
HCMV-IE2對果糖誘導的UL122轉基因模型小鼠肝臟脂肪變性的影響
2024-04-29 00:44:03周曉瓊張現娟王斌
精準醫學雜志 2024年1期
周曉瓊 張現娟 王斌

[摘要]目的探討人巨細胞病毒即刻早期蛋白2(HCMV-IE2)對果糖誘導的UL122轉基因模型小鼠肝臟脂肪變性的影響。方法構建UL122轉基因小鼠模型作為實驗組,以同齡野生型小鼠作為對照組,以60%的高果糖飼料飼養8周,監測小鼠體質量、血糖的變化,并行葡萄糖耐量實驗和胰島素耐量實驗后處死小鼠,測定小鼠血清中丙氨酸氨基轉移酶(ALT)和天冬氨酸氨基轉移酶(AST)水平以及肝臟組織中三酰甘油(TG)和總膽固醇(TC)水平;取小鼠肝臟組織行HE染色和油紅O染色進行組織病理學觀察;流式細胞術檢測肝臟組織中巨噬細胞極化情況;RT-qPCR和Western blot方法檢測肝臟脂質合成相關基因mRNA和蛋白的表達水平。結果與對照組相比,實驗組小鼠體質量和空腹血糖水平明顯升高(F=12.78~100.05,P<0.05),出現了明顯的糖耐量異常和胰島素抵抗,GTT和ITT曲線下面積明顯增大(t=3.25、4.70,P<0.05),血清中ALT、AST水平和肝臟中TG、TC水平顯著增高(t=4.52~13.12,P<0.05)。實驗組小鼠肝臟組織中M1型巨噬細胞比例明顯升高,而M2型巨噬細胞比例明顯下降(t=4.81~12.12,P<0.05),肝臟脂質合成基因SREBP1c、ACC1、FAS和CD36的mRNA和蛋白表達水平均明顯升高(t=3.54~9.96,P<0.05)。結論HCMV-IE2可能通過誘導肝臟中巨噬細胞M1型極化影響肝臟脂質代謝平衡,從而促進果糖誘導的非酒精性脂肪肝的進展。
[關鍵詞]非酒精性脂肪性肝病;果糖;小鼠,轉基因;即早蛋白質類;葡糖耐受不良;胰島素抵抗;巨噬細胞
[中圖分類號]R575.5[文獻標志碼]A
Effect of human cytomegalovirus immediate-early protein 2 on hepatic steatosis induced by fructose in UL122 transgenic model mice ZHOU Xiaoqiong, ZHANG Xianjuan, WANG Bin(School of Basic Medicine, Qingdao University, Qingdao 266071, China)
[ABSTRACT]ObjectiveTo investigate the effect of human cytomegalovirus immediate-early protein 2 (HCMV-IE2) on fructose-induced hepatic steatosis in UL122 transgenic model mice. MethodsThe UL122 transgenic mice were established as experimental group, and age-matched wild-type mice were established as control group. The mice were fed with high-fructose (60%) diet for 8 weeks, and the changes in body weight and blood glucose were monitored. After the glucose tolerance test and the insulin tolerance test were performed, the mice were sacrificed, and the serum levels of alanine aminotransferase (ALT) and aspartate aminotransferase (AST) were measured, as well as the levels of triglyceride (TG) and total cholesterol (TC) in the liver. Liver tissue was collected, and HE staining and oil red O staining were used to observe histopathological changes; flow cytometry was used to observe macrophage polarization in the liver; RT-qPCR and Western blot were used to measure the mRNA and protein expression levels of lipid synthesis-related genes in the liver. ResultsCompared with the control group, the experimental group had significant increases in body weight and fasting blood glucose level (F=12.78-100.05,P<0.05) and significantly impaired glucose tolerance and insulin resistance, with significant increases in the area under the GTT and ITT curves (t=3.25,4.70,P<0.05). There were significant increases in the serum levels of ALT and AST and the levels of TG and TC in the liver (t=4.52-13.12,P<0.05). The experimental group had a significant increase in the proportion of M1-type macrophages and a significant reduction in the proportion of M2-type macrophages in liver tissue (t=4.81-12.12,P<0.05), as well as significant increases in the mRNA and protein expression levels of the lipid synthesis genes SREBP1c, ACC1, FAS, and CD36 in the liver (t=3.54-9.96,P<0.05). ConclusionHCMV-IE2 affects the balance of liver lipid metabolism by inducing M1 polarization of macrophages in the liver, thereby promoting the progression of fructose-induced nonalcoholic fatty liver disease.
