產金屬β-內酰胺酶豬源ST201型多殺性巴氏桿菌的耐藥性和致病性分析
2024-12-31 00:00:00楊榮榮張婷唐萍萍何爽方趙苗苗雷連成張付賢
畜牧獸醫學報 2024年12期



摘" 要: 為探究湖北荊州某生豬養殖企業育肥豬呼吸道疾病的致病原,采集病豬肺組織進行病原的篩查和分離鑒定。本研究對分離菌株進行形態學觀察、生理生化、16S rDNA基因序列測定、血清型分型、多位點序列分析、藥物敏感性試驗、耐藥基因測序分析以及動物致病性試驗。結果顯示,分離菌株Pm0525為革蘭陰性短桿菌;生化試驗、PCR和16S rDNA測序鑒定為多殺性巴氏桿菌。該菌為莢膜血清型A型,分子分型為ST201型,與ST1615型聚為一支;分離菌株對慶大霉素、卡那霉素、麥迪霉素等13種抗生素敏感,對氯霉素、克林霉素、頭孢呋辛、頭孢他啶和四環素耐藥;分離菌株具有中等生物被膜形成能力,攜帶SulⅠ、SulⅡ、tetA和bla-TEM四種耐藥基因,可通過產金屬 β-內酰胺酶(MBL)方式發揮抗β-內酰胺類抗生素的作用。中藥藥敏試驗結果顯示,該分離菌株對中藥黃連呈現明顯的敏感性。該分離株攜帶18種毒力基因,并對小鼠具有較強的致病性。本研究系統性分析了豬源多殺性巴氏桿菌的血清型、分子分型、藥物敏感性和致病性,為進一步研究多殺性巴氏桿菌感染的致病機制、防控和精準施治提供科學依據和數據參考。
關鍵詞: 多殺性巴氏桿菌;金屬 β-內酰胺酶;MLST分析;耐藥性;致病性
中圖分類號:S852.61
文獻標志碼:A
文章編號:0366-6964(2024)12-5692-14
doi: 10.11843/j.issn.0366-6964.2024.12.032
開放科學(資源服務)標識碼(OSID):
收稿日期:2024-01-26
基金項目:國家科技部“十四五”重點研發項目(2021YFD1800405);國家自然科學基金面上項目(32072823)
作者簡介:楊榮榮(2001-),女,陜西寶雞人,碩士生,主要從事多殺性巴氏桿菌相關研究, E-mail:yangrr0702@163.com
*通信作者:張付賢,主要從事獸醫微生物與免疫學研究, E-mail:zhangfuxian99@163.com
Analysis of Drug Resistance and Pathogenicity of Metallo-β-Lactamase-producing
Porcine ST201 Pasteurella multocida
YANG Rongrong1, ZHANG" Ting1, TANG Pingping2, HE Shuangfang3, ZHAO Miaomiao1, LEI Liancheng1,4, ZHANG" Fuxian1*
(1.College of Animal Science and Technology, Yangtze University, Jingzhou 434023," China;
2.Qianjiang Bureau of Agriculture and Rural Affairs, Qianjiang 433199," China;
3.Zhangjin Town for Animal Husbandry and Veterinary Technical Service, Qianjiang 433140," China;
4.College of Veterinary Medicine, Jilin University, Changchun 130062," China)
Abstract:" The study was design to investigate the pathogenic agents of respiratory tract diseases of fattening pigs in a pig breeding enterprise in Jingzhou, Hubei Province. The pathogen was screened and isolated from the diseased pig lung tissue. Morphological observation, physiological and biochemical analysis, 16S rDNA gene sequence, serotype typing, multilocus sequence typing, drug sensitivity test, drug resistance gene sequencing analysis and animal pathogenicity test were performed for the isolated strains. The results showed that the isolated strain Pm0525 was Gram-negative brevibacterium. It was identified as Pasteurella multocida by biochemical assay, PCR and 16S rDNA sequence analysis. The isolated strain was identified as Pasteurella multocida type A. Multilocus sequence typing analysis