黃芪甲苷調控Nrf2/HO-1信號通路對血管內皮細胞氧化損傷的影響
2024-09-30 00:00:00譚維傅馨瑩楊仁義馬露丁煌劉曉丹張偉
湖南中醫藥大學學報 2024年9期
〔摘要〕 目的 基于細胞實驗研究黃芪甲苷(astragaloside Ⅳ, AST-Ⅳ)調控核轉錄因子紅系2相關因子2/血紅素加氧酶-1(nuclear factor-erythroid 2-related factor 2/heme oxygenase-1, Nrf2/HO-1)信號通路對血管內皮氧化損傷的影響。方法 采用1-棕櫚酰基-2-(5'-氧-戊酰基)-sn-甘油-3-磷酸膽堿[1-palmitoyl-2-(5-oxovaleroyl)-sn-glycero-3-phosphocholine, POVPC]刺激血管內皮細胞24 h建立血管內皮細胞氧化損傷模型,隨機分為模型(Model)組(35 μmol/L POVPC)、AST-Ⅳ低劑量(AST-L)組(35 μmol/L POVPC+50 μmol/L AST-Ⅳ)、AST-Ⅳ高劑量(AST-H)組(35 μmol/L POVPC+100 μmol/L AST-Ⅳ)及AST-Ⅳ高劑量+ML385(Nrf2抑制劑)(AST-H+ML385)組(35 μmol/L POVPC+100 μmol/L AST-Ⅳ+5 μmol/L ML385),以正常未處理細胞作為對照(Control)組。免疫熒光鑒定大鼠胸主動脈內皮細胞(aortic vascular endothelium cell, AVEC),CCK-8檢測AST-Ⅳ細胞增殖情況,Transwell小室檢測AVEC遷移情況,Matrigel基質膠檢測細胞血管形成功能,鬼筆環肽染色檢測細胞骨架結構,DCFH-DA熒光探針檢測細胞活性氧(reactive oxygen species, ROS)水平,試劑盒法檢測細胞培養上清液中超氧化物歧化酶(superoxide dismutase, SOD)水平,RT-qPCR和Western blot檢測Nrf2和HO-1的 mRNA及蛋白表達情況。結果 免疫熒光鑒定所提取的細胞,可特異性表達血管性血友病因子(von willebrand factor, vWF),鑒定為AVEC。與Control組相比,Model組細胞活力、遷移能力和血管形成功能顯著下降(P<0.01),細胞骨架破壞明顯,細胞內ROS水平顯著上升(P<0.01),細胞培養上清液中SOD含量顯著減少(P<0.01),細胞中Nrf2和HO-1的mRNA及蛋白表達水平下調(P<0.01或P<0.05)。與Model組比較,AST-L組及AST-H組細胞活力、遷移能力和血管形成功能顯著升高(P<0.01),細胞骨架破壞得到較好修復,細胞內ROS水平顯著下降(P<0.01),細胞培養上清液中SOD含量顯著增加(P<0.01),細胞中Nrf2和HO-1的mRNA及蛋白表達水平顯著上調(P<0.01),且以AST-H組效果更為顯著(P<0.01)。與AST-H組比較,AST-H+ML385組遷移能力和血管形成功能顯著下降(P<0.01),細胞骨架破壞增加,細胞內ROS水平顯著上升(P<0.01),細胞培養上清液中SOD含量顯著減少(P<0.01),細胞中Nrf2和HO-1的mRNA及蛋白表達水平顯著下調(P<0.01)。結論 AST對血管內皮氧化損傷具有保護作用,且有一定的劑量依賴性,其機制可能是通過激活Nrf2/HO-1信號通路發揮作用。
〔關鍵詞〕 心血管疾病;黃芪甲苷;Nrf2/HO-1信號通路;氧化損傷;血管內皮細胞
〔中圖分類號〕R285.5 〔文獻標志碼〕A 〔文章編號〕doi:10.3969/j.issn.1674-070X.2024.09.006
Effects of astragaloside Ⅳ on oxidative damage of vascular endothelial cells by regulating Nrf2/HO-1 signaling pathway
TAN Wei1,2, FU Xinying1,2, YANG Renyi1,2, MA Lu1,2, DING Huang1,2, LIU Xiaodan1,2, ZHANG Wei1,2*
1. School of Integrated Chinese and Western Medicine, Hunan University of Chinese Medicine, Changsha, Hunan 410208, China; 2. Hunan Key Laboratory of Integrated Chinese and Western Medicine for Prevention and Treatment of Heart and
Brain Diseases, Changsha, Hunan 410208, China
〔Abstract〕 Objective To explore the effects of astragaloside Ⅳ (AST-Ⅳ) on vascular endothelial oxidative damage by modulating the nuclear factor-erythroid 2-related factor 2/heme oxygenase-1 (Nrf2/HO-1) signaling pathway, based on cell experiments. Methods An endothelial cell oxidative damage model was established by stimulating vascular endothelial cells with 1-palmitoyl-2-(5-oxovaleroyl)-sn-glycero-3-phosphocholine (POVPC) for 24 hours. The cells were randomly divided into model group (35 μmol/L POVPC), low-dose AST-Ⅳ (AST-L) group (35 μmol/L POVPC+50 μmol/L AST-Ⅳ), high-dose AST-Ⅳ (AST-H) group (35 μmol/L POVPC+100 μmol/L AST-Ⅳ), and high-dose AST-Ⅳ+ ML385 (Nrf2 inhibitor) (AST-H+ML385) group (35 μmol/L POVPC+100 μmol/L AST-Ⅳ+5 μmol/L ML385). Untreated normal cells were used as the control group. Rat aortic vascular endothelium cells (AVECs) were identified by immunofluorescence. The proliferation of AST-Ⅳ cells was tested by CCK-8 assay. Transwell chambers were utilized to evaluate AVEC migration, while Matrigel matrix gel was employed to assess angiogenic function of the cells. The cytoskeletal structure was determined by phalloidin staining, reactive oxygen species (ROS) levels were measured by the DCFH-DA fluorescence probe, and superoxide dismutase (SOD) levels in cell culture supernatants were measured by kit method. The mRNA and protein expressions of Nrf2 and HO-1 were determined by RT-qPCR and Western blot. Results The expression of extracted cell surface marker von willebrand factor (vWF) was determined by immunofluorescence and was identified as AVEC. Compared with control group, the cell viability, migration ability, and angiogenic function in Model group decreased significantly (P<0.01), with obvious cytoskeleton disruption, a significant increase in intracellular ROS level (P<0.01), a significant decrease in SOD content in the cell culture supernatant (P<0.01), and downregulated mRNA and protein expression levels of Nrf2 and HO-1 in cells (P<0.01 or P<0.05). Compared with model group, AST-L group and AST-H group showed significantly increased cell viability, migration ability, and angiogenic function (P<0.01), with better repair of cytoskeleton disruption, a significant decrease in intracellular ROS level (P<0.01), a significant increase in SOD content in the cell culture supernatant (P<0.01), and significantly upregulated mRNA and protein expression levels of Nrf2 and HO-1 in cells (P<0.01), with more pronounced effects in AST-H group (P<0.01). Compared with AST-H group, AST-H+ML385 group showed significantly decreased migration ability and angiogenic function (P<0.01), increased cytoskeleton disruption, a significant increase in intracellular ROS level (P<0.01), a significant decrease in SOD content in the cell culture supernatant (P<0.01), and significantly downregulated mRNA and protein expression levels of Nrf2 and HO-1 in cells (P<0.01). Conclusion AST has protective effects on vascular endothelial oxidative damage, with a certain dose dependence, and its mechanism may involve the activation of Nrf2/HO-1 signaling pathway.