[KEY WORDS]Non-alcoholic fatty liver disease; Fructose; Mice, transgenic; Immediate-early proteins; Glucose intole-rance; Insulin resistance; Macrophages
人巨細胞病毒(HCMV)是雙鏈DNA病毒,屬皰疹病毒β亞科[1]。研究發現, HCMV感染期間機體脂肪酸生物合成增加[2],可能與糖脂代謝障礙之間存在關聯。
近幾年來,非酒精性脂肪性肝病(non-alcoholic fatty liver disease,NAFLD)在人群中的患病率呈上升趨勢,其原因可能與高糖(尤其是高果糖)飲食模式有關[3]。NAFLD的主要危險因素為慢性輕度組織炎癥[4],肝臟是機體糖脂代謝的關鍵場所,肝內巨噬細胞的活化和招募直接導致胰島素抵抗(IR),影響肝內糖脂代謝,促進NAFLD發展[5]。有研究顯示,M2型巨噬細胞向M1型巨噬細胞的轉化對IR有促進作用[6]。轉錄組學顯示HCMV感染可以上調與M1型巨噬細胞表型密切相關的基因[7]。
HCMV即刻早期蛋白2(HCMV-IE2)由病毒基因組中的UL122基因編碼表達,是HCMV感染時關鍵蛋白,在對抗宿主免疫能力中發揮重要作用。IE2蛋白功能的研究有助于深入了解HCMV感染相關疾病發病機制。前期研究表明在正常飲食的小鼠中IE2蛋白可以通過上調SREBP1c的表達促進肝臟脂滴的合成[8],但在果糖攝入條件下的影響尚不明確。
HCMV感染具有嚴格的種屬特異性,所以至今尚未能成功構建感染HCMV的動物模型。本課題組前期成功構建了能穩定表達IE2蛋白的UL122轉基因的小鼠模型[9],本研究在該模型基礎上,給予了模型小鼠8周高果糖飼料喂養,構建高果糖誘導的非酒精性脂肪肝模型,探討IE2對小鼠肝臟脂肪變性的影響。
1材料和方法
1.1主要試劑
60%高果糖純化飼料購自江蘇美迪森生物醫藥有限公司;丙氨酸氨基轉移酶(ALT)、天冬氨酸氨基轉移酶(AST)、三酰甘油(TG)和總膽固醇(TC)試劑盒購自南京建成生物工程研究所;PCR引物購自上海生工生物工程有限公司;兔抗鼠單克隆抗體Rabbit anti-ACC1、Rabbit anti-CD36和Rabbit anti-FASN均購自武漢博士德生物工程有限公司;多克隆抗體Rabbit anti-SREBP1c購自成都正能生物技術有限公司;流式細胞術(FCM)檢測使用的抗體PE-Cy7-anti-F4/80、FITC-anti-CD11c和PC5.5-anti-CD206購自美國Biolegend公司。
1.2實驗動物及分組
C57BL/6小鼠購自北京斯貝福生物技術有限公司,依據文獻[9]的方法獲得UL122轉基因陽性小鼠。隨機選取6~8周UL122轉基因陽性雄性小鼠(實驗組)和同齡野生型C57BL/6雄性小鼠(對照組)各12只,適應性喂飼1周后,開始造模,喂飼60%高果糖飼料56 d,每周定期測量小鼠體質量和血糖。
1.3小鼠的葡萄糖耐量實驗(GTT)和胰島素耐量實驗(ITT)