confirmed that the isolated strain is type ST201 and merged with type ST1615. The isolated strain was sensitive 13 kinds of antibiotics, such as gentamicin, kanamycin and medemycin, and resistant to chloramphenicol, clindamycin, cefuroxime, ceftazidime and tetracycline. The isolated strain showed moderate biofilm formation ability, carrying out four resistance genes, SulⅠ, SulⅡ, tetA and bla-TEM. The isolated strain may play the role of anti-β-lactam antibiotics by producing metal-beta-lactamase (MBL). In addition, traditional Chinese medicine Coptis had obvious inhibitory effect on the isolated strain in vitro. The isolated strain carried 18 virulence genes, and showed strong pathogenicity to mice. This study systematically analyzed the serotype, molecular typing, drug sensitivity and pathogenicity of swine Pasteurella multocida, providing scientific basis and data reference for further research on the transmission, prevention and precise treatment of Pasteurella infection.
Key words: Pasteurella multocida; metallo-beta-lactamase; MLST analysis; drug resistance; pathogenicity
*Corresponding author:" ZHANG Fuxian,E-mail:zhangfuxian99@163.com
多殺性巴氏桿菌(Pasteurella multocida,Pm)作為一種重要的人獸共患革蘭陰性病原菌,廣泛存在于全世界大部分地區,可引起包括人在內的多種動物的呼吸道疾病和出血性疾病[1-2]。Pm感染造成的損失和危害仍然是困擾公共安全和畜禽養殖業的主要問題之一。多殺性巴氏桿菌莢膜血清型眾多,不同血清型之間的交叉保護力差異顯著,且不同血清型引起動物的臨床癥狀也不盡相同。Pm罕見的莢膜型的出現以及可能存在莢膜型的轉變和跨種傳播問題,引發廣泛關注[3]。多殺性巴氏桿菌病的病型、致病性、宿主特異性和免疫性等,均與其血清型密切相關。Pm通常依據菌株攜帶毒力因子的血清學反應差異進行分型。根據Pm莢膜抗原與血清發生的被動凝集試驗,可分為A、B、D、E和F 5種莢膜血清型[4-5];基于Pm脂多糖的差異性,利用瓊擴試驗可將Pm分為16種血清型[6-7];多位點序列分型(multilocus sequence typing,MLST)是近年來發展很快的分子生物學分析方法,也常被用于Pm的分子分型和溯源分析[8]。近年來由于抗生素的濫用,造成臨床致病菌對抗生素的耐藥情況日趨嚴重,而產超廣譜β-內酰胺酶(extended-spectrum β-lactamases,ESBLs)和金屬β-內酰胺酶(metallo-β-lactamases,MBLs)細菌一直是臨床上公認的多重耐藥病原體。其中產ESBLs和MBLs是導致細菌耐β-內酰胺類抗菌藥物的主要機制之一,使細菌對大多數β-內酰胺類抗生素產生耐藥性。Pm菌株的流行病學調查、溯源、血清型和耐藥性分析等生物學特性研究是探索Pm感染多個宿主致病機制的有效途徑。
豬是巴氏桿菌病的自然宿主,感染后引起豬巴氏桿菌肺炎和傳染性萎縮性鼻炎等急性流行性或散發性和繼發性傳染病,危害嚴重[9-10]。本研究從患呼吸道疾病豬肺組織中分離到一株Pm,經鑒定為莢膜A型多殺性巴氏桿菌,MLST為ST201型。對分離菌株進行致病性試驗、耐藥基因測序分析和抗生素、中藥敏感性試驗,以期為豬源多殺性巴氏桿菌的溯源、流行病學調查和巴氏桿菌病的防控以及藥物精準施治提供參考依據。
1" 材料與方法
1.1" 病料來源
湖北省荊州市某生豬養殖企業育肥豬出現減食、體溫升高、咳嗽、鼻孔有黏性分泌物和呼吸困難等臨床癥狀,抗生素治療后病情好轉,但時有反復。剖檢病死豬的肺、淋巴結和鼻咽拭子進行病原的篩查以及細菌的分離培養。
1.2" 培養基與試劑
TSA和TSB購買自青島海博生物科技有限公司;革蘭染色劑和四季青新生牛血清購自北京索萊寶科技有限公司;藥敏紙片和細菌生化微量鑒定管購買自杭州微生物試劑有限公司;細菌基因組提取試劑盒,病毒基因組提取試劑盒,購自北京天根生物科技有限公司,Taq酶,反轉錄試劑購自北京聚合美生物科技有限公司。