〔Keywords〕 cardiovascular disease; astragaloside Ⅳ; Nrf2/HO-1 signaling pathway; oxidative damage; vascular endothelial cell
心血管疾病(cardiovascular disease, CVD)是醫療領域的棘手問題,其死亡率在我國城鄉居民疾病死亡構成比中占據首位[1]。研究發現,引起CVD發生發展的病理因素眾多,發病機制復雜,其中,血管內皮損傷是CVD的始動因素與共同病理基礎,可引發血液流變學及血液動力學異常,引起糖尿病、高血壓和動脈粥樣硬化(atherosclerosis, AS)等疾病,推動心血管危機事件的發生[2-4]。血管內皮細胞(vascular endothelial cell, VEC)是血管內皮的主要組成部分,當氧化還原平衡被破壞時,抗氧化物質超氧化物歧化酶(superoxide dismutase, SOD)消耗,引起活性氧(reactive oxygen species, ROS)大量累積,最終導致血管內皮氧化損傷,進而推進CVD的發生發展[5]。1-棕櫚酰基-2-(5'-氧-戊酰基)-sn-甘油-3-磷酸膽堿[1-palmitoyl-2-(5-oxovaleroyl)-sn-glycero-3-phosphocholine, POVPC]是氧化低密度脂蛋白中參與AS發生發展的一種重要氧化磷脂,可激活內皮細胞,誘導炎癥反應,破壞內皮功能,因其生物活性穩定,被廣泛應用于CVD相關研究中[6-7]。核轉錄因子紅系2相關因子2/血紅素加氧酶-1(nuclear factor-erythroid 2-related factor 2/heme oxygenase-1, Nrf2/HO-1)是心血管系統中調控氧化應激反應的重要通路,具有抗氧化、抗炎、減少細胞凋亡和抑制血管鈣化等作用,能夠抑制ROS的產生,減少SOD的消耗,進而抑制氧化損傷,延緩CVD的發生發展[8-10]。課題組前期研究結果表明,補陽還五湯苷類組分具有減輕炎癥反應,保護血管內皮的作用[11-12]。黃芪是補陽還五湯發揮心腦血管效應的主要藥物,而黃芪甲苷(astragaloside Ⅳ, AST-Ⅳ)是黃芪的主要藥效物質[12]。同時,有研究證明AST-Ⅳ具有抗炎、抗氧化的功效[13-14]。因此,我們推測AST-Ⅳ也能發揮復方的相關作用,如抑制VEC氧化損傷,保護VEC。
本研究通過POVPC誘導主動脈血管內皮細胞(aortic vascular endothelium cell, AVEC)建立細胞氧化損傷模型,以Nrf2/HO-1信號通路為靶點,探究AST-Ⅳ對血管內皮氧化損傷的保護作用。
1 材料
1.1 動物
SPF級8周齡雄性SD大鼠2只,購自湖南斯萊克景達實驗動物有限公司,大鼠體質量200~250 g,飼養于湖南中醫藥大學SPF級屏障系統,使用標準動物飼料喂養,溫度為恒溫(25±2) ℃,室內明暗周期為12 h一循環。動物實驗過程均符合倫理學要求,動物生產許可證號:SCXK(湘)2019-0004。本研究通過湖南中醫藥大學動物實驗中心倫理審查(審批號:LL2022072703)。
1.2 主要試劑
AST-Ⅳ(成都埃法生物科技有限公司,批號:84687-43-4);DEM/F12、PBS緩沖液、胎牛血清(武漢普諾賽生命科技有限公司,批號:WHB823G041、WHB823P091、SA230608);ML385(美國MedChemexpress公司,批號:24435);POVPC、CCK-8檢測試劑盒(美國APE-BIO公司,批號:C360111346294、貨號:K1018);ROS測定試劑盒、羅丹明標記鬼筆環肽、DAPI溶液、抗熒光衰減封片劑、(北京索萊寶科技有限公司,批號:2307002、A426D011、2308001、20220224);結晶紫染色液(北京索萊寶科技有限公司,貨號:CA1610);總SOD測試盒(羥胺法)(南京建成生物有限公司,貨號:A000-1-1);Matrigel基質膠、Transwell小室(美國康寧公司,貨號:356234、3422);Nrf2(武漢三鷹生物技術有限公司,批號:16396-1-AP);HO-1(成都正能生物技術有限責任公司,貨號:R24541);兔抗血管性血友病因子(von willebrand factor, vWF)多克隆抗體(北京博奧森生物技術有限公司,批號:BS-0586R);FITC標記山羊抗兔IgG(H+L)(武漢三鷹生物技術有限公司,貨號:SA00003-2)。