在造模的第52天,兩組小鼠禁食12 h后,灌胃質量濃度200 g/L葡萄糖溶液,分別于灌胃后第0、15、30、60、90、120分鐘時,從尾靜脈取血1~2 μL,用血糖試紙條測定各時間點血糖濃度,并計算GTT曲線下面積。
在造模的第55天,兩組小鼠禁食4~6 h后,腹腔注射胰島素0.75 U/kg,分別于注射后第 0、15、30、60、90、120分鐘時,從尾靜脈取血1~2 μL,以血糖試紙測定各時間點血糖濃度,并計算ITT曲線下面積。
1.4酶法檢測小鼠血清中ALT、AST水平和肝組織中TG、TC水平
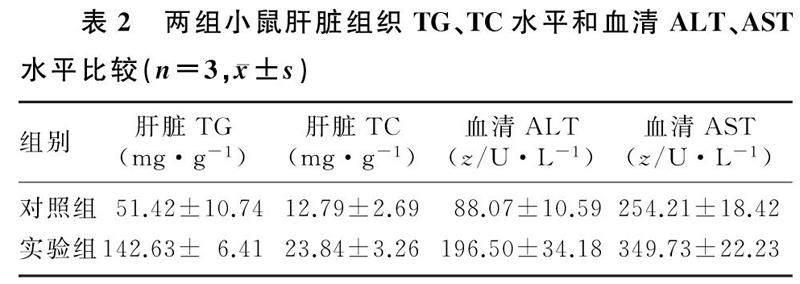
在造模后第56天,經腹腔注射戊巴比妥鈉麻醉兩組小鼠,摘眼球取血,室溫靜置2 h,3 000 r/min離心10 min,收集小鼠血清,按試劑盒說明書方法檢測血清中ALT、AST水平。脫頸處死兩組小鼠,剖取小鼠肝臟,置于-80 ℃保存備用。稱取新鮮肝臟組織,按照1 mg組織加入20 μL裂解液比例,在冰浴條件下機械勻漿,根據試劑盒說明書要求檢測組織中TG、TC的水平。
1.5油紅O染色和HE染色觀察小鼠肝臟組織病理學變化
每組隨機取3只小鼠肝臟中葉組織,以40 g/L多聚甲基醛固定24 h后,分別進行油紅0染色和HE染色,光學顯微鏡下觀察兩組小鼠肝臟組織病理學改變情況。
1.6免疫組織化學染色和FCM檢測小鼠肝臟組織巨噬細胞極化情況
每組隨機取3只小鼠部分肝臟組織,在40 g/L多聚甲醛中固定24 h。梯度乙醇脫水、透明、石蠟包埋、烤片、脫水,抗原修復后,用血清封閉30 min,加一抗F4/80(1∶500),4 ℃下孵育過夜,PBS洗滌后加相應二抗室溫孵育1 h,DAB辣根過氧化物酶顯色,封片,顯微鏡下觀察。
每組隨機取3只小鼠全部肝臟組織,200目尼龍網上低溫研磨新鮮肝臟組織,離心去除雜質及肝實質細胞,通過紅細胞裂解液去除紅細胞,以40% Percoll梯度分層,800 r/min離心5 min后,收集肝臟單個核細胞并計數。取1×106個細胞,同時加入相應細胞群的檢測抗體:以F4/80-PE-Cy7、CD11c-FITC抗體標記M1型巨噬細胞,F4/80-PE-Cy7和CD206-PC5.5抗體標記M2型巨噬細胞。隨后避光染色30 min,PBS洗滌重懸細胞后200目尼龍網過濾,經FCM檢測M1型和M2型巨噬細胞比例。
1.7實時熒光定量PCR(RT-qPCR)方法檢測各組小鼠肝臟組織當中的腫瘤壞死因子-α(TNF-α)、白細胞介素-6(IL-6)、IL-10、固醇調節元件結合蛋白(SREBP1c)、脂肪酸合酶(FAS)、乙酰輔酶A羧化酶(ACC1)和脂肪酸轉運體(CD36)基因表達
每組隨機取3只小鼠部分肝臟組織,用Trizol法提取小鼠肝臟總RNA,逆轉錄合成cDNA,RT-qPCR方法檢測小鼠肝臟組織中M1型、M2型巨噬細胞極化基因的mRNA表達,引物序列詳見表1。RT-qPCR的反應條件如下:95 ℃預變性30 s;然后95 ℃變性10 s,65 ℃退火30 s,共進行40個循環。以GAPDH作為內參基因,目的基因的相對表達水平通過2-△△CT計算。
1.8Western blot方法檢測小鼠肝臟組織之中SREBP1c、FASN、ACC1和CD36蛋白的表達