1.3" 實驗動物和供試菌株
SPF級昆明雌性小鼠50只,6~8周齡,購自湖北實驗動物中心。豬圓環病毒2型(porcine circovirus type 2,PCV2)、豬繁殖與呼吸綜合征病毒(porcine reproductive and respiratory syndrome virus,PRRSV)、豬鏈球菌2型(Streptococcus suis type 2,SS2)、多殺性巴氏桿菌(Pasteurella multocida,Pm)、副豬嗜血桿菌(Haemophilus parasuis,HPS)和肺炎克雷伯菌(Klebsiella pneumoniae,Kp)的陽性毒株和陽性菌株由長江大學動物重要病原分子生物學實驗室保存。
1.4" 豬呼吸道疾病病原檢測
取病死豬的肺、淋巴結等病變組織研磨勻漿后用300 μL無菌PBS稀釋,轉移至1.5 mL離心管中,按照組織基因組提取試劑盒操作說明書提取病料的DNA和RNA,RNA以M5 Super plus qPCR RT kit with gDNA remover試劑盒進行反轉錄。合成豬呼吸道疾病病原引物[11-13],以提取的DNA與反轉錄的cDNA為模板,通過PCR方法分別檢測PCV2、PRRSV、SS2、Pm、HPS和Kp的特異性基因,PCR法檢測患病豬的致病原。
1.5" 病原菌的分離及純化培養和形態學生理生化鑒定
取病原檢測為陽性的豬肺病料,在超凈工作臺中無菌條件下劃開肺組織,接種環蘸取組織并劃線接種于TSA固體平板,37 ℃恒溫培養箱內培養14 h;選取單菌落接種于含5%血清的TSB培養基,37 ℃純化培養12 h。觀察TSA平板上單菌落的形態,挑取疑似巴氏桿菌典型菌落,均勻涂布于載玻片上,革蘭染色后鏡檢觀察。分離菌株的純培養物接種至含5%血清的TSB液體培養基,培養至對數生長期,制樣用于掃描電鏡觀察。
取分離菌株的純培養物接種至含5%血清的TSB液體培養基,37 ℃、180 r·min-1培養12 h,接種細菌微量生化管,按照細菌生化微量鑒定管說明書進行結果判定。
1.6" PCR鑒定16S rDNA基因序列鑒定
合成多殺性巴氏桿菌kmt1基因的引物(F:5′-ATCCGCTATTTACCCAGTGG-3′,R:5′-GCTGTAAACGAACTCGCCAC-3′),以細菌基因組提取試劑盒提取的分離菌株DNA為模板進行PCR擴增[14];反應體系(20 μL):2×Taq PCR Master Mix酶 10 μL,上下游引物(10 μmol·L-1)各1 μL,ddH2O 7 μL,模板1 μL;PCR擴增后以1.2%凝膠對PCR產物進行電泳檢測。
合成細菌16S rDNA基因通用引物27F:5′-AGAGTTTGATCCTGGCTCAG-3′與1492R:5′-TACGCTACCTTGTTACGACTT-3′,PCR反應體系為20 μL:2×PCR mix 10 μL,上、下游引物各1 μL,dd H2O 7 μL,模板1 μL。PCR擴增產物送上海生工生物工程技術服務有限公司進行測序,使用NCBI BLAST對測序結果進行分析,使用MEGA軟件來構建系統進化樹。
1.7" 血清型鑒定
以提取的分離菌株DNA為模板,利用多殺性巴氏桿菌的莢膜分型引物(表1)對分離菌株進行血清型分析,PCR產物進行測序,確定分離菌株的血清型[14]。PCR反應體系為20 μL:2×PCR mix 10 μL,上、下游引物1 μL,ddH2O 7 μL,模板1 μL。PCR反應條件:94 ℃預變性5 min;94 ℃變性30 s,退火時間30 s,退火溫度55 ℃,72 ℃延伸30 s,循環35次,72 ℃延伸3 min。PCR擴增后以1.2%凝膠對PCR產物進行電泳檢測。PCR擴增產物送上海生工生物工程技術服務有限公司進行測序,測序結果利用NCBI BLAST工具進行比對分析。
1.8" 多位點序列分析(Multi-locus sequence analysis,MLST)
根據MLST官網提供的多殺性巴氏桿菌信息,合成7個管家基因(adk、aroA、deoD、gdhA、g6pd、mdh和pgi)的引物,以分離菌株提取的DNA為模板進行PCR擴增[8];PCR反應體系為20 μL:2×PCR Mix 10 μL,上、下游引物1 μL,ddH2O 7 μL,模板1 μL;擴增后的產物電泳并切下條帶送去測序,最后使用MLST網絡數據庫來獲取等位基因信息,使用eBURST程序分析繪圖。
1.9" 藥敏試驗
1.9.1" 耐藥基因檢測
參考文獻合成18種常見耐藥基因的引物[15],以提取的分離菌株的基因組為模板進行PCR擴增,篩查分離菌攜帶耐藥基因的情況。
1.9.2" 生物被膜形成能力檢測
采用96孔板結晶紫染色法分析分離菌株的生物被膜形成能力。取分離菌株的純培養物按1∶100的比例接種于含5%血清的TSB培養基,取200 μL的菌液加入到96孔板內,37 ℃培養24 h;棄去菌液,以無菌PBS緩沖液清洗3次,加入100 μL甲醇固定15 min;棄去甲醇,清洗、烘干后,結晶紫染色5 min;洗滌后,加入100 μL 33%(體積比)的冰醋酸。以液體培養基為對照,用酶標儀測定每孔的OD590 nm值,作為空白對照ODc(ODc為對照孔的平均值);試驗重復三次。判定標準:ODclt;OD≤2ODc,為弱成膜能力;2ODclt;OD≤4ODc,為中等成膜能力;ODgt;4 ODc,則判定被測菌株具有強的生物被膜形成能力。