1.3 主要儀器
CO2培養箱(美國賽默飛公司,型號:3427);超凈工作臺(蘇州安泰技術有限公司,型號:SW-CJ-1FD);倒置熒光顯微鏡、正置熒光顯微鏡(德國Zeiss公司,型號:Axio Vert.A1、Axio scope 5);全景掃描儀(匈利亞3D HISTECH公司,型號:Pannoramic MIDI);電泳槽、轉印槽、多功能酶標成像系統儀、Gel DocXR+凝膠成像系統(美國Bio-Rad公司,型號:1658001、1703930、Cytation3、Chemi DocTM XRS+)。
2 方法
2.1 AVEC的提取與鑒定
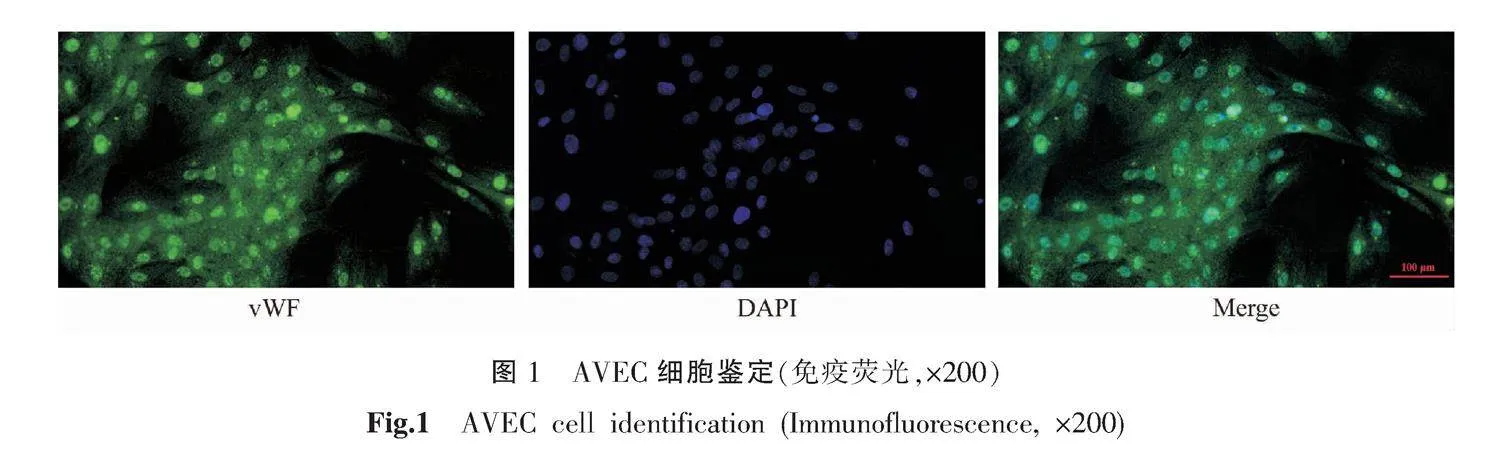
參照前期研究方法[15],選用SD大鼠,分離其胸主動脈,提取培養AVEC。將分離的AVEC使用DMEM/F12(含10%胎牛血清、1%鏈霉素、1%青霉素)完全培養基培養,并且使用免疫熒光法鑒定細胞表面標志物vWF的表達。將AVEC加入含玻璃片的24孔板中生長24 h,使用4%多聚甲醛將處于對數生長期的AVEC固定。PBS緩沖液清洗后加入vWF(1:200),室溫孵育2 h。待孵育結束后PBS清洗3次,加入熒光二抗(1∶200)室溫孵育1 h。爬片清洗后進行DAPI核染,抗熒光淬滅封片劑封片,熒光顯微鏡觀察。
2.2 AVEC氧化損傷模型建立、分組及給藥
選擇3~6代生長良好的AVEC進行實驗。根據以往研究結果[16],選擇POVPC 35 μmol/L構建內皮細胞氧化損傷模型。參照AST-Ⅳ毒性實驗選擇合適的作用濃度,ML385采用5 μmol/L[17]作為干預濃度。將對數生長期的AVEC隨機分為對照(Control)組、模型組(Model組,35 μmol/L POVPC)、AST-Ⅳ低劑量組(AST-L組,35 μmol/L POVPC+50 μmol/L AST-Ⅳ)、AST-Ⅳ高劑量組(AST-H組,35 μmol/L POVPC+100 μmol/L AST-Ⅳ)和AST-Ⅳ高劑量+ ML385組(AST-H+ML385組,35 μmol/L POVPC+100 μmol/L AST-Ⅳ+5 μmol/L ML385)。培養24 h后,收集細胞及其上清液用于后續實驗研究。
2.3 CCK-8法檢測AST-Ⅳ毒性與給藥后細胞增殖情況
將2~6代處于對數生長期的AVEC用胰酶消化,終止消化后接種于96孔板中,細胞密度約為1×106個/mL,每孔接種細胞懸液100 μL,置于37 ℃、5% CO2培養箱中培養24 h。細胞加藥處理,培養24 h后,每孔加入1% CCK-8溶液100 μL,放置于培養箱中避光培養30~60 min,酶標儀測定450 nm處的吸光度(A)。細胞存活率=(A實驗組-A空白組)/(A對照組-A空白組)×100%。
2.4 Transwell小室測定AVEC遷移能力
將對數生長期的AVEC用胰酶消化,接種于24孔板中培養24 h。AVEC加藥處理后,用胰酶消化。終止消化后800 r/min離心5 min(離心半徑為16 cm)。使用無血清培養基重懸,調整密度為1×106個/mL,取100 μL接種于小室上室。在24孔板下室中加入600 μL含15% FBS的完全培養基,放入培養箱中培養24 h。取出小室,棄去培養基,用棉簽輕輕擦拭上室。取新的24孔板加入600 μL 4%多聚甲醛固定液,將小室放入后固定20 min。棄固定液,0.2%結晶紫染色10 min,PBS清洗3次,風干后隨機選擇5個視野進行鏡檢。