每組隨機取3只小鼠部分肝臟組織,用RIPA抽取總蛋白,取蛋白樣品行SDS-PAGE凝膠電泳,濕電轉印至PVDF膜,將PVDF膜在含體積分數為0.05的脫脂牛奶中封閉2 h;加入兔抗鼠SREBP1c多克隆抗體(1∶1 000)、兔抗鼠FASN單克隆抗體(1∶1 000)、兔抗鼠ACC1單克隆抗體(1:1000)、兔抗鼠CD36單克隆抗體(1∶1 000),于4 ℃下孵育過夜;以TBST溶液清洗后,加入相應的羊抗兔二抗(1∶5 000),室溫下孵育2 h;洗膜,以化學發光劑處理,顯影,用Image J軟件分析條帶灰度值,計算目的蛋白的相對表達量。
1.9統計學處理
使用Graphpad prism 5和SPSS 26軟件對數據進行整理和分析,組間比較采用非配對的t檢驗和重復測量設計的方差分析,以P<0.05為差異有統計學意義。
2結果
2.1HCMV-IE2對小鼠體質量、血糖和IR的影響
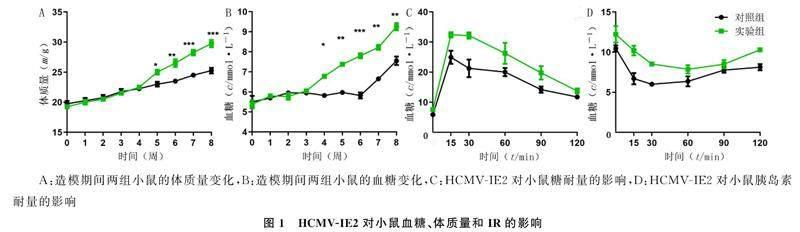
重復測量設計方差分析顯示,時間、組別及時間與組別交互作用對兩組小鼠體質量及空腹血糖水平均有顯著影響(F組別=10.81~67.13,F時點=93.80~154.50,F時點*組別=17.13~19.92,P<0.05);單獨效應分析顯示,第5周以后,實驗組小鼠體質量明顯大于對照組(F=12.78~45.00,P<0.05),見圖1A。第6周以后,實驗組小鼠空腹血糖水平明顯高于對照組(F=36.89~100.05,P<0.05),見圖1B。在灌胃葡萄糖后,實驗組小鼠血糖水平較對照組小鼠明顯升高,對照組和實驗組小鼠的GTT曲線下面積分別為2 100.66±279.65、2 856.33±289.54,兩組比較差異有顯著性(t=3.25,P<0.05),見圖1C。在腹腔注射胰島素后,與對照組小鼠比較,實驗組血糖水平下降幅度較小,對照組和實驗組的ITT曲線下面積分別為860.03±35.30、1 082.00±73.72,兩組比較差異有顯著性(t=4.70,P<0.01),見圖1D。
2.2高果糖條件下HCMV-IE2對小鼠肝臟組織病理學的影響
HE染色結果顯示,實驗組小鼠肝臟組織出現大面積脂肪樣空泡,細胞核被擠向邊緣;對照組小鼠僅出現一些散在的、面積較小的空泡;油紅O染色結果顯示,實驗組小鼠肝臟切面可見大面積的脂滴分布,而對照組小鼠肝臟僅出現了大小不等的散在紅色脂滴(圖2)。實驗組小鼠肝臟組織TG、TC水平和血清ALT、AST水平均顯著高于對照組(t=4.52~13.12,P<0.01),見表2。
2.3高果糖條件下HCMV-IE2對肝臟組織中巨噬細胞M1/M2型極化的影響
免疫組織化學染色結果顯示,實驗組肝臟組織中巨噬細胞(F4/80)浸潤明顯多于對照組(圖3)。同時,FCM和RT-qPCR檢測結果顯示,與對照組比較,實驗組小鼠肝臟組織中M1型巨噬細胞的比例及相關標志物TNF-α和IL-6 mRNA的水平顯著增高(t=4.87~9.65,P<0.05),而M2型巨噬細胞的比例及其相關標志物IL-10? mRNA的水平顯著降低(t=4.81~12.12,P<0.05),見表3。
2.4高果糖條件下HCMV-IE2對肝臟脂質代謝相關基因和蛋白表達的影響