1.9.3" 抗生素敏感性試驗
采用Kirby-Bauer(K-B)紙片擴散法,測量抑菌圈直徑(inhibition zone diameter,IZD),參照CLSI標準分析多殺性巴氏桿菌分離菌株對常用抗菌藥物的敏感性[16]。
結合藥敏結果,根據CLSI推薦的雙紙片協同擴散試驗,分析分離菌株產超廣譜β-內酰胺酶和金屬β-內酰胺酶的情況。將分離菌株的純培養物調整為0.5麥氏濁度,均勻涂布于固體培養基表面,分別將含有頭孢他啶(30 μg)、頭孢他啶/克拉維酸(30 μg/10 μg)、頭孢噻肟(30 μg)及頭孢噻肟/克拉維酸(30 μg/10 μg)藥敏紙片貼于平板,37 ℃恒溫培養18~24 h,觀察并記錄各抗生素的IZD。若頭孢他啶、頭孢噻肟分別與克拉維酸聯用時的抑菌圈比單獨使用時直徑增加5 mm以上,則表明分離菌株為ESBLs陽性。
使用EDTA-紙片協同擴散試驗,檢測分離菌株產MBLs情況。取分離菌株0.5麥氏濁度的菌液均勻涂布于固體培養基上,將兩片含亞胺培南(10 μg·片-1)的藥敏片貼于平板表面,兩紙片相距15 mm,向其中一片亞胺培南藥敏片上滴加10 μL 0.5 mol·L-1的EDTA,置于37℃持續培養24 h,觀察并記錄藥敏片的抑菌情況。若兩個藥敏片抑菌圈的直徑差異超過5 mm,則判定檢測菌株產MBL[17-19]。
1.9.4" 中藥體外抑制試驗
連翹、黃連、五倍子、肉桂、款冬花、金銀花、炒僵蠶、金蝎、桑皮、薄荷、蘇子、桑白皮、黃芩、茯苓、烏梅、秦皮、荊芥、五味子、蒲公英、枳殼、板藍根、羌活、炙白附子、魚腥草、貝川母、胖大海、首烏、羅漢果共28味中藥購自南京同仁堂大藥房,每種中藥各取50 g,加入100 mL蒸餾水浸泡、煮2 h后過濾藥液;再加入蒸餾水重復熬制,合并兩次藥液,小火濃縮后制成濃度為1 g·mL-1的中藥藥液[20]。空白藥敏紙片浸泡在1 g·mL-1中藥提取液中,待浸透后風干備用。取分離菌株純培養物均勻涂布在含 5%血清的TSA固體培養基表面,待菌液稍吸收后取含中藥藥液的藥敏片貼于平板上,37 ℃靜置培養24 h,測量IZD;設置3個平行,并計算平均值和標準差。
1.10" 致病性試驗
1.10.1" 毒力基因檢測
合成Pm 23種毒力基因的引物,以分離菌株的基因組為模板,PCR法篩查分離菌株攜帶毒力基因情況[21]。
1.10.2" 動物回歸試驗
取分離菌株的純培養物接種于含5%血清的TSB液體培養基,37 ℃、180 r·min-1培養12 h,無菌PBS梯度稀釋并經平板計數,制備2×106、2×104、2×102和2×101 CFU·mL-1的菌懸液。小鼠于實驗室內暫養一周,分成5組,每組10只,每組腹腔注射不同濃度的菌液,每只小鼠腹腔注射0.2 mL,對照組小鼠注射同等體積無菌PBS;攻毒后觀察記錄小鼠的精神狀態和死亡情況,剖檢死亡小鼠,記錄臟器病變情況,采集病變組織進行病原菌的分離與測序鑒定。
2" 結" 果
2.1" 致病菌的分離和鑒定
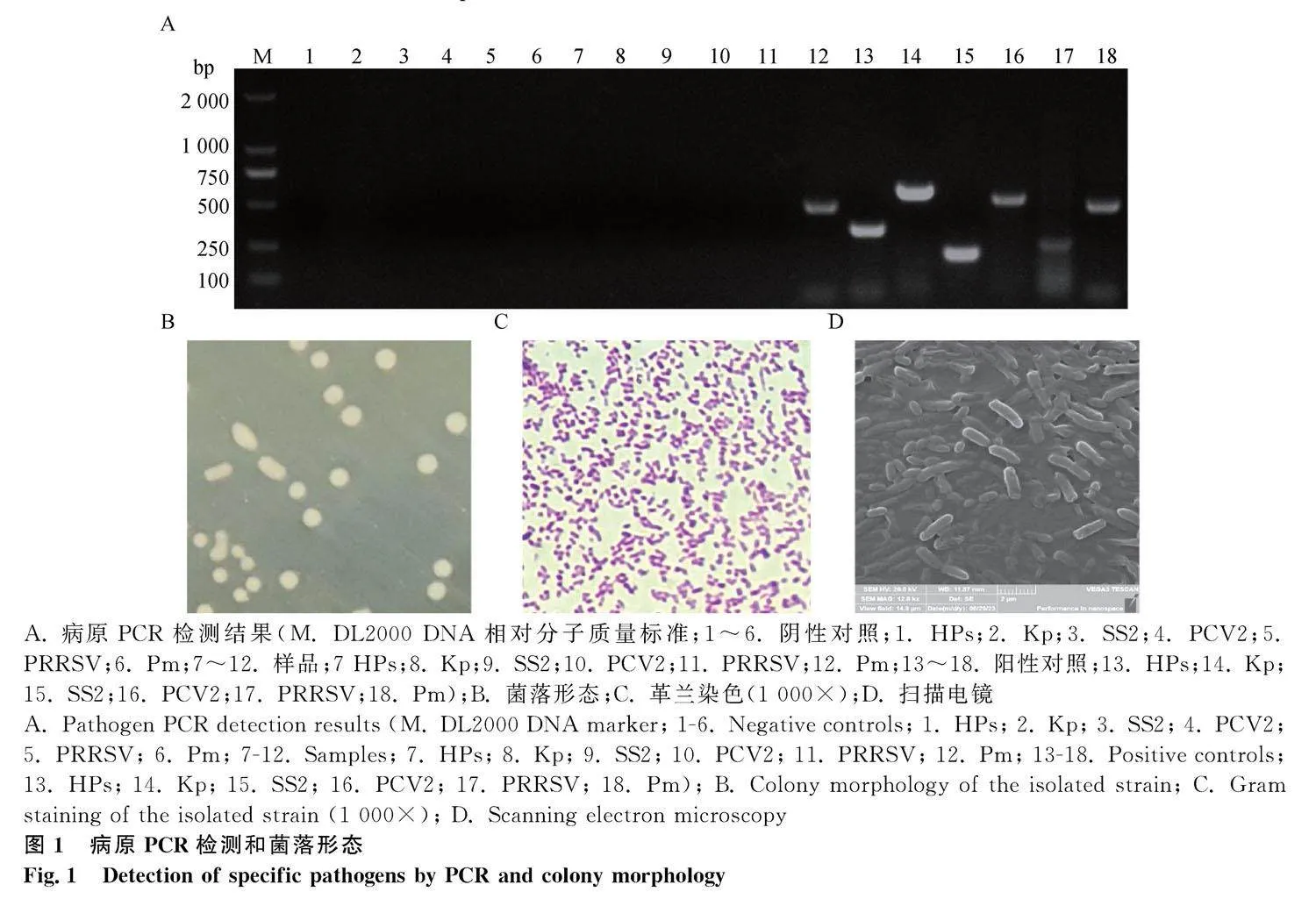
患呼吸道疾病病豬剖檢時可見肺臟組織明顯病變,肺組織壞死、外周有纖維組織包裹。提取病豬病變組織的基因組進行病原PCR篩查,結果如圖1A所示,多殺性巴氏桿菌的引物擴增出460 bp大小的條帶,與多殺性巴氏桿菌參考菌株擴增的條帶大小一致;PCV2、PRRSV、SS2、HPS和Kp的檢測均為陰性,推測該豬場病豬的呼吸道疾病可能與多殺性巴氏桿菌感染相關。無菌條件下取病變肺組織劃線培養,分離菌株在TSA固體培養基上生長出圓形露珠樣、有光澤的菌落(圖1B);革蘭染色后,鏡檢可見紅色革蘭陰性短桿菌,兩極濃染,多呈單個存在(圖1C);掃描電鏡下可見單在或成雙排列、兩端鈍圓的短桿狀菌(圖1D)。生化鑒定結果顯示,分離菌株能夠發酵葡萄糖、木糖、甘露醇、山梨醇和果糖,不能發酵衛茅醇、阿拉伯糖、鼠李糖、麥芽糖、乳糖,肌醇、硫化氫、馬尿酸鈉、甲基紅和V-P試驗為陰性,其生化特性與《伯杰手冊》中巴氏桿菌的生化特性一致。
2.2" 16S rDNA擴增及進化樹構建
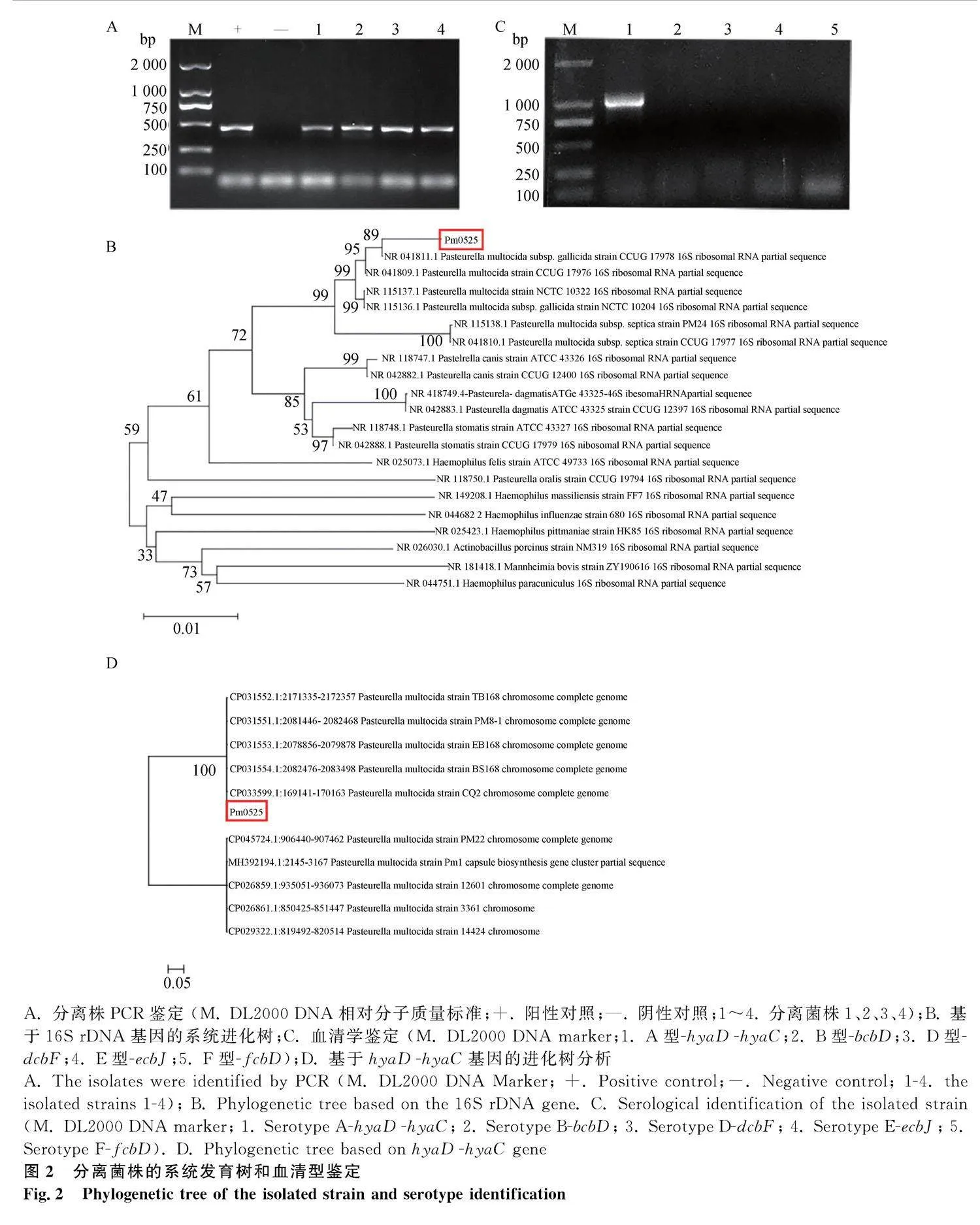
用提取的分離菌株DNA為模板,以Pm的特異性引物進行PCR擴增,結果如圖2A所示,從病變組織中分離出的四株菌均擴增出與Pm陽性對照大小一致的特異性條帶。通用引物擴增分離菌株的16S rDNA并進行同源序列分析,結果顯示,分離菌株與多殺性巴氏桿菌亞種CCUG 17978(NR.041811.1)基因序列的相似性最高,在系統發育樹中分離菌株與多殺性巴氏桿菌聚為一支(圖2B),確定分離菌株為多殺性巴氏桿菌,命名為Pm0525。
2.3" 血清型鑒定
多殺性巴氏桿菌莢膜分型引物的PCR結果顯示,分離菌株Pm0525擴增出1 044 bp左右的條帶,與多殺性巴氏桿菌A型的預期條帶大小一致(圖2C);其它的血清型引物均無特異性條帶擴增。分離菌株hyaD-hyaC基因的PCR產物測序分析發現,分離菌株Pm0525的hyaD-hyaC基因與GenBank中其他A型多殺性巴氏桿菌hyaD-hyaC基因的序列相似性達99.7%;構建進化發育樹結果如圖2D所示,分離菌株與A型多殺性巴氏桿菌PM8-1等匯成一支,確定分離菌株Pm0525為莢膜血清型A型。
2.4" MLST分型
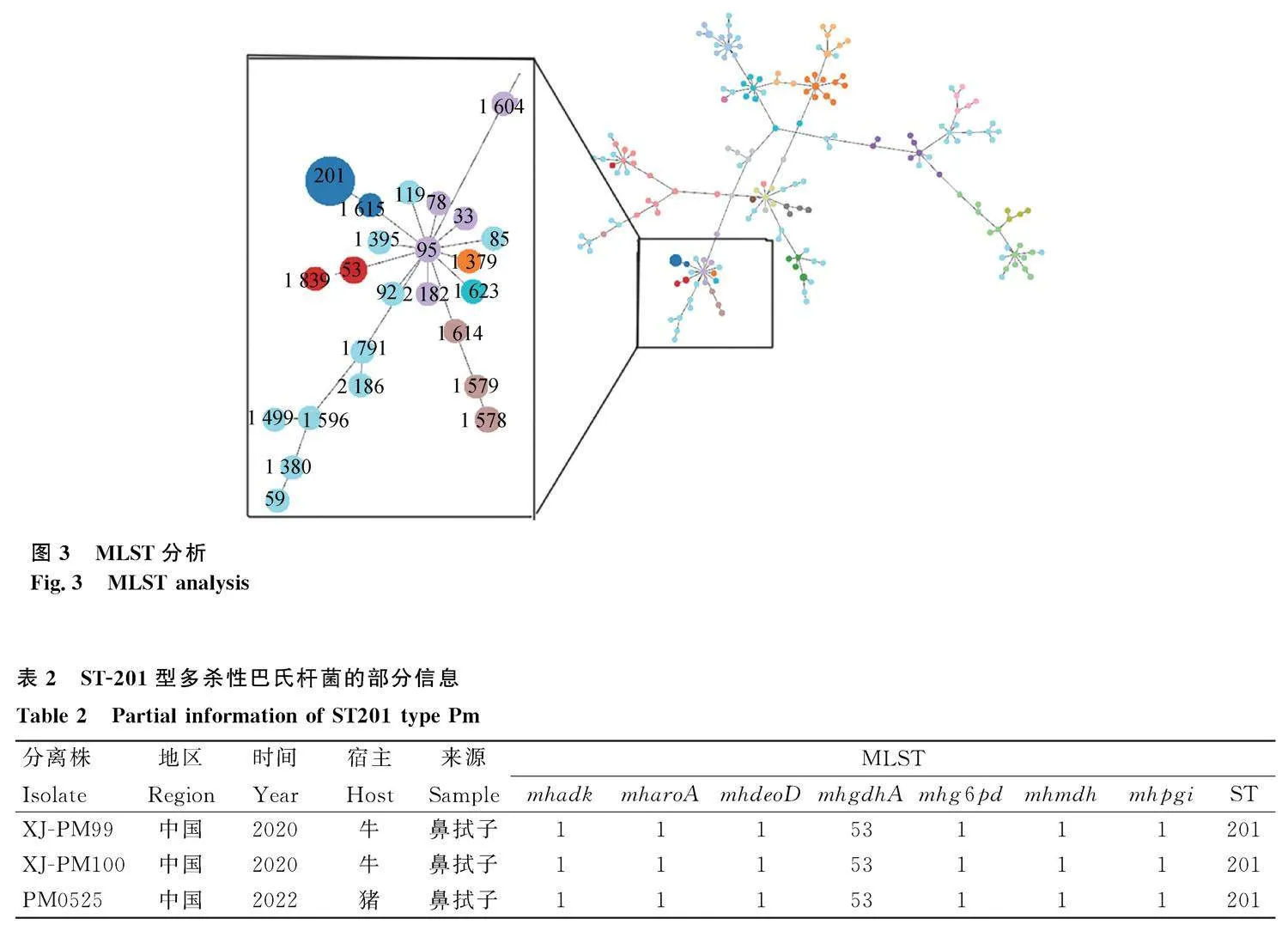