2.5 Matrigel基質膠成管實驗測定AVEC血管形成功能
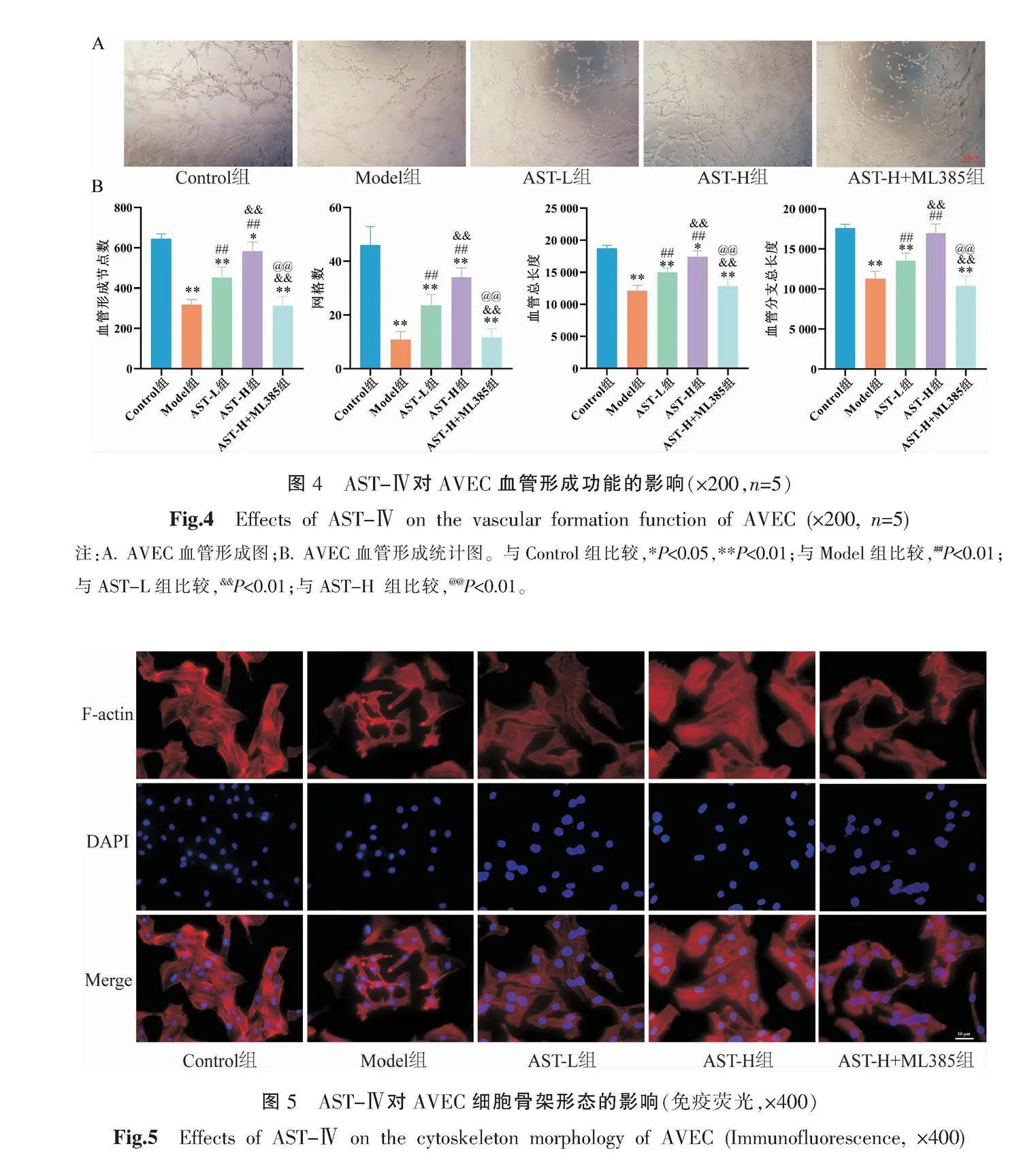
將Matrigel基質膠提前放置于4 ℃融化,加入至提前預冷的96孔板中,每孔50 μL。放置于培養箱中凝固。AVEC分組處理后,培養24 h,用胰酶消化。終止消化后800 r/min離心5 min(離心半徑為16 cm),使用無血清培養基重懸,調整密度為1×106個/mL。將細胞懸液加入配制好的Matrigel膠上(100 μL/孔,1×105個細胞),放入細胞培養箱中培養4~8 h,顯微鏡觀察、拍照。采用Image J進行圖像分析,每張圖像取相同放大倍數視野,計數血管形成的節點、網格數目、血管總長度、血管分支長度,并進行統計分析。
2.6 熒光顯微鏡觀察AVEC骨架形態
將AVEC接種于含細胞爬片的24孔板中,細胞密度約為1×105個/mL,每孔接種細胞懸液500 μL,放入培養箱培養24 h。AVEC加藥處理后,培養24 h,PBS清洗1次。加入4%多聚甲醛固定20 min。3%BSA(PBS配制)封閉30 min,加入配制好的鬼筆環肽(1∶500)300 μL(3% BSA配制),室溫孵育30 min。PBS 清洗3次,DAPI染核6 min,PBS清洗3次后,抗熒光衰減封片劑封片,熒光顯微鏡下觀察細胞骨架形態、纖維絲排列情況、結構是否斷裂缺失等。
2.7 DCFH-DA熒光探針檢測細胞ROS水平
將AVEC用胰酶消化,終止消化后接種于24孔板中,細胞密度約為1×105個/mL,每孔接種細胞懸液500 μL,放入培養箱培養24 h。AVEC加藥處理后,培養24 h,加入DCFH-DA探針應用液(300 μL/孔,1∶1 000)。置于培養箱中孵育30 min,避光,棄去培養液,無血清培養基清洗3次,熒光顯微鏡觀察檢測,采用Image J軟件,測定相同面積下的熒光強度,以平均熒光強度定量細胞內ROS水平。
2.8 試劑盒法檢測AVEC培養液中SOD水平
細胞加藥處理后,收集細胞培養上清液,按試劑盒說明書步驟配制試劑及應用液。將細胞培養上清液加入96孔板中,與相應試劑用旋渦混勻器充分混勻,置37 ℃恒溫水浴40 min。加入顯色液混勻,室溫放置10 min,酶標儀測定波長550 nm處的吸光度,計算上清液中SOD的含量。
2.9 RT-qPCR檢測Nrf2和HO-1 mRNA表達水平
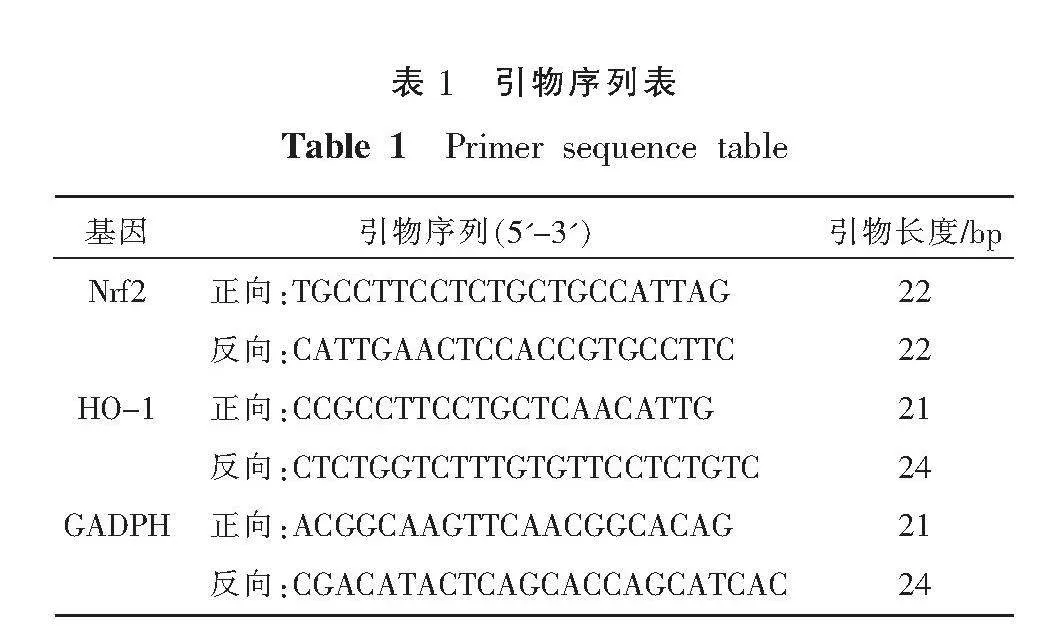
AVEC加藥處理后,培養24 h,使用總RNA試劑盒從細胞中提取總RNA,逆轉錄合成cDNA,隨后進行PCR擴增。反應條件按試劑盒的操作說明進行。內參使用GAPDH,數據分析采用2-ΔΔCt法。各基因及其內參的擴增引物序列由上海生工生物有限公司合成。詳見表1。
2.10 Western blot法檢測細胞內相關蛋白表達水平
收集處理后的細胞,用BCA法測定濃度后,加入上樣緩沖液,金屬浴15 min變性,儲存于4 ℃備用。配制10%的分離膠和5%的濃縮膠,濃縮膠恒壓60 V,當蛋白電泳至分離膠位置,更改電壓為80 V電泳,待溴酚藍電泳至下端2~5 mm時結束電泳,200 mA恒流轉膜一定時間。5%脫脂奶粉室溫封閉1 h,TBST洗3次,每次10 min。一抗 HO-1(1∶600)、Nrf2(1∶2 000)4 ℃孵育過夜。TBST洗3次,每次10 min。二抗(1∶8 000)37 ℃孵育1 h 后,再用TBST洗 3次,每次10 min。加入ECL發光液,使用數碼成像發光儀曝光顯影。采用Image J軟件分析條帶灰度。以β-actin作為內參,目的蛋白與內參蛋白的比值作為蛋白相對表達量。
2.11 統計學分析
實驗數據使用SPSS 23.0軟件進行統計,計量資料以“x±s”表示,若滿足正態性、方差齊性則采用單因素方差分析LSD法檢驗進行組間比較;方差不齊時,采用Dunnett T3檢驗。P<0.05表示差異具有統計學意義。
3 結果
3.1 大鼠AVEC鑒定
免疫熒光結果顯示,所提取的細胞可特異性表達vWF,顯示綠色熒光,鑒定為AVEC。詳見圖1。