RT-qPCR檢測結果顯示,與對照組相比,實驗組小鼠肝臟組織中SREBP1c、ACC1、FAS、CD36 mRNA表達水平均上調(t=3.54~5.69,P<0.05),見表4。Western blot結果顯示,與對照組相比,實驗組小鼠肝臟組織中SREBP1c、ACC1、FASN和CD36的蛋白表達水平均上調,SREBP1c、ACC1和FASN蛋白表達水平顯著升高(t=7.94~9.96,P<0.05),見表4、圖4。
3討論
由于HCMV感染具有高度的種屬特異性,目前尚未建立感染HCMV的動物模型。目前關于HCMV與糖脂代謝紊亂機制的研究主要還是局限在細胞水平方面,很難進行體內研究。UL122轉基因小鼠的構建克服了物種特異性難題,為研究IE2對機體代謝紊亂的影響提供了有效的途徑。本研究結果顯示,在果糖誘導的非酒精性脂肪肝模型小鼠中,IE2加速了血糖的升高,促進了IR和肝臟脂肪變性。在果糖喂養的UL122轉基因小鼠肝臟組織中,巨噬細胞浸潤明顯多于對照組,且表現出獨特M1/M2極化現象,趨向于向M1型巨噬細胞極化。以上結果表明HCMV編碼的IE2蛋白可通過誘導M1型巨噬細胞極化促進果糖誘導的肝脂肪病變。
臨床研究表明,HCMV感染的肥胖和肝損傷患者通常伴有脂質代謝紊亂。然而,HCMV感染引起人體脂質代謝紊亂的分子機制尚不清楚。HCMV感染的一個特征是它以有利于病毒復制的方式調節感染細胞的新陳代謝,HCMV感染已被證明可誘導機體葡萄糖轉運蛋白4(GLUT4)的表達及其易位到細胞表面,從而導致用于脂肪酸從頭合成的細胞質葡萄糖增加[10-11]。生物體合成的過多脂肪酸為病毒包膜的形成提供了充足的條件[2,12]。
巨噬細胞是機體最重要的免疫細胞之一,對于穩定機體內部環境和保護身體免受外界入侵具有至關重要的作用,它可極化為不同的類型并發揮不同的功能。其中M1型巨噬細胞在促炎方面起到了非常重要的作用,而M2型巨噬細胞則表現出抗炎、促進組織重塑等特點[13]。研究顯示,非酒精性脂肪性肝炎(NASH)模型小鼠肝臟中常伴隨著M1/M2型巨噬細胞百分比的增高。也有研究顯示,清除小鼠肝臟巨噬細胞可以減少因高脂肪飲食而導致的肝臟脂質積累和IR,從而降低NASH的發生風險[14]。因此肝臟巨噬細胞所特有的M1/M2型極化狀態在NAFLD的發生發展中扮演著重要角色。
在HCMV感染期間,巨噬細胞是其作用的第一個靶標,對HCMV持久存在和傳播是必不可少的[15-16]。CHAN等[17]發現HCMV感染單核細胞4 h后,與經典M1型巨噬細胞極化相關的65%的基因被上調,而與經典M2型巨噬細胞極化相關的基因只有4%被上調。另外有研究顯示,NF-κB活性對于受HCMV感染的單核細胞偏向M1型促炎巨噬細胞極化至關重要,約79%的HCMV誘導的M1型巨噬細胞表型相關基因以NF-κB依賴性方式調節[7]。本研究結果顯示,與喂養高果糖的野生型小鼠相比,喂養高果糖的UL122轉基因小鼠的巨噬細胞表現出獨特的M1/M2極化現象,在肝臟組織中趨向于M1型巨噬細胞極化。因此可以推測,這種獨特的M1型極化可能是由IE2誘導的,但其具體機制還有待后續實驗深入研究。M1型巨噬細胞可釋放多種炎性遞質,如促炎細胞因子和趨化因子等[18]。