對菌株Pm0525的管家基因進行PCR和測序,序列比對結果顯示:Pm0525的7個管家基因(adk、aroA、deoD、gdhA、g6pd、hmdh 和pgi)的編號分別為1、1、1、53、1、1和1,經比對確定分離菌株Pm0525為ST201型。數據收集發現,已有的Pm ST201型病例均來自國內,主要來源于牛的鼻拭子(表2)。選擇Pm不同的ST型進行分組聚類分析,結果如圖3所示,豬源ST201型多殺性巴氏桿菌與ST1615型聚為一支,兩者親緣關系最近。
2.5" 藥敏試驗
2.5.1" 耐藥基因檢測
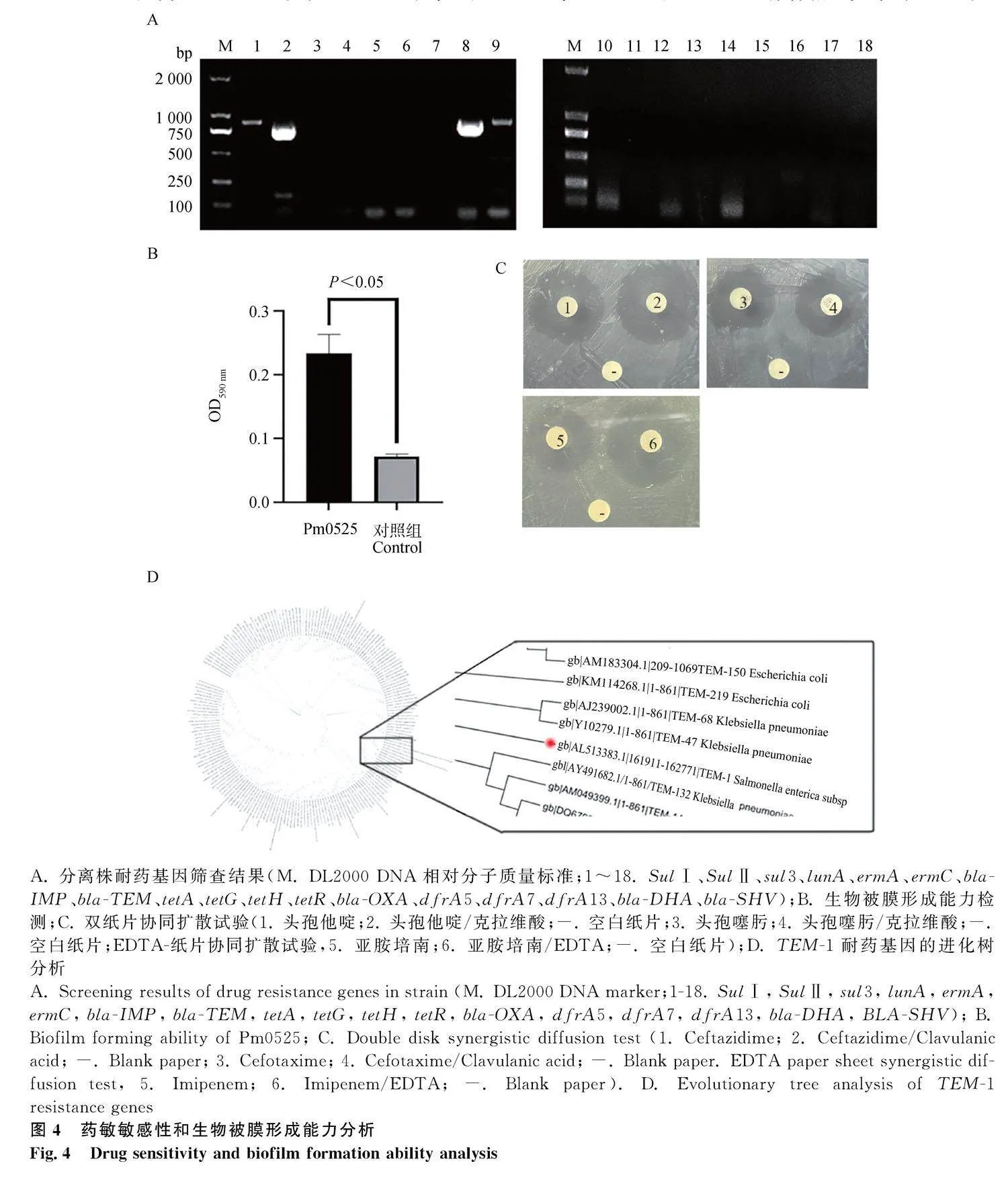
耐藥基因檢測結果如圖4A所示,在篩查的18中耐藥基因中,Pm0525攜帶SulⅠ、SulⅡ、tetA 和bla-TEM四種耐藥基因,表明分離菌株Pm0525具有一定的耐藥性,為多重耐藥菌株。
2.5.2" 生物被膜形成能力測定
通過結晶紫染色法測得分離菌株Pm0525的OD590 nm為0.2650,空白對照組的OD590 nm值為0.071(圖4B),分離菌株的OD590 nm值介于2ODc到4ODc之間,表明Pm0525具有中等生物被膜形成能力。
2.5.3" 抗生素敏感性測定
抗生素的敏感性試驗結果如表3所示,Pm0525對慶大霉素、卡那霉素、麥迪霉素、新霉素、哌拉西林、頭孢氨芐、頭孢咪唑、頭孢拉定、頭孢哌酮、環丙沙星、諾氟沙星、丁胺卡那、氧氟沙星13種抗生素敏感,對紅霉素、呋喃唑酮、羧芐西林、氨芐西林4種抗生素中度敏感,對氯霉素、克林霉素、頭孢呋辛、頭孢他啶和四環素5種抗生素呈現耐藥。
雙紙片協同擴散試驗結果顯示,分離菌株對頭孢他啶及頭孢他啶/克拉維酸的IZD分別為24和25 mm,對頭孢噻肟及頭孢噻肟/克拉維酸的IZD分別為20和21 mm,頭孢他啶和頭孢噻肟分別與克拉維酸組合均未使IZD顯著增加,表明Pm0525為不產ESBLs細菌;EDTA-紙片協同擴散試驗結果顯示,分離菌株對美羅培南及美羅培南/EDTA的IZD分別為13和23 mm(圖4C),進一步判定Pm0525為MBL陽性菌株。雙紙片協同擴散試驗證實,分離菌株為一株產MBL的耐藥菌株。對分離菌株Pm0525攜帶的β-內酰胺類抗生素耐藥基因bla-TEM進行測序分析,并將測序結果進行BLAST在線工具進行比對。結果顯示,分離菌株的bla-TEM基因與TEM-1(GenBank登錄號:JX268625.1)的基因序列一致;可確定分離菌株的bla-TEM基因為TEM-1型。從CARD耐藥基因數據庫下載所有bla-TEM基因分型序列,構建進化發育樹,分析耐藥分型之間的關系,發現其TEM-1與TEM-47、TEM-132有著相近關系(圖4D)。
2.5.4" 中藥抑制試驗結果
中藥藥敏試驗結果如表4所示,連翹、黃連和五倍子等28味中藥中,以黃連對Pm0525的抑菌效果最好,其IZD為19 mm,為高度敏感;五倍子以及荊芥的IZD分別為11和10 mm,為中度敏感;烏梅、連翹、肉桂、秦皮的IZD低于10 mm,均為低敏;其他中藥對分離菌株Pm0525無顯著抑制作用。
2.6" 致病性試驗
2.6.1" 毒力基因鑒定
毒力基因的篩查結果如圖5A、B所示,在23種毒力基因中,Pm0525攜帶18種:sodA、sodC、tadD、pfhA、Hsf-2、fimA、ptfA、pmHAS、plpB、omp87、ompH、ompA、Fur、tonB、exbD、exbB、hgbA和nanH,不攜帶毒力基因tbpA、toxA、hsf-1、hgbB和nanB。
2.6.2" 動物回歸試驗
Pm0525經腹腔注射試驗組小鼠后,最高濃度組小鼠在攻毒后4 h開始出現反應遲鈍、呼吸急促、被毛凌亂,抱團取暖等臨床癥狀,感染后14 h死亡8只,死亡率為80%;2.0×104 CFU·mL-1組小鼠在攻毒后3 d死亡3只,死亡率為30%;對照組小鼠未見異常。根據攻毒后小鼠死亡情況,計算出分離菌株的半數致死量(LD50)為2.5×105 CFU。死亡小鼠剖檢發現,死亡小鼠的肺臟出血,以高劑量組的病變最為明顯;肝臟腫大、出血;脾臟和腎臟均有不同程度的出血和壞死(圖5C);死亡小鼠的肝、肺、腦、脾、心和腎組織中的載菌量分別為3.08×109、5.23×108、1.29×108、3.64×108、5.68×108和1.1×109 CFU·g-1,其中肝臟中細菌載量最高(圖5D),表明分離菌株具有一定的組織嗜性。病理組織學觀察發現,與健康對照組相比,感染組小鼠的肺泡隔毛細血管充血,肺泡內可見大量纖維素和中性粒細胞滲出;脾臟的白髓有大量中性粒細胞浸潤;肝中央靜脈中有中性粒細胞滲出(圖5E),表明分離株Pm0525具有較強的致病性,可造成小鼠嚴重的臟器損傷。
3" 討" 論
多殺性巴氏桿菌病作為一種人獸共患病,多殺性巴氏桿菌是人和動物呼吸道疾病重要的條件致病性病原菌,具有極強的致病性,造成嚴重的經濟損失和危害公共安全[22-24]。近年來,在豬群中不斷分離到不同血清型的Pm,臨床上常與多種病毒、細菌共同感染或繼發感染,引起高度重視[25-26]。本研究從患呼吸道疾病豬肺組織中分離到一株優勢菌,經電鏡觀察、生理生化和16S rDNA 基因測序分析,確定分離菌株為豬源多殺性巴氏桿菌。該養殖企業采集病豬肺組織進行宏基因組測序,測序結果顯示病豬肺組織檢測到了一定豐度的多殺性巴氏桿菌核酸序列。本研究病原分離結果與該養殖企業宏基因組測序和病原分析結果相一致。血清型鑒定分離菌株為莢膜血清型A型,這與國內豬源多殺性巴氏桿菌流行的優勢莢膜型為A型的流調結果相符[8,27]。MLST鑒定分離菌株的分子分型為ST201型菌株,該型主要分布在國內,已報道的病例均來源于牛,未見該型在國外的報道。流行病學調查發現,我國豬群中流行的基因型主要為ST11型、ST10型和ST3型[8];本研究系首次分離到豬源多殺性巴氏桿菌ST201型菌株(莢膜血清型A型),提示Pm作為一種人獸共患病病原,對畜禽養殖業和公共衛生安全具有潛在威脅。