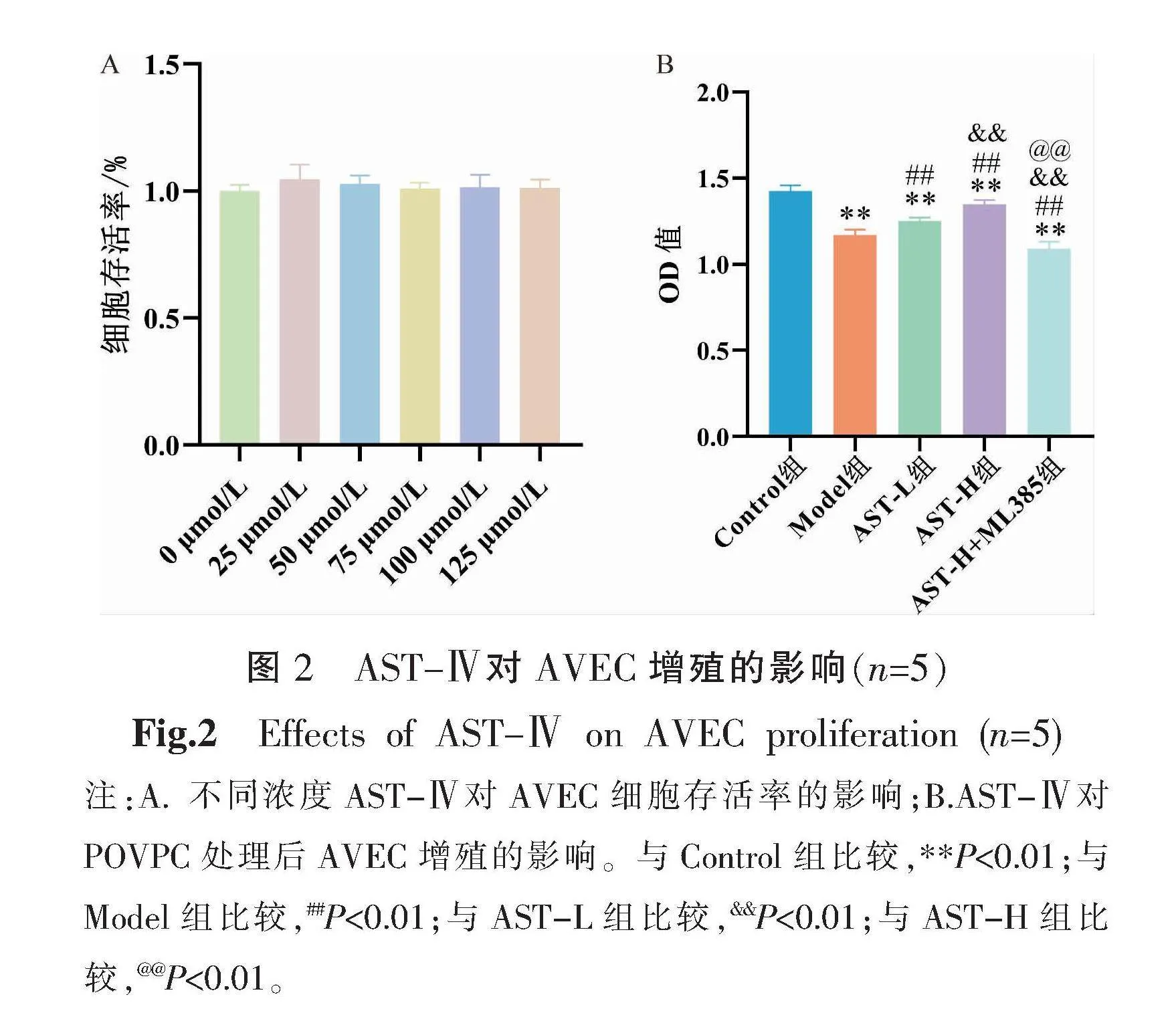
3.2 AST-Ⅳ對AVEC增殖的影響
與0 μmol/L AST-Ⅳ比較,梯度劑量的AST-Ⅳ干預AVEC,細胞存活率差異無統計學意義(P>0.05),提示AST-Ⅳ對內皮細胞無明顯毒性。因此,后期實驗選用50 μmol/L AST-Ⅳ干預濃度作為低劑量組,100 μmol/L AST-Ⅳ干預濃度作為高劑量組。與Control組相比,Model組AVEC增殖活力明顯下降(P<0.01);與Model組比較,AST-L組及AST-H組細胞活力顯著上升(P<0.01),且AST-H組細胞活力強于AST-L組(P<0.01);與AST-H組比較,AST-H+ML385組細胞活力明顯下降(P<0.01)。詳見圖2。
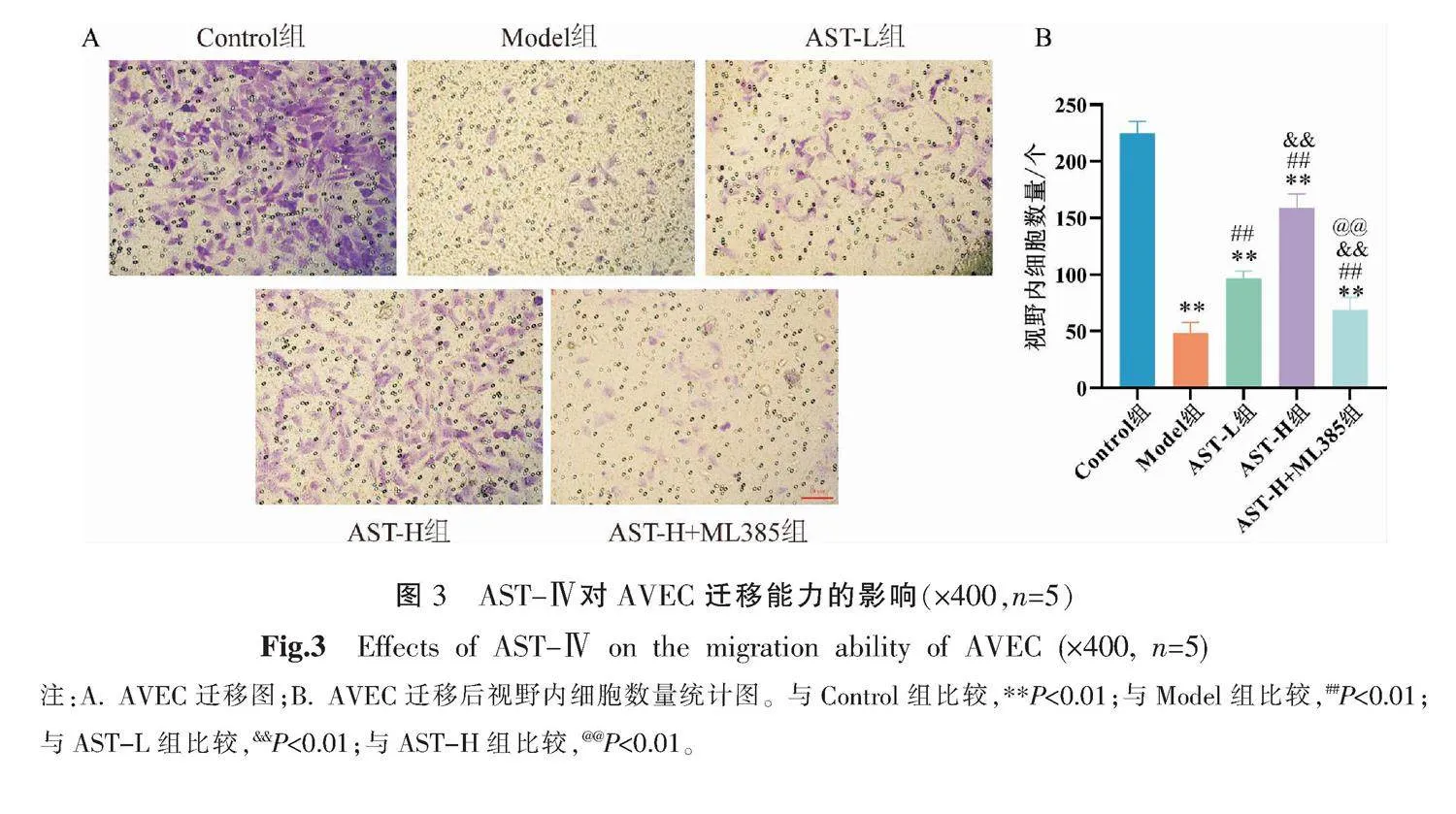
3.3 AST-Ⅳ對AVEC遷移能力的影響
與Control組比較,Model組細胞遷移數量明顯減少(P<0.01);與Model組比較,AST-L組及AST-H組細胞遷移數量明顯增多(P<0.01),且AST-H組細胞遷移數量高于AST-L組(P<0.01);與AST-H組比較,AST-H+ML385組細胞遷移數量明顯下降(P<0.01)。詳見圖3。
3.4 AST-Ⅳ對AVEC血管形成功能的影響
與Control組比較,Model組血管形成的節點數和血管形成個數既網格數顯著降低(P<0.01),血管形成的血管總長度和血管分支長度顯著縮短(P<0.01);與Model組比較,AST-L組及AST-H組血管形成的節點數和網格數顯著升高(P<0.01),血管總長度和血管分支總長度顯著增長(P<0.01),且AST-H組血管形成的節點數、網格數高于AST-L組(P<0.01),血管總長度、血管分支總長度長于AST-L組(P<0.01);與AST-H組比較,AST-H+ML385組AVEC血管形成的節點數和網格數下降(P<0.01),血管總長度和血管分支長度顯著縮短(P<0.01)。詳見圖4。
3.5 AST-Ⅳ對AVEC細胞骨架形態的影響
經鬼筆環肽染色后,細胞骨架F-肌動蛋白(F-actin)表現為紅色熒光。Control組細胞內骨架微絲排列均勻整齊,細胞間連接完整,無明顯斷裂。與Control組比較,Model組內皮細胞內骨架微絲排列紊亂,纖維絲出現交叉或斷裂,細胞間連接不完整,細胞骨架破壞;與Model組比較,AST-L組及AST-H組細胞骨架修復,細胞間連接較完整,骨架微絲分布整齊,且以AST-H組骨架修復更為完善;與AST-H組比較,AST-H+ML385組對于細胞骨架的修復作用被減弱。詳見圖5。
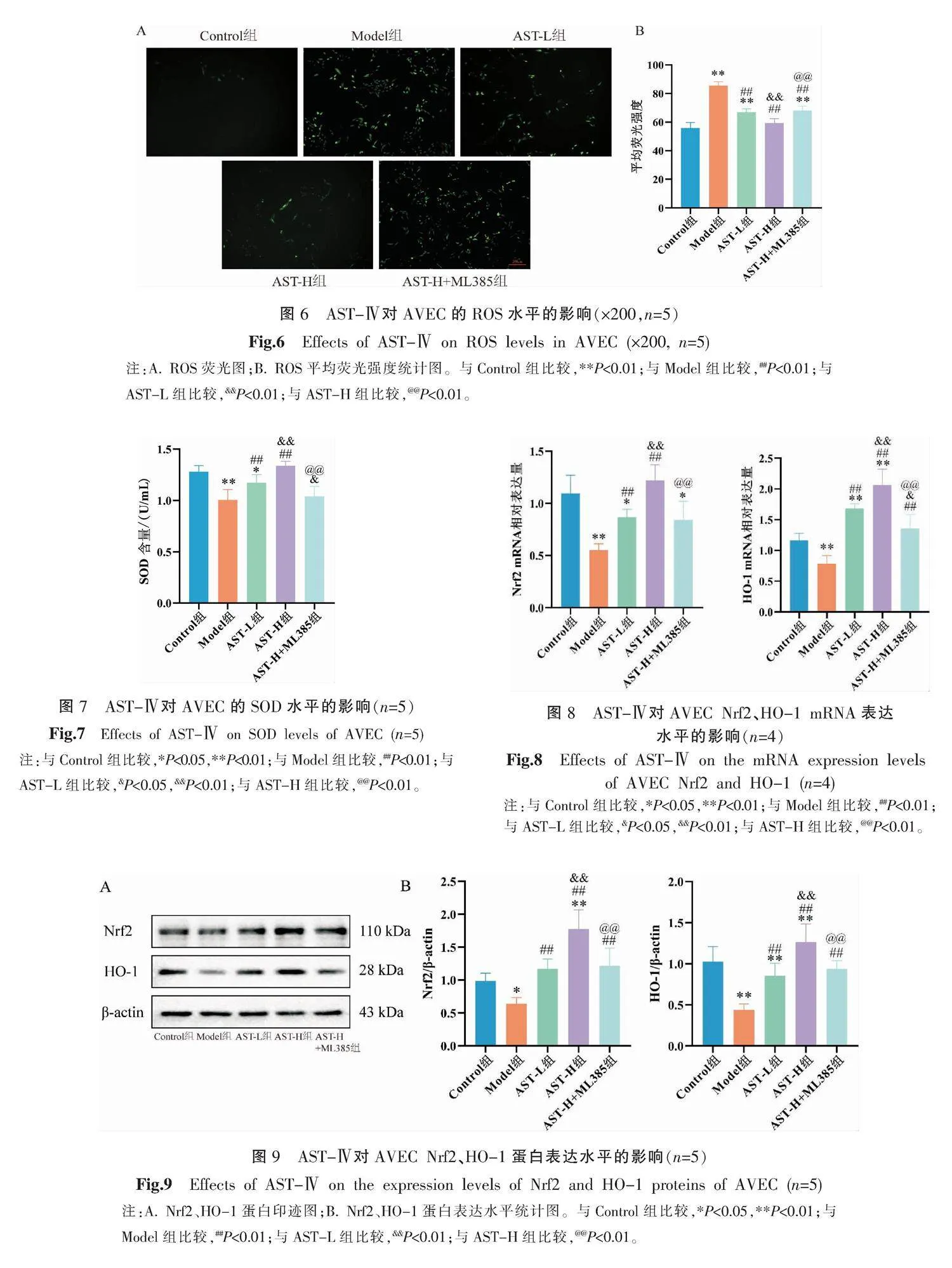
3.6 AST-Ⅳ對AVEC ROS水平的影響
與Control組比較,Model組細胞內ROS熒光強度顯著增加(P<0.01);與Model組比較,AST-L組及AST-H組細胞內ROS熒光強度顯著下降(P<0.01),且AST-H組細胞內ROS水平低于AST-L組(P<0.01);與AST-H組比較,AST-H+ML385組細胞內ROS水平升高(P<0.01)。詳見圖6。
3.7 AST-Ⅳ對AVEC SOD水平的影響
與Control組比較,Model組SOD含量顯著降低(P<0.01);與Model組比較,AST-L組及AST-H組SOD含量顯著升高(P<0.01),且AST-H組SOD水平高于AST-L組(P<0.01);與AST-H組比較,AST-H+ML385組SOD水平顯著降低(P<0.01)。詳見圖7。
3.8 AST-Ⅳ對AVEC Nrf2、HO-1 mRNA表達的影響
與Control組比較,Model組細胞內Nrf2、HO-1 mRNA表達水平顯著降低(P<0.01);與Model組比較,AST-L組及AST-H組細胞內Nrf2、HO-1 mRNA表達水平顯著升高(P<0.01),且AST-H組Nrf2、HO-1 mRNA表達水平高于AST-L組(P<0.01);與AST-H組比較,AST-H+ML385組Nrf2、HO-1 mRNA表達水平顯著降低(P<0.01)。詳見圖8。
