
份香蕉種質資源SRAP分子標記親緣關系分析
2024-06-09 15:12:40周海琪夏玲呂順曾莉莎王芳黃曉彥陳東儀劉文清梁少麗劉麗琴
熱帶作物學報 2024年4期
周海琪 夏玲 呂順 曾莉莎 王芳 黃曉彥 陳東儀 劉文清 梁少麗 劉麗琴

關鍵詞:香蕉;SRAP;聚類分析;親緣關系
中圖分類號:S668.1 文獻標志碼:A
香蕉是目前世界上產量和貿易量最大的水果之一,也是熱帶、亞熱帶地區重要的經濟作物和糧食作物[1]。香蕉味道香甜,營養豐富,物美價廉,深得人們喜愛,而且香蕉植株優美,也常被人們用來裝點環境。我國是香蕉的原產地之一,具有豐富的種質資源,同時也具有悠久的栽培歷史。目前香蕉的栽培品種主要有香牙蕉、龍牙蕉、貢蕉、粉蕉和大蕉,這些品種基本由尖葉蕉(Musaacuminata Colla,A 基因組)和長梗蕉(Musabalbisiana Colla,B 基因組)這2 個野生品種發展而來[2],染色體倍性包含二倍體、三倍體(主要)和四倍體。由于來源、地域、育種手段等因素,不同的香蕉品種間的遺傳背景存在差異。香蕉一直沿用SIMMONDS[3]的分類方法來識別和分類不同的香蕉品種,但是隨著研究的深入和研究技術手段的更新,這種單一的、依靠形態特征的分類方法已經無法滿足對香蕉親緣關系和遺傳多樣性等研究的需求,甚至對研究造成一定的阻礙,如香蕉品種的分類分組與基因組組成不完全一致[4],影響香蕉種質資源的合理利用。因此,結合分子標記技術鑒定香蕉種質資源、分析其親緣關系和研究遺傳多樣性是非常必要的。
國內外關于運用分子標記技術來研究香蕉種質的報道眾多,SSR、AFLP、ISSR、RAPD、SRAP等分子標記[5-9]被應用在分析香蕉品種(系)的種群鑒定與分類、遺傳多樣性等方面。SRAP 技術是一種基于PCR 的分子標記技術[10],具有簡便、快速、引物設計簡便、多態性和信息量豐富等特點,廣泛應用于園藝植物遺傳育種[11]、作物改良[12]、果樹種質資源鑒定和親緣關系分析[13-17]等方面。謝子四等[18]利用SRAP標記對29 份香蕉材料進行了基因組多態性分析,擴增出324 條可辨認條帶,多態性比率為80.86%;漆艷香等[9]應用SRAP 標記對21 份香蕉種質的遺傳多態性進行分析。但這些報道都以少量或地方代表的個別品種為試驗材料,不夠系統和全面。本研究采用SRAP 分子標記技術,以89 份來至國內外的香蕉種質,包括香牙蕉、龍牙蕉、貢蕉、大蕉、粉蕉和不同類型的野蕉等多種香蕉類型為材料,探討各類型、品種之間的親緣關系,為香蕉育種、品種鑒定、新品種保護等提供分子水平上的依據。
1 材料與方法
1.1 材料
供試89 份材料取自東莞市農業科學研究中心的香蕉種質資源圃和麻涌試驗基地,89 份香蕉種質資源樣品的基因型和來源見表1,SRAP 引物序列由英濰捷基(上海)貿易有限公司合成,DL2000 DNA marker、dNTPs、Taq DNA 聚合酶,10×Buffer 等均購自TranGene 公司。
1.2 方法