毒力基因檢測發現,分離菌株攜帶18種毒力基因,毒力型為sodA+/sodC+/tadD+/pfhA+/Hsf-2+/fimA+/ptfA+/pmHAS+/plpB+/omp87+/ompH+/ompA+/Fur+/tonB+/exbD+/exbB+/hgbA+/nanH+其中毒力基因fimA、hsf-2、tadD、pfhA與Pm的菌毛和黏附素相關,sodA和sodC與Pm的超氧化物歧化酶相關,nanH與Pm的唾液酸酶相關,plpB、omp87、ompH、ompA與外膜蛋白相關,Fur、tonB、exbD和exbB與鐵攝取相關;pmHAS與Pm的透明質酸酶相關[8];研究證實,pmHAS基因與莢膜血清A型顯著相關,pfhA 和 pmHAS 基因在莢膜血清A型的Pm菌株中的檢出率較高[28]。豬源多殺性巴氏桿菌ST201型菌株(莢膜血清型A型)攜帶毒力基因pfhA 和pmHAS,與前人的研究相一致[29]。動物致病性試驗結果顯示,攻毒劑量為2×106 CFU·mL-1時能夠導致小鼠在攻毒后14 h 80%的死亡,并出現肺臟、脾臟和肝臟等臟器嚴重的病變,具有較強的致病性。Pm0525的耐藥基因檢測結果顯示,其攜帶SulⅠ、SulⅡ、tetA、bla-TEM四種耐藥基因,包括含有針對β-內酰胺類抗生素的耐藥基因bla-TEM;雙藥敏片協同試驗證實,Pm0525是通過產MBL的方式對β-內酰胺類抗生素耐藥,其具體的耐藥機制有待進一步深入研究[30-32];對氯霉素、克林霉素、頭孢呋辛、頭孢他啶、四環素五種藥物耐藥,屬于多重耐藥菌株;其對四環素耐藥可能與耐藥基因tetA相關,頭孢呋辛、頭孢他啶屬于β-內酰胺類抗生素,對其產生的耐藥性可能與耐藥基因bla-TEM有關。在生物被膜形成能力方面,分離株Pm0525具有中等的生物被膜形成能力,這與分離菌株在藥敏試驗中表現的多重耐藥性相符。結合分離株KP0123生物被膜形成能力、耐藥基因以及藥敏篩查結果,在一定程度解釋了該養殖場豬群Pm感染病情反復和抗生素治療不理想的原因。提示臨床在給予畜群使用抗菌藥物之前,應監測耐藥菌譜,減少耐藥性的產生和避免濫用抗生素[33]。
28味中藥的體外抑菌試驗發現,黃連對分離菌株Pm0525的抑菌效果最好;五倍子、荊芥、烏梅、連翹、肉桂、秦皮均為低敏,與李達怡等單味中藥對巴氏桿菌的抑菌效果一致[34]。研究表明,包括黃連堿、小檗堿、巴馬汀和表小檗堿組成的生物堿是黃連的主要活性成分,具有廣譜抗細菌、抗真菌、抗病毒的作用[35-36]。在黃連堿抑制禽多殺性巴氏桿菌的研究中發現,黃連堿可通過影響多殺性巴氏桿菌基因組、蛋白質合成、呼吸代謝、細菌結構的相關基因表達的方式發揮抑菌作用[37]。黃連中的黃連素可通過抑制肺炎克雷伯菌生物膜的形成與下調肺炎克雷伯菌的Ⅰ型和Ⅲ型菌毛結構基因表達的方式發揮抗菌活性[38]。同時,黃連素與抗生素的聯合使用可顯著增強抑菌效果[39],這可為臨床上開展中藥黃連與抗生素聯合用藥治療豬多殺性巴氏桿菌感染提供借鑒,做到合理用藥和精準施治。
豬多殺性巴氏桿菌常定殖在消化道或者呼吸道,在應激或者長途運輸、氣候劇變、抵抗力降低時誘發Pm的感染,是重要的人畜共患條件致病菌[1,9]。對于該菌感染的防治首要是改善養殖環境,強化飼養管理,提高動物機體免疫力,減少感染概率。在治療時,兼顧Pm的流行特點、致病性、耐藥性以及敏感抗生素和中藥的聯合用藥,嚴格控制病原的流行和傳播。因此,針對上述問題,應進一步加強Pm的流行病學調查和傳播機制研究,探究其多宿主感染和致病機制,為系統性、源頭防控該菌引發的感染奠定基礎。
4" 結" 論
本試驗證實從患病豬肺組織中分離出一株巴氏桿菌(血清A型),ST型為ST201,攜帶sodA、sodC、tadD、pfhA、Hsf-2、fimA、ptfA、pmHAS、plpB、omp87、ompH、ompA、Fur、tonB、exbD、exbB、hgbA、nanH 18種毒力基因,具有一定的致病性,且對公共衛生安全具有潛在威脅;分離菌株具有多重耐藥特性,攜帶SulⅠ、SulⅡ、tetA、bla-TEM四種耐藥基因,可通過產金屬 β-內酰胺酶(MBL)方式發揮抗β-內酰胺類抗生素的作用;對慶大霉素、卡那霉素、麥迪霉素等13種抗生素敏感,對氯霉素、克林霉素、頭孢呋辛、頭孢他啶和四環素耐藥;中藥黃連在體外試驗中對分離菌株Pm0525有顯著抑菌作用。相關結果旨在為ST201型巴氏桿菌(血清型A型)的研究提供科學參考。
參考文獻(References):
[1]" WILKIE I W, HARPER M, BOYCE J D, et al. Pasteurella multocida: diseases and pathogenesis[M]//AKTORIES K, ORTH J, ADLER B. Pasteurella multocida: Molecular Biology, Toxins and Infection. Berlin: Springer, 2012:1-22.