3.9 AST-Ⅳ對AVEC Nrf2、HO-1蛋白表達水平的影響
與Control組比較,Model組細胞內Nrf2和HO-1蛋白表達水平顯著降低(P<0.05或P<0.01);與Model組比較,AST-L組及AST-H組細胞內Nrf2和HO-1蛋白表達水平顯著升高(P<0.01),且AST-H組Nrf2和HO-1蛋白表達水平高于AST-L組(P<0.01);與AST-H組比較,AST-H+ML385組Nrf2和HO-1蛋白表達水平顯著降低(P<0.01)。詳見圖9。
4 討論
CVD是常見的嚴重威脅生命健康的疾病,具有高發病率、高死亡率、高致殘率[18-20]。AS作為CVD的病理基礎,主要表現為血管內脂質浸潤、血管平滑肌細胞增殖及表型轉化、斑塊形成或破裂鈣化、血栓形成等病理過程,最終導致血管功能發生障礙[20-22]。引起AS發生的病理機制十分復雜,其中,氧化應激在疾病發展過程中可誘導VEC損傷。一方面,血管完整性破壞引起單核細胞募集,血小板黏附聚集,引起局部血栓形成[23];另一方面,受損的VEC分泌大量生物因子,引起平滑肌細胞增殖,細胞外基質大量分泌,加速AS的進展[24]。因此,抑制氧化損傷,保護VEC,是防治AS的重要手段之一。
中醫學認為,氣虛血瘀是AS臨床最常見的病機之一,益氣活血法是治療AS的基本治療法則之一[25]。益氣活血法主方補陽還五湯對于AS發展階段的脂質沉積、炎癥浸潤、平滑肌細胞增殖、巨噬細胞表型轉化均具有顯著的抑制作用[25-26]。AST-Ⅳ是補陽還五湯中的主要苷類物質,也是中藥黃芪的提取物,可抑制CVD發展中的氧化應激、減少炎癥浸潤與細胞凋亡,但其對于VEC保護、抗氧化應激的作用機制尚不明確[27-29]。
本研究提取了原代細胞,免疫熒光染色結果顯示所提取的細胞可特異性表達vWF,鑒定為AVEC。為明確高濃度的AST-Ⅳ對于AVEC是否有毒性作用,采用CCK-8檢測梯度濃度的AST-Ⅳ對AVEC細胞活力的影響,因此選定AST-Ⅳ的干預劑量。利用POVPC建立AVEC氧化損傷模型,AVEC增殖、遷移、血管形成功能顯著下降,骨架結構顯著破壞,提示POCPC破壞了VEC的形態結構和基本功能。ROS過度產生及清除不足是引起機體氧化、抗氧化功能紊亂的主要原因之一,大量ROS的蓄積可改變組織細胞中蛋白質的結構,損害其功能[30]。SOD是一種清除氧自由基的酶,可通過酶促反應催化氧自由基降解,保護組織細胞,維持機體氧化與抗氧化平衡[31]。POVPC作用于AVEC后,氧化應激產物ROS水平顯著上升,抗氧化物質SOD水平顯著下降,提示POVPC可誘導氧化應激的產生;AST-Ⅳ作用后,AVEC細胞活力、遷移和血管形成功能顯著升高,同時伴隨細胞骨架的修復,氧化應激產物ROS水平降低及抗氧化物質SOD水平升高,提示AST-Ⅳ具有抗氧化應激、保護AVEC的藥理作用,且具有一定的劑量依賴性。Nrf2是調節細胞氧化應激反應的關鍵因子,當細胞受到氧化應激及其他病理因素刺激時,細胞啟動轉錄過程,激活一系列抗氧化酶,清除氧化損傷產物ROS等[31-32]。HO-1是受Nrf2調控的下游抗氧化酶之一,其本身及誘導產生的內源性保護物質也可發揮抗炎、抗氧化的作用。因此,Nrf2/HO-1信號通路激活也將減輕氧化應激產物對于VEC的損傷,延緩AS的進展[33-35]。ML385是Nrf2的抑制劑,可與Nrf2結合并抑制其下游靶基因表達[36]。在AST-Ⅳ高劑量作用下,應用ML385減輕了AST-Ⅳ對AVEC細胞活力、遷移、血管形成功能的保護作用,抑制了細胞結構的恢復,同時減輕了AST-Ⅳ對于AVEC的抗氧化應激作用。Nrf2、HO-1
mRNA及蛋白的檢測結果顯示,POVPC可引起AVEC出現Nrf2/HO-1通路抑制,而AST-cKT0LlsG9vkkUdck1aQXDA==Ⅳ干預后,激活Nrf2/HO-1信號通路,誘導AVEC保護作用,并且AST-H的通路激活作用更為顯著;但ML385干預后,AST-H對于Nrf2/HO-1的激活作用也被部分逆轉,說明AST-Ⅳ可能通過激活Nrf2/HO-1信號通路抑制氧化損傷,保護血管內皮。