1.2.1 香蕉基因組DNA 的提取及檢測 取香蕉幼嫩的新葉,采用改良的CTAB 法[19]提取DNA,經蛋白核酸檢測儀檢測DNA 濃度與純度后,用0.8%瓊脂糖凝膠電泳檢測DNA 質量,最后將DNA 稀釋至所需濃度置于?20 ℃冰箱貯存備用。
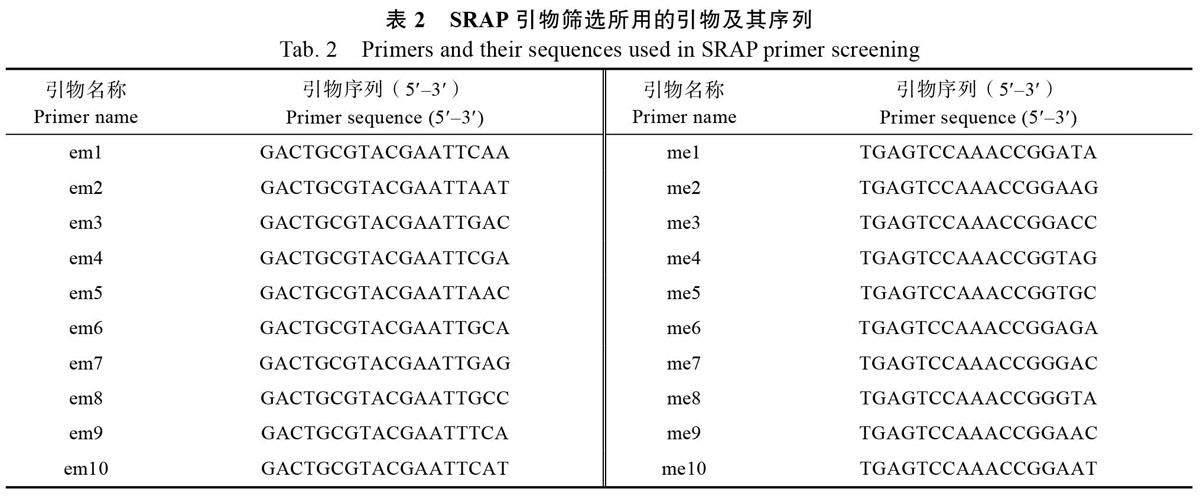
1.2.2 SRAP 引物的篩選及PCR 擴增 運用SRAP 分子標記引物(10 個正向引物和10 個反向引物兩兩組合成100 對引物組合,表2),以5 份香蕉品種為模板,對100 對引物進行篩選,每個引物重復2~3 次,篩選能擴增出多態性好且條帶多的最適合引物組合。PCR 擴增反應在Bio-lePCR 擴增儀上進行,反應體系:20 μL 的PCR 反應體系含有10×buffer (含Mg2+)2 μL,0.2 mmol/LdNTPs(2.5 nmol/L)2 μL,上、下引物(10 μmol/L)各1 μL,DNA(20 ng/L)2 μL 及Taq DNA 聚合酶1 μL。反應程序為94 ℃預變性5 min;94 ℃變性1 min,35 ℃退火1 min,72 ℃延伸1 min,5 個循環;94 ℃變性1 min,50 ℃退火1 min,72 ℃延伸1 min,循環35 次;72 ℃延伸10 min。擴增反應結束后,加入4 μL 6×loading buffer,取8~10 μL 擴增產物在1.8%瓊脂糖凝膠中電泳,電極緩沖液為0.5×TBE,在凝膠成像儀中觀察并照相。
1.3 數據處理
經過2~3 次重復,統計穩定可靠的多態性位點,擴增條帶在同一位置不同樣品中,有條帶記為“1”,無條帶記為“0”,建立(0,1)矩陣,用Ntsys 軟件采用UPGMA(unweghed pair methodairthtnetic average)進行聚類分析,構建樹狀圖。
2 結果與分析
2.1 89 份香蕉種質資源基因組DNA 的檢測
將提取的89 份香蕉樣品基因組DNA 采用0.8%瓊脂糖凝膠電泳進行檢測,結果顯示香蕉基因組DNA 主帶明顯,呈現一條完整發亮的譜帶,無降解拖尾現象。經核酸蛋白儀檢測后,所得DNA母液OD260/OD280 在1.8 左右,OD260/OD230≥2.0,濃度在1000~1800 ng/μL 之間,表明所提取的香蕉基因組DNA 完整性較好,純度高,無小分子物質和蛋白質的污染,能夠滿足后續試驗要求。將DNA 分裝至PCR 管中備用,用時稀釋至30 ng/μL。
2.2 香蕉SRAP 引物篩選及PCR 擴增
利用表2 所列的正反引物各10 條,共100 對引物組合進行篩選,以其中擴增條帶易于識別、帶型清晰、多態性高的13 對引物對供試材料進行分析。結果表明,13 對引物共擴增出170 條條帶,平均每對引物13.08 條,其中多態性帶共140 條(圖1,圖2)。各引物產生的多態性帶比例為64.3%~92.9%,平均為79.0%(表3)。
2.3 遺傳相似性系數分析