[2]" 李貴琴. 牛巴氏桿菌的分離鑒定及黃芩苷對巴氏桿菌肺炎的治療作用[D]. 楊凌:西北農林科技大學, 2022.
LI G Q. Isolation and identification of Pasteurella bovis and the therapeutic effect of baicalin on Pasteurella pneumonia[D]. Yangling:Northwest Aamp;F University, 2022. (in Chinese)
[3]" SARANGI L N, THOMAS P, GUPTA S K, et al. Virulence gene profiling and antibiotic resistance pattern of Indian isolates of Pasteurella multocida of small ruminant origin[J]. Comparat Immunol Microbiol Infect Dis, 2015, 38:33-39.
[4]" CARTER G R. The type specific capsular antigen of Pasteurella multocida[J]. Canad J Med Sci, 1952, 30(1):48-53.
[5]" TOWNSEND K M, BOYCE J D, CHUNG J Y, et al. Genetic organization of Pasteurella multocida cap Loci and development of a multiplex capsular PCR typing system[J]. J Clin Microbiol, 2001, 39(3):924-929.
[6]" HARPER M, JOHN M, TURNI C, et al. Development of a rapid multiplex PCR assay to genotype Pasteurella multocida strains by use of the Lipopolysaccharide outer core biosynthesis Locus[J]. J Clin Microbiol, 2015, 53(2):477-485.
[7]" BROGDEN K A, RHOADES K R, HEDDLESTON K L. A new serotype of Pasteurella multocida associated with fowl cholera[J]. Avian Dis, 1978, 229(1):185-190.
[8]" 彭" 忠, 梁" 婉, 艾偉誠, 等. 我國豬群中多殺性巴氏桿菌的基因型分析[J]. 畜牧獸醫學報, 2019, 50(5):1064-1072.
PENG Z, LIANG W, AI W C, et al. Genotypical characteristics of swine Pasteurella multocida in China[J]. Acta Veterinaria et Zootechnica Sinica, 2019, 50(5):1064-1072. (in Chinese)
[9]" 王豪男. 我國部分省區豬源多殺性巴氏桿菌的分子流行病學調查[D]. 武漢:華中農業大學, 2017.
WANG H N. Molecular epidemiology of Pasteurella multocida from swine in some provinces of China[D]. Wuhan:Huazhong Agricultural University, 2017. (in Chinese)
[10]" 許文博, 吳麗梅, 劉" 鑫, 等. 1株多殺性巴氏桿菌的全基因組序列及致病相關基因分析[J]. 畜牧獸醫學報, 2022, 53(6):1858-1869.
XU W B, WU L M, LIU X, et al. Analysis on complete genome sequence and pathogenic genes of a Pasteurella multocida strain[J]. Acta Veterinaria et Zootechnica Sinica, 2022, 53(6):1858-1869. (in Chinese)
[11]" WANG Y H, ZHU J, LU C P, et al. Evidence of circulation of an epidemic strain of Pasteurella multocida in Jiangsu, China by multi-locus sequence typing (MLST)[J]. Infect Genet Evolut, 2013, 20:34-38.
[12]" 張亞楠, 李亞菲, 陳汝佳, 等. A:L1 ST128型鴨源多殺性巴氏桿菌的耐藥性及毒力分析[J]. 畜牧獸醫學報, 2021, 52(10):2852-2863.
ZHANG Y N, LI Y F, CHEN R J, et al. Resistance and virulence analysis of type A:L1 ST128 Pasteurella multocida from Ducks[J]. Acta Veterinaria et Zootechnica Sinica, 2021, 52(10):2852-2863. (in Chinese)
[13]" MAY B J, ZHANG Q, LI L L, et al. Complete genomic sequence of Pasteurella multocida, Pm70[J]. Proc Natl Acad Sci USA, 2001, 98(6):3460-3465.
[14]" CHRISTENSEN H, SAJID S M, BISGAARD M, et al. Prediction of Pasteurella multocida serotypes based on whole genomic sequences[J]. Vet Microbiol, 2022, 271:109492.
[15]" 鄔" 琴, 張星星, 顧曉曉, 等. 牛源多殺性巴氏桿菌血清分型及毒力相關基因的檢測研究[J]. 中國畜牧獸醫, 2020, 47(6):1910-1920.
WU Q, ZHANG X X, GU X X, et al. Study on serotyping and detection of virulence-associated genes of Pasteurella multocida isolated from bovine[J]. China Animal Husbandry amp; Veterinary Medicine, 2020, 47(6):1910-1920. (in Chinese)
[16]" 王" 斐, 楊" 潔, 呂慶杰, 等. 致腦膜炎多殺性巴氏桿菌的分離鑒定及全基因組重測序[J]. 畜牧獸醫學報, 2022, 53(12):4346-4355.
WANG F, YANG J, L Q J, et al. Isolation and genomic characterization of a meningitis causing Pasteurella multocida[J]. Acta Veterinaria et Zootechnica Sinica, 2022, 53(12):4346-4355. (in Chinese)
[17]" 阮紫涵, 黃安雄, 王秀娟, 等. CLSI、EUCAST和中國耐藥判定標準概述[J]. 生物技術通報, 2022, 38(9):47-58.
RUAN Z H, HUANG A X, WANG X J, et al. Overview of CLSI, EUCAST, and susceptibility breakpoints in China[J]. Biotechnology Bulletin, 2022, 38(9):47-58. (in Chinese)
[18]" 林祥宏. 金屬β-內酰胺酶檢測方法學研究進展[J]. 國際檢驗醫學雜志, 2007, 28(3):255-258.
LIN X H. Research progress in the detection methodology of Metallo-β-lactamases[J]. International Journal of Laboratory Medicine, 2007, 28(3):255-258. (in Chinese)
[19]" YONG D, LEE K, YUM J H, et al. Imipenem-EDTA disk method for differentiation of metallo-β-lactamase-producing clinical isolates of Pseudomonas spp." and Acinetobacter spp. [J]. J Clin Microbiol, 2002, 40(10):3798-3801.