綜上所述,AST-Ⅳ可通過激活Nrf2/HO-1信號通路,增強AVEC增殖、遷移、血管形成功能,促進細胞結構的修復,增強抗氧化能力,對POVPC誘導的AVEC損傷有一定的抑制作用,提示AST-Ⅳ可能成為抗AS的有效中藥單體成分。本研究可為靶向中醫藥復方單體成分及精準醫療提供一定的研究基礎。
參考文獻
[1] 馬麗媛, 王增武, 樊 靜, 等. 《中國心血管健康與疾病報告2022》要點解讀[J]. 中國全科醫學, 2023, 26(32): 3975-3994.
[2] NOALE M, LIMONGI F, MAGGI S. Epidemiology of cardiovascular diseases in the elderly[J]. Advances in Experimental Medicine and Biology, 2020, 1216: 29-38.
[3] PICH?魪 M E, TCHERNOF A, DESPR?魪S J P. Obesity phenotypes, diabetes, and cardiovascular diseases[J]. Circulation Research, 2020, 126(11): 1477-1500.
[4] GUIEU R, RUF J, MOTTOLA G. Hyperhomocysteinemia and cardiovascular diseases[J]. Annales De Biologie Clinique, 2022, 80(1): 7-14.
[5] 劉彩霞, 鄧常清. 中藥及復方改善血管內皮細胞氧化應激損傷的研究進展[J]. 湖南中醫藥大學學報, 2021, 41(1): 151-158.
[6] LI Y, ZHANG Y X, NING D S, et al. Simvastatin inhibits POVPC-mediated induction of endothelial-to-mesenchymal cell transition[J]. Journal of Lipid Research, 2021, 62: 100066.
[7] HAN Z Y, HU H D, YIN M Z, et al. HOXA1 participates in VSMC-to-macrophage-like cell transformation via regulation of NF-κB p65 and KLF4: A potential mechanism of atherosclerosis pathogenesis[J]. Molecular Medicine, 2023, 29(1): 104.
[8] ZHANG Q, LIU J, DUAN H, et al. Activation of Nrf2/HO-1 signaling: An important molecular mechanism of herbal medicine in the treatment of atherosclerosis via the protection of vascular endothelial cells from oxidative stress[J]. Journal of Advanced Research, 2021, 34: 43-63.
[9] MANSOURI A, REINER ?譕, RUSCICA M, et al. Antioxidant effects of statins by modulating Nrf2 and Nrf2/HO-1 signaling in different diseases[J]. Journal of Clinical Medicine, 2022, 11(5): 1313.
