的快速發(fā)展為SSR標記的開發(fā)提供了一種快速有效的方法[31]。與普通的EST-SSR標記物相比,在基因或候選基因中開發(fā)的SSR可以更好地適應特定和多樣化的市場需求。本研究在牡丹轉(zhuǎn)錄組數(shù)據(jù)庫中檢索了74 756個單基因拼接的SSR,獲得了5 204個SSR位點,頻率為6.56%,高于馬尾松的4.32%[32]、菊花的2.84%[33],但低于三角梅的44.91%[34],24.46%的杜鵑[35]和15.2%的鳳丹[14]。這表明轉(zhuǎn)錄組SSR中包含的信息不僅在物種之間不同,而且在品種之間也不同,或者SSR分析軟件設定的標準不同。
多態(tài)性SSR分子標記的開發(fā)是物種重要性狀分子標記輔助育種的基礎(chǔ)和前提。在本研究中,獲得了12個多態(tài)性引物,占有效引物的26.67%,低于牡丹(62%)[16],但高于牡丹(19.61%)[14]。它接近牡丹的27%[36]、25%[13]和24%[17]。大量研究表明,SSR基因座中重復序列的長度越長,SSR標記的多態(tài)性程度通常越高[37-38]。此外,多態(tài)性水平也可能受到遺傳基礎(chǔ)和實驗材料樣本量的影響[38]。例如,在24個芝麻樣品中,多態(tài)性SSR的百分比為11.59%。樣本量增加到25個后,多態(tài)性SSR的百分比上升到60.50%[39]。據(jù)預測,SSR引物多態(tài)性的比例也會隨著樣本遺傳背景和樣本量的增加而增加。
在本研究中,我們利用牡丹的轉(zhuǎn)錄組信息開發(fā)了多個與澇害脅迫基因相關(guān)的SSR分子標記,并驗證了這些標記的有效性和普遍性。研究結(jié)果進一步豐富了牡丹的特異性分子標記數(shù)量,為后續(xù)牡丹分子標記輔助育種及相關(guān)分子生物學研究奠定了基礎(chǔ)。接下來,我們將進一步驗證這些SSR標記與耐澇性狀之間的關(guān)系,為牡丹耐澇種質(zhì)鑒定、遺傳多樣性分析和分子標記輔助育種提供科學依據(jù)。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(shù)(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現(xiàn)代語文(2016年21期)2016-05-25 13:13:50