[20]" ARUHOMUKAMA D. Review of phenotypic assays for detection of extended-spectrum β-lactamases and carbapenemases:a microbiology laboratory bench guide[J]. Afr Health Sci, 2020, 20(3):1090-1108.
[21]" 徐" 州, 周德端, 段國勛, 等. 中藥對幽門螺桿菌抑殺作用的實驗研究[J]. 中國醫藥學報, 1993, 8(5):25-26.
XU Z, ZHOU D D, DUAN G X, et al. Experimental study on the inhibitory and killing effects of traditional Chinese medicine on Helicobacter pylori[J]. Chinese Medical Journal, 1993, 8(5):25-26. (in Chinese)
[22]" WILSON B A, HO M. Pasteurella multocida:from zoonosis to cellular microbiology[J]. Clin Microbiol Rev, 2013, 26(3):631-655.
[23]" 王立程, 朱" 雄, 陳" 海, 等. 四株臨床分離多殺性巴氏桿菌分子特征分析[J]. 疾病監測, 2023, 38(8):983-988.
WANG L C, ZHU X, CHEN H, et al. Molecular characterization of four strains of Pasteurella multocida isolated from clinical sources[J]. Disease Surveilliance, 2023, 38(8):983-988. (in Chinese)
[24]" 張哲瑋, 曹維維, 代小童, 等. 豬多殺性巴氏桿菌的分離鑒定、血清分型及耐藥性分析[J]. 中國獸醫學報, 2023, 43(5):930-936.
ZHANG Z W, CAO W W, DAI X T, et al. Isolation, identification, serotyping and drug resistance analysis of Pasteurella multocida suis[J]. Chinese Journal of Veterinary Science, 2023, 43(5):930-936. (in Chinese)
[25]" 王" 羽, 董文龍, 王" 巍, 等. 豬源莢膜血清F型多殺性巴氏桿菌分離鑒定[J]. 中國獸醫雜志, 2017, 53(6):31-33, 36.
WANG Y, DONG W L, WANG W, et al. Isolation and identification of a Pasteurella multocida strain belonged to capsular type F from swine[J]. Chinese Journal of Veterinary Medicine, 2017, 53(6):31-33, 36. (in Chinese)
[26]" 林星宇, 胡" 凌, 王" 印, 等. 豬源多殺性巴氏桿菌的分離鑒定及莢膜血清分型[J]. 浙江農業學報, 2016, 28(4):558-562.
LIN X Y, HU L, WANG Y, et al. Isolation, identification and capsule serotyping of Pasteurella multocida originated from swine[J]. Acta Agriculturae Zhejiangensis, 2016, 28(4):558-562. (in Chinese)
[27]" PENG Z, WANG H N, LIANG W, et al. A capsule/lipopolysaccharide/MLST genotype D/L6/ST11 of Pasteurella multocida is likely to be strongly associated with swine respiratory disease in China[J]. Arch Microbiol, 2018, 200(1):107-118.
[28]" 胡" 璇, 蔡" 平, 李紅婷, 等. 一株豬源A型多殺性巴氏桿菌的分離鑒定[J]. 中國獸醫科學, 2021, 51(12):1568-1578.
HU X, CAI P, LI H T, et al. Isolation and identification of a strain of Pasteurella multocida type A from swine[J]. Chinese Veterinary Science, 2021, 51(12):1568-1578. (in Chinese)
[29]" TANG X B, ZHAO Z Q, HU J Y, et al. Isolation, antimicrobial resistance, and virulence genes of Pasteurella multocida strains from swine in China[J]. J Clin Microbiol, 2009, 47(4):951-958.
[30]" 張曉兵, 府偉靈, 廖" 揚, 等. 臨床產ESBLs細菌的耐藥特征和基因分型的研究[J]. 中華醫院感染學雜志, 2005, 15(4):386-389.
ZHANG X B, FU W L, LIAO Y, et al. Identification, antibacterial susceptibility and gene typing of clinical ESBLs-producing Bacteria[J]. Chinese Journal of Nosocomiology, 2005, 15(4):386-389. (in Chinese)
[31]" 司紅彬, 宋劍武, 王鵬霞, 等. 救必應中藥血清與抗菌藥聯合對產ESBLs細菌抑菌效果的研究[J]. 中國畜牧獸醫, 2015, 42(4):1009-1014.
SI H B, SONG J W, WANG P X, et al. Study on the antibacterial effect of Chinese material medica serum of holly bark combined with antimicrobials on ESBLs-producing bacteria[J]. China Animal Husbandry amp; Veterinary Medicine, 2015, 42(4):1009-1014. (in Chinese)
[32]" JU L C, CHENG Z S, FAST W, et al. The continuing challenge of metallo-β-lactamase inhibition:mechanism matters[J]. Trends Pharmacol Sci, 2018, 39(7):635-647.
[33]" 李" 娟, 鄧秋連, 龍" 燕, 等. 腸桿菌科產ESBLs細菌的臨床耐藥研究[J]. 中國當代醫藥, 2017, 24(15):126-128.
LI J, DENG Q L, LONG Y, et al. Clinical drug resistance of Enterobacteriaceae producing ESBLs bacteria[J]. China Modern Medicine, 2017, 24(15):126-128. (in Chinese)
[34]" 李達怡, 佘琳曼, 黃妙容, 等. 中藥聯合對巴氏桿菌的體外抑菌作用研究[J]. 中獸醫醫藥雜志, 2020, 39(3):82-85.
LI D Y, SHE L M, HUANG M R, et al. Bacteriostasis of traditional Chinese medicine on Pasteurella in vitro[J]. Journal of Traditional Chinese Veterinary Medicine, 2020, 39(3):82-85. (in Chinese)
[35]" 姚姍姍, 劉靜茹, 陳" 靜, 等. 基于網絡藥理學探究中藥黃連抗菌作用機制[J]. 中國畜牧獸醫, 2020, 47(5):1593-1601.
YAO S S, LIU J R, CHEN J, et al. Study on anti-bacterial mechanism of Coptidis rhizoma based on network pharmacology[J]. China Animal Husbandry amp; Veterinary Medicine, 2020, 47(5):1593-1601. (in Chinese)
[36]" MENG F C, WU Z F, YIN Z Q, et al. Coptidis rhizoma and its main bioactive components:recent advances in chemical investigation, quality evaluation and pharmacological activity[J]. Chin Med, 2018, 13:13.
[37]" 張" 睿. 黃連堿含量的測定及其對禽多殺性巴氏桿菌抑菌機制的研究[D]. 荊州:長江大學, 2023.
ZHANG R. Determination of the content of coptisine and its inhibition mechanism against Pasteurella multocida[D]. Jingzhou:Yangtze University, 2023. (in Chinese)
[38]" 劉冬梅, 郭夢雨, 費" 冰, 等. 黃連素抑制肺炎克雷伯菌生物膜形成作用機制研究[J]. 中國現代醫藥雜志, 2024, 26(1):6-10.
LIU D M, GUO M Y, FEI B, et al. Research on the mechanism of Berberine inhibiting the formation of Klebsiella pneumoniae biofilm[J]. Modern Medicine Journal of China, 2024, 26(1):6-10. (in Chinese)
[39]" 干鐵兒, 林少華, 李文杰, 等. 黃連素聯合亞胺培南西司他丁對耐碳青霉烯類肺炎克雷伯菌的體外抑菌作用[J]. 中華醫院感染學雜志, 2021, 31(21):3211-3215.
GAN T E, LIN S H, LI W J, et al. In vitro antibacterial effect of berberine combined with imipenem-cilastatin on carbapenem-resistant Klebsiella pneumoniae[J]. Chinese Journal of Nosocomiology, 2021, 31(21):3211-3215. (in Chinese)
(編輯" 白永平)