[10] ZHANG Q, WANG L, WANG S Q, et al. Signaling pathways and targeted therapy for myocardial infarction[J]. Signal Transduction and Targeted Therapy, 2022, 7: 78.
[11] 傅馨瑩, 孫正驥, 龍清吟, 等. 補陽還五湯苷類組分介導NF-κB通路抗動脈粥樣硬化炎癥反應的機制研究[J]. 中國中藥雜志, 2023, 48(1): 202-210.
[12] FU X Y, SUN Z J, LONG Q Y, et al. Glycosides from Buyang Huanwu Decoction inhibit atherosclerotic inflammation via JAK/STAT signaling pathway[J]. Phytomedicine: International Journal of Phytotherapy and Phytopharmacology, 2022, 105: 154385.
[13] WANG Z X, WU Y C, PEI C X, et al. Astragaloside Ⅳ pre-trea?鄄
tment attenuates PM2.5-induced lung injury in rats: Impact on autophagy, apoptosis and inflammation[J]. Phytomedicine: Intern?鄄ational Journal of Phytotherapy and Phytopharmacology, 2022, 96: 153912.
[14] ZHANG X Q, QU H Y, YANG T, et al. Astragaloside Ⅳ attenuate MI-induced myocardial fibrosis and cardiac remodeling by inhibiting ROS/caspase-1/GSDMD signaling pathway[J]. Cell Cycle, 2022, 21(21): 2309-2322.
[15] YAN F C, DING H, SUN Z J, et al. Glycoside combinations of Buyang Huanwu Decoction ameliorate atherosclerosis via STAT3, HIF-1, and VEGF[J]. Naunyn-Schmiedeberg's Archives of Pharmacology, 2023, 396(6): 1187-1203.
[16] 馬 露, 楊 雷, 鄧常清, 等. 基于PI3K/Akt通路研究內皮祖細胞來源外泌體協同丹參酮ⅡA對血管內皮細胞氧化損傷的保護作用[J]. 中國中藥雜志, 2023, 48(23): 6423-6433.
[17] JI L L, MOGHAL N, ZOU X R, et al. The NRF2 antagonist ML385 inhibits PI3K-mTOR signaling and growth of lung squamous cell carcinoma cells[J]. Cancer Medicine, 2023, 12(5): 5688-5702.
[18] ZHAO D, LIU J, WANG M, et al. Epidemiology of cardiovascular disease in China: Current features and implications[J]. Nature Reviews Cardiology, 2019, 16(4): 203-212.
[19] VALK R, HAMMILL J, GRIP J. Saturated fat: Villain and bogeyman in the development of cardiovascular disease?[J]. European Journal of Preventive Cardiology, 2022, 29(18): 2312-2321.
[20] YI H J, LEE D H, HONG B Y, et al. The long-term functional effect of thrombectomy on patients with middle cerebral artery occlusion who exhibit moderate to severe disability[J]. Medicina, 2021, 57(5): 509.
[21] TANG H Y, CHEN A Q, ZHANG H, et al. Vascular smooth muscle cells phenotypic switching in cardiovascular diseases[J]. Cells, 2022, 11(24): 4060.
[22] FAN J L, WATANABE T. Atherosclerosis: Known and unknown[J]. Pathology International, 2022, 72(3): 151-160.
[23] TABARES-GUEVARA J H, VILLA-PULGARIN J A, HERNANDEZ J C. Atherosclerosis: Immunopathogenesis and strategies for immunotherapy[J]. Immunotherapy, 2021, 13(14): 1231-1244.
[24] 林云釵, 王 航, 周 強. LINC00707靶向miR-30c-5p對ox-LDL誘導的人血管平滑肌細胞增殖、遷移及炎癥因子的影響[J]. 中國藥理學通報, 2023, 39(10): 1829-1835.
[25] 潘 楊, 周明學, 郭家娟. 益氣活血中藥防治動脈粥樣硬化的研究[J]. 中國中醫基礎醫學雜志, 2021, 27(2): 362-366.
[26] 李菀榆, 龍清吟, 傅馨瑩, 等. 補陽還五湯和黃芪當歸配伍對動脈粥樣硬化小鼠炎癥反應的影響[J]. 中國中藥雜志, 2023, 48(15): 4164-4172.
[27] 張紅珍, 劉建春, 王 青, 等. 補陽還五湯含藥鼠血清對血管平滑肌細胞Rho 激酶及TFmRNA、基質金屬蛋白酶MMP-2mRNA、MMP-9mRNA表達的影響[J]. 中華中醫藥學刊, 2021, 39(1): 17-21.
[28] LENG B, ZHANG Y J, LIU X R, et al. Astragaloside Ⅳ suppresses high glucose-induced NLRP3 inflammasome activation by inhibiting TLR4/NF-κB and CaSR[J]. Mediators of Inflammation, 2019, 2019: 1082497.
[29] CHENG S Y, ZHANG X X, FENG Q, et al. Astragaloside Ⅳ exerts angiogenesis and cardioprotection after myocardial infarction via regulating PTEN/PI3K/Akt signaling pathway[J]. Life Sciences, 2019, 227: 82-93.
[30] HEUSCH G, ANDREADOU I, BELL R, et al. Health position paper and redox perspectives on reactive oxygen species as signals and targets of cardioprotection[J]. Redox Biology, 2023, 67: 102894.
[31] 徐 暢, 宋藝蘭, 姜京植, 等. 川芎嗪通過AMPK/NF-κB和Nrf-2/HO-1途徑減輕過敏性氣道炎癥和氧化應激的實驗研究[J]. 免疫學雜志, 2021, 37(2): 100-106.
[32] SHEN Y M, LIU X J, SHI J H, et al. Involvement of Nrf2 in myocardial ischemia and reperfusion injury[J]. International Journal of Biological Macromolecules, 2019, 125: 496-502.
[33] HE F, RU X L, WEN T. NRF2, a transcription factor for stress response and beyond[J]. International Journal of Molecular Sciences, 2020, 21(13): 4777.
[34] CHEN X H, WAN W G, GUO Y, et al. Pinocembrin ameliorates post-infarct heart failure through activation of Nrf2/HO-1 signaling pathway[J]. Molecular Medicine, 2021, 27(1): 100.
[35] WANG Y R, YANG C X, ELSHEIKH N A H, et al. HO-1 reduces heat stress-induced apoptosis in bovine granulosa cells by suppressing oxidative stress[J]. Aging, 2019, 11(15): 5535-5547.
[36] XIAN P P, HEI Y, WANG R, et al. Mesenchymal stem cell-derived exosomes as a nanotherapeutic agent for amelioration of inflammation-induced astrocyte alterations in mice[J]. Theranostics, 2019, 9(20): 5956-5975.
本文引用: 譚 維, 傅馨瑩, 楊仁義, 馬 露, 丁 煌, 劉曉丹, 張 偉. 黃芪甲苷調控Nrf2/HO-1信號通路對血管內皮細胞氧化損傷的影響[J]. 湖南中醫藥大學學報, 2024, 44(9): 1592-1600.
〔收稿日期〕2023-11-14
〔基金項目〕國家自然科學基金項目(82174218);湖南省科技創新計劃資助項目(2023SK2021);湖南省科技創新領軍人才項目(2023RC1066)。
〔通信作者〕*張 偉,男,博士,教授,博士研究生導師,E-mail:zhangwei1979@hnucm.edu.cn。