因此本研究又進一步檢測了肝組織中M1型巨噬細胞標志物TNF-α和IL-6 mRNA的水平,結果顯示,喂養高果糖的UL122轉基因小鼠肝組織中TNF-α以及IL-6 mRNA的表達均顯著升高。SREBP1c是參與調控脂肪酸、TG和TC合成的轉錄因子,該因子能夠激活其下游的目標基因,并且增強FASN、ACC1、CD36等多種參與脂肪酸從頭合成的基因的表達[19]。SREBP1c及其下游靶基因FASN以及ACC1在NAFLD患者的肝臟組織中表達水平高于正常人群[20]。本研究發現喂養高果糖的UL122轉基因小鼠肝組織當中的SREBP1c、FASN和ACC1的mRNA表達水平明顯上調。推測IE2誘導的MI型巨噬細胞極化可能與此有關,可能是由于其促進了炎癥細胞因子的分泌,并同時上調了脂質生成相關基因的表達所致,進一步促進了脂肪從頭合成,增強了肝細胞的脂質合成代謝,最終導致更多的脂質沉積。
綜上所述,HCMV-IE2通過誘導肝組織中巨噬細胞向M1型極化影響肝臟脂質代謝平衡,從而促進果糖誘導的NAFLD的進展,這一發現為HCMV感染引起的糖脂代謝紊亂機制的研究提供了新的研究方向。
倫理批準和動物權利聲明:本研究涉及的所有動物實驗均已通過青島大學實驗動物福利倫理委員會的審核批準(20220308C57402022-0907107)。所有實驗過程均遵照《實驗動物管理條例》的規定進行。
作者聲明:周曉瓊、王斌均參與了研究設計;周曉瓊、張現娟、王斌參與了論文的寫作和修改。所有作者均閱讀并同意發表該論文,且均聲明不存在利益沖突。
[參考文獻]
[1]MURPHY E, RIGOUTSOS I, SHIBUYA T, et al. Reeva-luation of human cytomegalovirus coding potential[J]. Proc Natl Acad Sci USA, 2003,100(23):13585-13590.
[2]YU Y J, MAGUIRE T G, ALWINE J C. Human cytomegalovirus infection induces adipocyte-like lipogenesis through activation of sterol regulatory element binding protein 1[J]. J Vi-rol, 2012,86(6):2942-2949.
[3]YILMAZ Y. Review article: Fructose in non-alcoholic fatty liver disease[J]. Aliment Pharmacol Ther, 2012,35(10):1135-1144.
[4]PELLEGRINELLI V, RODRIGUEZ-CUENCA S, ROUAU-LT C, et al. Dysregulation of macrophage PEPD in obesity determines adipose tissue fibro-inflammation and insulin resis-tance[J]. Nat Metab, 2022,4(4):476-494.