在遺傳相似系數矩陣中,89 份香蕉種質資源的相似系數的變化范圍在0.241~1.000 之間,在這些香蕉種質中,8818 與天寶野蕉、8818 與羅壩大蕉、羅壩大蕉與夫人指、畦頭大蕉與紅研2 號、矮大蕉與粵豐1 號的相似系數最低,為0.241。海貢和貢蕉相似性系數最高,為1.000,其SRAP 分子標記帶型基本一致,但在田間表現中,海貢和貢蕉是有一定差異的,海貢具有枯萎病抗性,而貢蕉是感病品種。究其原因一方面可能我們采用的SRAP 分子標記位點還比較少,不足以區分這2 個品種,另一方面也有可能樣品混淆,需要進一步實驗證明。此外,分別來自于云南和廣西的云南那邦和廣西憑祥相似性系數為0.983,同時來自于云南的野蕉1 號與這2 個種質資源的相似性系數也在0.900 以上,這3 個品種均為BB 類野蕉,親緣關系較近。五指山1 號和五指山2 號相似性系數為0.983,五指山3 號與五指山1 號和2 號的相似性系數相對較低,分別是0.81 和0.828,親緣關系也比較近,五指山1 號、五指山2 號及五指山3 號是課題組在海南五指山不同地點采的樣品,均為阿寬蕉類野生蕉。可見這些種質資源的遺傳多樣性非常豐富,香蕉種質資源的親緣關系很復雜,而并不是簡單的平行關系。
2.4 親緣關系聚類分析
采用NTSYSpc 2.10 t 軟件對SRAP 標記統計后的綜合結果進行數據分析,建立樹狀圖(圖3)。在相似性系數為0.49 時,可以將89 份香蕉種質資源分為六大類群。
第一大類群:主要包括香牙蕉、貢蕉、龍牙蕉、AA 型野蕉,基因型主要為AAA 型、AAB型和AA 型,共33 份種質資源,在相似性系數為0.65 處可以將此大類分為四大亞類。第一亞類包括G30、NK4-1、海貢、貢蕉、河口龍牙(貴妃蕉)、夫人指、1442、漳選2 號、漳選8-2、1443、8818、巴西、天寶矮蕉、大蜜舍、泰國、菲律賓、粵優抗1 號、泰國貢蕉、catas1、佳麗、LF、飛亞(飛亞-25)、粵豐1 號、catas5、金指、紫莖蕉、金沙香、玫瑰蕉、紅研2 號;第二亞類包括佛手蕉、過山香;第三亞類是千指蕉;第四亞類是紫花蕉(美葉芭蕉)。
第二大類群:主要包括粉蕉、粉大蕉、BB型野蕉,基因型主要為ABB 型,共21 份種質資源;在相似性系數為0.64 處可以將此大類分為二大亞類。第一亞類包括粉雜1 號、孟加拉粉大蕉、千蒡培、灰大蕉、增城野蕉、云南那邦、廣西憑祥、野蕉1 號、孟加拉菜、五山野蕉、馬來西亞粉蕉、BB 野蕉、南海粉大蕉、東莞粉大蕉、銀豐1 號、棱指蕉、四方大蕉、大粉蕉;第二亞類包括天寶野蕉、東莞細粉、沙巴。
第三大類群:主要包括大蕉、阿寬蕉類野蕉以及芭蕉類野蕉,共32 份種質資源。在遺傳距離為0.64 處可以將此大類分為五大亞類。第一亞類包括云南牛角蕉;第二亞類包括矮大蕉、羅壩大蕉、飯芭蕉、三江大蕉、番禺大蕉、廣西中把大蕉、封開大蕉、東莞高把大蕉、興隆酸蕉、木棉蕉、牛芭蕉、中山大蕉、潮州大蕉、海南芭蕉、柴蕉、巴馬高把大蕉、五指山1 號、五指山2 號、五指山3 號、防城大蕉、海南牛角蕉、畦頭大蕉;第三亞類包括阿寬野蕉、龍巖野蕉、三明野蕉、云南漾濞、廣寧野蕉;第四亞類包括賓陽野蕉、龍州野蕉;第五亞類包括盛2、芭蕉。
第四、五、六類群:都只包含了1 份種質資源,分別是紅花蕉、地涌金蓮、蝎尾蕉。
3 討論
