
荔枝LcWRKY47基因的克隆、亞細胞定位、表達及延緩果實褐變的功能初探
2024-12-31 00:00:00程南譜王鑫MuhammadMuzammalAslam邵遠志李雯
熱帶作物學報 2024年10期

關鍵詞:荔枝;WRKY 轉錄因子;基因克隆;亞細胞定位;表達分析
中圖分類號:S667.1 文獻標志碼:A
荔枝(Litchi chinensis Sonn.)是無患子科荔枝屬的亞熱帶常綠果樹,是非呼吸躍變型水果,原產于中國南方[1],主要產區為廣東、廣西、福建、海南等地區,果肉多汁,口感極佳,營養豐富,深受大眾喜愛,經濟價值高,有“嶺南佳果”的美稱[2],是我國亞熱帶地區重要的果樹品種之一,也是我國特色經濟果樹[3]。但由于荔枝在夏季高溫高濕的特殊時期成熟,果實含水量高,新陳代謝旺盛,采收后營養物質會被迅速消耗[4],生理變化劇烈,同時果皮上的病原菌迅速生長,容易引起荔枝褐變和變質,導致荔枝商品經濟價值下降[5],荔枝自身的衰老褐變嚴重影響到荔枝的采后貯藏品質。草酸(oxalic acid, OA)是一種普遍存在的有機酸,在各種代謝過程中起著重要作用[6]。據報道,OA 可以通過提高果實抗病能力和抗氧化活性,延緩呼吸速率和果實成熟衰老,有效地保持園藝產品的品質[7]。
WRKY 轉錄因子家族是植物中最大的轉錄因子家族之一[8]。第1 個WRKY 基因SPE1 于1994年從甘薯中分離得到[9],隨著其他植物的基因組測序,越來越多的WRKY 基因在不同植物物種中得到鑒定和克隆,已在擬南芥、番茄、小麥、大豆、水稻和玉米等許多物種中得到鑒定[10]。轉錄因子通常包含4 個功能區,即轉錄調控域(激活區和抑制區)、DNA 結合域、寡聚化位點和核定位信號,使轉錄因子在不同情況發揮不同功能[11]。之所以將其命名為WRKY 轉錄因子,是因為WRKY蛋白質序列包含60 個氨基酸殘基組成的DNA 結合域,并且該結構域的N 端具有WRKYGOK 序列組成的WRKY 高度保守結構域[12]。然而,在少數WRKY 蛋白中,WRKYGQK 的氨基酸序列存在不同的突變類型,有WRKYGEK、WRKYGMK、WRKYGKK、WSKYEQK 或WIKYGEN 等。一般而言N 端至少存在1 個WRKY 保守結構域,C 端則包含1 個鋅指結構基序C2H2(C-X4-5-C-X22-23-H-X-H) 或C2HC(C-X7-C-X23-H-X-C)[13] 。根據WRKY 結構域的數量和鋅指基序的類型,WRKY轉錄因子可分為3個主要亞家族,Ⅰ類亞家族有2個含有C2H2 型鋅指基序的WRKY 結構域,Ⅱ類亞家族有1 個含有C2H2 型鋅指基序的WRKY結構域,Ⅲ類亞家族具有1 個含有C2H2 型鋅指基序的WRKY 結構域[14]。
大量研究證明,WRKY 轉錄因子在植物的生長發育中起著重要作用,參與調節植物多種生理過程,擬南芥AtWRKY23 調控根毛發育[15],小麥TaWRKY71 調控種子萌發等過程[16] , 玉米ZmWRKY11 調控種皮發育[17],棉花GhWRKY15調控莖生長[18]、參與碳水化合物合成、次生代謝產物合成,擬南芥AtWRKY75 通過影響水楊酸(salicylic acid, SA)積累和H2O2 清除參與調控葉片衰老過程[19],蕓苔型油菜BnWRKY47 能夠促進氮從老葉到幼葉或種子的再動員,使幼葉優先生長,抑制整株植株的衰老死亡[20]。
迄今為止,越來越多WRKY 轉錄因子在植物生長發育以及生物與非生物脅迫中的功能得到研究。但目前關于荔枝的WRKY 轉錄因子的研究較少,對荔枝LcWRKY47 基因的研究未見報道。因此,本研究以采后妃子笑荔枝為試驗材料,克隆LcWRKY47 基因,進行生物信息學分析和亞細胞定位分析, 同時通過RT-qPCR (quantitativereal-time, PCR)分析LcWRKY47 基因在果皮和果肉中的表達水平,進一步利用過表達體系分析LcWRKY47 轉錄因子調控采后荔枝成熟衰老的功能,為深入分析WRKY 轉錄因子在采后荔枝果實中的生物學功能提供理論依據。