[5]SAMUEL V T, SHULMAN G I. The pathogenesis of insulin resistance: Integrating signaling pathways and substrate flux[J]. J Clin Invest, 2016,126(1):12-22.
[6]CHEN L Y, ZHU Y X, HOU X H, et al. The role of gut bacteria and fungi in alcohol-associated liver disease[J]. Front Med (Lausanne), 2022,9:840752.
[7]CHAN G, BIVINS-SMITH E R, SMITH M S, et al. NF-kappaB and phosphatidylinositol 3-kinase activity mediates the HCMV-induced atypical M1/M2 polarization of monocytes[J]. Virus Res, 2009,144(1-2):329-333.
[8]ZHANG X J, WANG Y Y, QIAN D M, et al. HCMV-encoded IE2 promotes NAFLD progression by up-regulation of SREBP1c expression in UL122 genetically modified mice[J]. Int J Clin Exp Pathol, 2018,11(8):4213-4220.
[9]張現娟. HCMV IE2對UL122轉基因小鼠糖脂代謝的影響研究[D]. 青島:青島大學.
[10]MUNGER J, BAJAD S U, COLLER H A, et al. Dynamics of the cellular metabolome during human cytomegalovirus infection[J]. PLoS Pathog, 2006,2(12):e132.
[11]MUNGER J, BENNETT B D, PARIKH A, et al. Systems-level metabolic flux profiling identifies fatty acid synthesis as a target for antiviral therapy[J]. Nat Biotechnol, 2008,26(10):1179-1186.
[12]SPENCER C M, SCHAFER X L, MOORMAN N J, et al. Human cytomegalovirus induces the activity and expression of acetyl-coenzyme A carboxylase, a fatty acid biosynthetic enzyme whose inhibition attenuates viral replication[J]. J Virol, 2011,85(12):5814-5824.
[13]MURRAY P J. Macrophage polarization[J]. Annu Rev Phy-siol, 2017,79:541-566.
[14]HUANG W, METLAKUNTA A, DEDOUSIS N, et al. Depletion of liver Kupffer cells prevents the development of diet-induced hepatic steatosis and insulin resistance[J]. Diabetes, 2010,59(2):347-357.
[15]BAYER C, VARANI S, WANG L, et al. Human cytomegalovirus infection of M1 and M2 macrophages triggers inflammation and autologous T-cell proliferation[J]. J Virol, 2013,87(1):67-79.
[16]MOORE P S, BOSHOFF C, WEISS R A, et al. Molecular mimicry of human cytokine and cytokine response pathway genes by KSHV[J]. Science, 1996,274(5293):1739-1744.
[17]CHAN G, NOGALSKI M T, STEVENSON E V, et al. Human cytomegalovirus induction of a unique signalsome during viral entry into monocytes mediates distinct functional changes: A strategy for viral dissemination[J]. J Leukoc Biol, 2012,92(4):743-752.
[18]DEVISSCHER L, VERHELST X, COLLE I, et al. The role of macrophages in obesity-driven chronic liver disease[J]. J Leukoc Biol, 2016,99(5):693-698.
[19]FERR P, FOUFELLE F. Hepatic steatosis: A role for de novo lipogenesis and the transcription factor SREBP-1c[J]. Diabetes Obes Metab, 2010,12:83-92.
[20]SCORLETTI E, BYRNE C D. Omega-3 fatty acids and no-n-alcoholic fatty liver disease: Evidence of efficacy and mechanism of action [J]. Molecular aspects of medicine, 2018,64:135-146.
(本文編輯 耿波 厲建強)
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
學苑創造·A版(2020年9期)2020-10-13 09:41:02
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年12期)2019-05-21 02:55:32
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
學苑創造·A版(2015年11期)2016-01-14 09:03:27
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
七彩語文·畫刊(2012年4期)2012-04-29 00:00:00