香蕉種質資源豐富,種類品種繁多,然而由于特殊的生物學特性如多倍體、無性繁殖、營養結實、小染色體等[18],使其種質資源的分類及系統學研究一直是個難題。本研究以香牙蕉、龍牙蕉、大蕉、粉蕉和不同類型的野蕉,基因型主要包括AA、BB、AAA、AAB、ABB 和AAAA 等類型的89 份香蕉種質資源為材料,運用SRAP 分子標記技術研究不同香蕉種質資源之間的親緣關系。由10 個正向引物和10 個反向引物兩兩組合成100 對引物組合,從中篩選出13 對擴增條帶易于識別、帶型清晰、多態性高的SRAP 引物,這13 對引物在89 個香蕉種質資源間擴增出170 條可辨認條帶,其中表現出多態性的帶有140 條,多態性比率為79.00%。
在相似性系數為0.49 時,可以將這89 份香蕉種質資源分成六大類,包括:香牙蕉、貢蕉、龍牙蕉、AA 型野蕉為主的第一大類(AAA、AA、AAB 基因型型),粉大蕉、粉蕉和BB 型野蕉為主的第二大類(ABB、BB 基因型),大蕉、阿寬蕉類野蕉為主的第三大類,紅花蕉、地涌金蓮、蝎尾蕉、分別所屬的第四、五、六類群,該方法的分類結果與傳統的基于形態的分類結果基本一致,說明用SRAP 標記對香蕉種質資源進行分類是可行的。但該研究中發現部分香蕉種質的分類分組并沒有與品種的基因組組成一致,例如,以巴西為代表的AAA 型香牙蕉、以貢蕉為代表的AA 型及以龍牙蕉為代表的AAB 型聚類群體并不明顯,均分散在第一類。三者在遺傳上沒有太大的差異,說明栽培類香牙蕉、貢蕉和龍牙蕉具有相同的祖先,與AA 基因型野蕉親緣關系近,這與寧淑萍等[20]利用SSR 技術分析的結果一致。粉蕉及粉大蕉(ABB 型)聚為一類,可見二者遺傳背景相似,與BB 基因型的野蕉親緣關系較近,但無法明顯區別是否含A 基因組,這與郭計華[21]的研究結果相似。不同基因組成的香蕉品種聚到一起,也說明香蕉品種的親緣關系可能與起源有關。前人利用細胞遺傳學方法[22]、RFLP 分子標記[23-24]和SSR分子標記[25-26]方法對香蕉品種資源進行分類分析,也發現與應用傳統SIMMONDS[3]分類法命名的基因組型結果不一致。香牙蕉(AAA)、大蕉(ABB)、粉蕉(ABB)和龍牙蕉(AAB)都是根據形態學性狀分類而籠統地確定基因組類型,難免會造成同物異名、同名異物的現象[27],也說明使用單一的分類方法無法明確地把香蕉種質區分,還需要從形態學、細胞學、分子生物學方面作進一步的研究和鑒定。
早期研究認為大蕉基因組為ABB 或者BBB,項目組前期研究發現大蕉倍性值(或DNA 相對含量)平均高于其他ABB 基因型資源,而且利用大蕉與長梗蕉人工雜交的后代均為四倍體,而粉蕉與長梗蕉的雜交后代為三倍體或四倍體[28],說明大蕉與ABB 基因型資源有所差別。但是,項目組在進行B 基因組相關的SCAR 標記的研究中又發現大蕉是含有A 和B 基因組的[29]。在本研究中21 份大蕉和10 份阿寬蕉聚到同一類,說明大蕉和阿寬蕉親緣關系較近,但是從整個聚類圖中無法看出大蕉是否含有A 和B 基因組,可見,大蕉的遺傳背景及起源比較復雜,有待進一步研究。