1 材料與方法
1.1 材料
選用妃子笑荔枝鮮果為試驗材料,挑選無病蟲害、無機械損傷、形狀、大小、外觀顏色一致的荔枝果實。以2 mmol/L OA 溶液浸泡10 min 的荔枝果實為處理組,以清水浸泡10 min 為對照組,處理后隨機選取30 個荔枝為1 組,每組3 個重復,分別裝入保鮮袋中,貯藏在8 ℃、相對濕度為85%~90%的培養箱中,每2 d 取1 次樣,用液氮將所取樣品速凍后保存在?80 ℃超低溫冰箱中,用于后續試驗。
1.2 方法
1.2.1 RNA 的提取與LcWRKY47 基因全長克隆以荔枝果皮為材料,使用改良的CTAB 法從果皮、果肉中提取RNA[21],隨后分別逆轉錄成cDNA。使用NOVA 逆轉錄試劑盒(All-in-One First-StrandSynthesis MasterMix with dsDNase)進行cDNA 的合成。利用Primer 6 軟件設計LcWRKY47 的全長擴增特異性引物,序列分別為LcWRKY47-F:5’-ATGTCAAGTTCTAAAATGTC-3’,LcWRKY47-R:5’-CTAGTTGGTAGAGAAAGTGG-3’。用2×Phanta?Max Master Mix (Dye plus)高保真酶進行PCR 擴增,PCR 反應程序如下:95 ℃預變性30 s;95 ℃變性15 s,55 ℃退火15 s,72 ℃延伸30 s,35 個循環;最后72 ℃反應5 min,得到LcWRKY47 的開放閱讀框(open reading frame, ORF)序列,PCR 產物經過瓊脂糖凝膠電泳檢測,隨后用Omega 膠回收試劑盒(E.Z.N.A. Gel Extraction Kit)回收目的基因條帶。純化產物連接pCE2 TA/Blunt-Zero 載體(Vazyme TA/Blunt-Zero Cloning" Kit),然后將連接產物轉化至大腸桿菌感受態(維地生物,E. coliDH5α),挑取陽性菌落于LB 培養基中培養,送生工生物工程(上海)股份有限公司測序。
1.2.2 LcWRKY47 蛋白的生物信息學分析 利用ExPASy-ProtParam ( https://web.expasy. org/protparam)分析蛋白的分子量、等電點、不穩定系數等理化性質,利用ExPASy-ProtScale(https://web.expasy.org/protscale)分析蛋白的親疏水性,利用TMHMM( https://services.healthtech.dtu.dk/services/TMHMM-2.0)和SignalP(https://services.healthtech.dtu.dk/services/SignalP-6.0)分析蛋白質的跨膜結構及信號肽預測。利用SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa%20_sopma.html)分析LcWRKY47 蛋白的二級結構,并使用SWISS-MODEL(https://swissmodel.expasy.org)分析預測LcWRKY47 蛋白的三維結構;利用在線網站Clustal Omega(https://www.ebi.ac.uk/jdispatcher/msa/clustalo)進行多序列比對;利用在線網站MEME 5.5.0(https://memesuite.org/meme)分析motif 結構,將motif 數設置為8;使用MEGA 5 軟件構建系統進化樹,并使用鄰接法構建。
1.2.3 亞細胞定位 利用Novopro Bioscience(https://www.novoprolabs.com/tools/nls-signal-prediction)預測蛋白的核定位序列(nuclear localizationsequence, NLS)。使用洋蔥內表皮作為侵染材料,通過洋蔥內表皮轉化法進行亞細胞定位分析[22]。設計雙酶切正向引物F:TCTAGAATGTCAAGTTCTAAAATGTCAGCCC,反向引物R:GGTACCGTTGGTAGAGAAAGTGGTGCAAGA。通過雙酶切法將LcWRKY47 基因構建到pCAMBIA1300-GFP 載體上,生成35S::LcWRKY47-GFP 融合表達載體,將重組質粒轉化至GV3101 農桿菌,制備OD600 為0.8~1.0 的懸浮液, 再將洋蔥內表皮(1.5 cm×1.5 cm)浸泡在懸浮液中減壓10 min,用濾紙吸去洋蔥內表皮上多余的菌液,再將其轉移到鋪有濾紙的MS 固體培養基上,放置在25 ℃培養箱中避光培養2 d 后,制成玻片,在熒光生物攝像顯微鏡下觀察熒光信號。
1.2.4 LcWRKY47 基因的表達量分析 利用Primer5 軟件設計LcWRKY47 基因定量引物,分別為正向引物F:5-CACGGCAATGGCTAACACAACC-3‘,反向引物R:5‘-AATGGTGCAGAAGCGGAGAGG-3‘; 內參基因引物Actin-F : 5‘-ACCGTATGAGCAAGGAAATCACTG-3‘,Actin-R: 5‘-TCGTCGTACTCACCCTTTGAAATC-3‘ 。使用試劑盒(TOLOBIO 2×Q3 SYBR qPCR Master Mix)進行RT-qPCR 擴增,采用2?ΔΔCt 法計算基因的相對表達量,每次測定均有3個重復。
1.2.5 瞬時過表達體系驗證LcWRKY47 的功能設計雙酶切正向引物F : 5’-GGATCCATGTCAAGTTCTAAAATGTC-3‘, 反向引物R: 5‘-GAATTCCTAGTTGGTAGAGAAAGTGG-3‘"。酶切位點為:上游BamH Ⅰ(GGATCC),下游EcoRⅠ(GAATTC)。通過雙酶切法將擴增的LcWRKY47編碼序列構建到pGreenII 62-SK 載體上,并將SKLcWRKY47和空SK 載體轉化至農桿菌GV3101,其中空載體組作為對照。參考GONG 等[23]的方法,并進行一些修改,在果實莖部附近注射1 mL 侵染液,然后將注射過的荔枝果實在侵染液中減壓浸泡20 min,對荔枝果實進行侵染,隨后將侵染過的荔枝果實置于(25±1)℃、相對濕度為85%~90%條件下進行培養。在注射后的第1 天和第3天進行取樣,通過RT-qPCR 測定對應基因的表達量,每次測定均有3 個重復。
1.3 數據處理
使用Office 2021軟件進行數據統計與計算,使用SPSS 22.0 軟件分析數據差異顯著性(IBMCorp,美國),采用Duncan 法分析比較差異數據顯著性(Plt;0.05),使用GraphPad Prism 8 軟件對LcWRKY47基因相對表達量進行作圖。
2 結果與分析
2.1 LcWRKY47 基因的克隆及其蛋白的理化性質分析
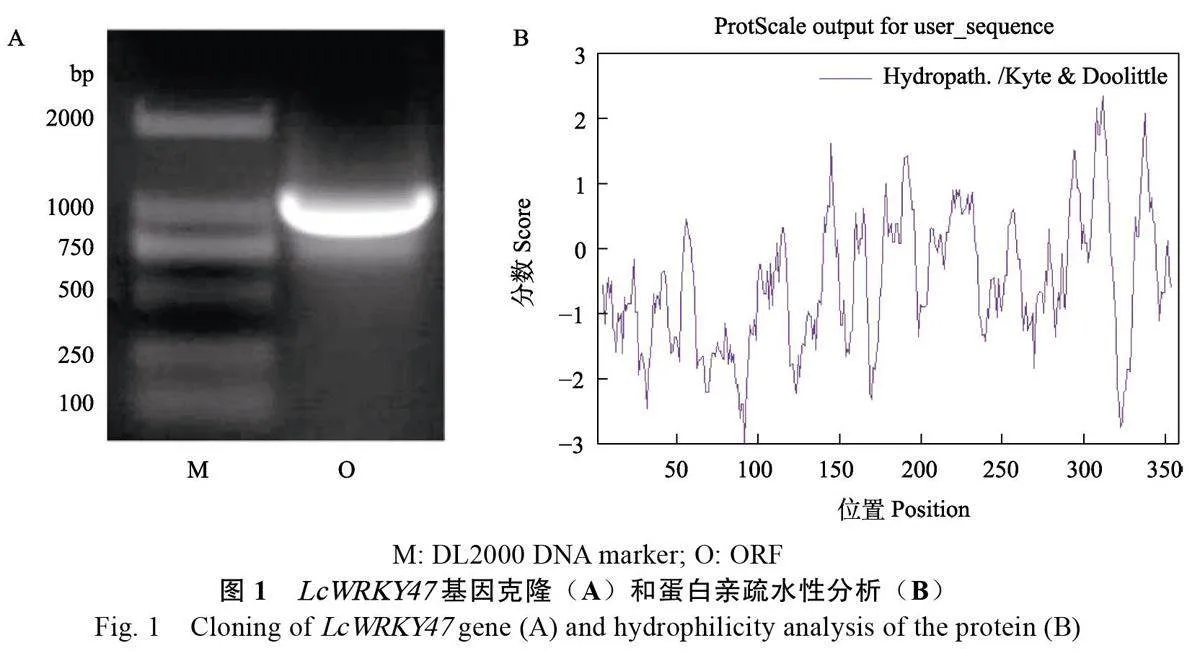
使用LcWRKY47 的全長引物克隆出ORF 長度為1077 bp的擴增片段(圖1A)。LcWRKY47基因編碼358 個氨基酸,對LcWRKY47 蛋白理化性質進行預測,結果顯示LcWRKY47 蛋白分子式為C1651H2622N490O525S21, 相對分子質量為38 409.21 kDa,總原子數為5309,理論等電點為8.84;氨基酸組成中帶負電殘基數量(Asp+Glu)為26 個,帶正電殘基數量(Arg+Lys)為31 個,蛋白質中脂肪族側鏈氨基酸指數為58.94,總平均親水性值是?0.535,表現為親水性(圖1B),不穩定性指數是54.03,為不穩定蛋白。綜上,該蛋白為堿性不穩定親水蛋白。
2.2 LcWRKY47蛋白結構分析
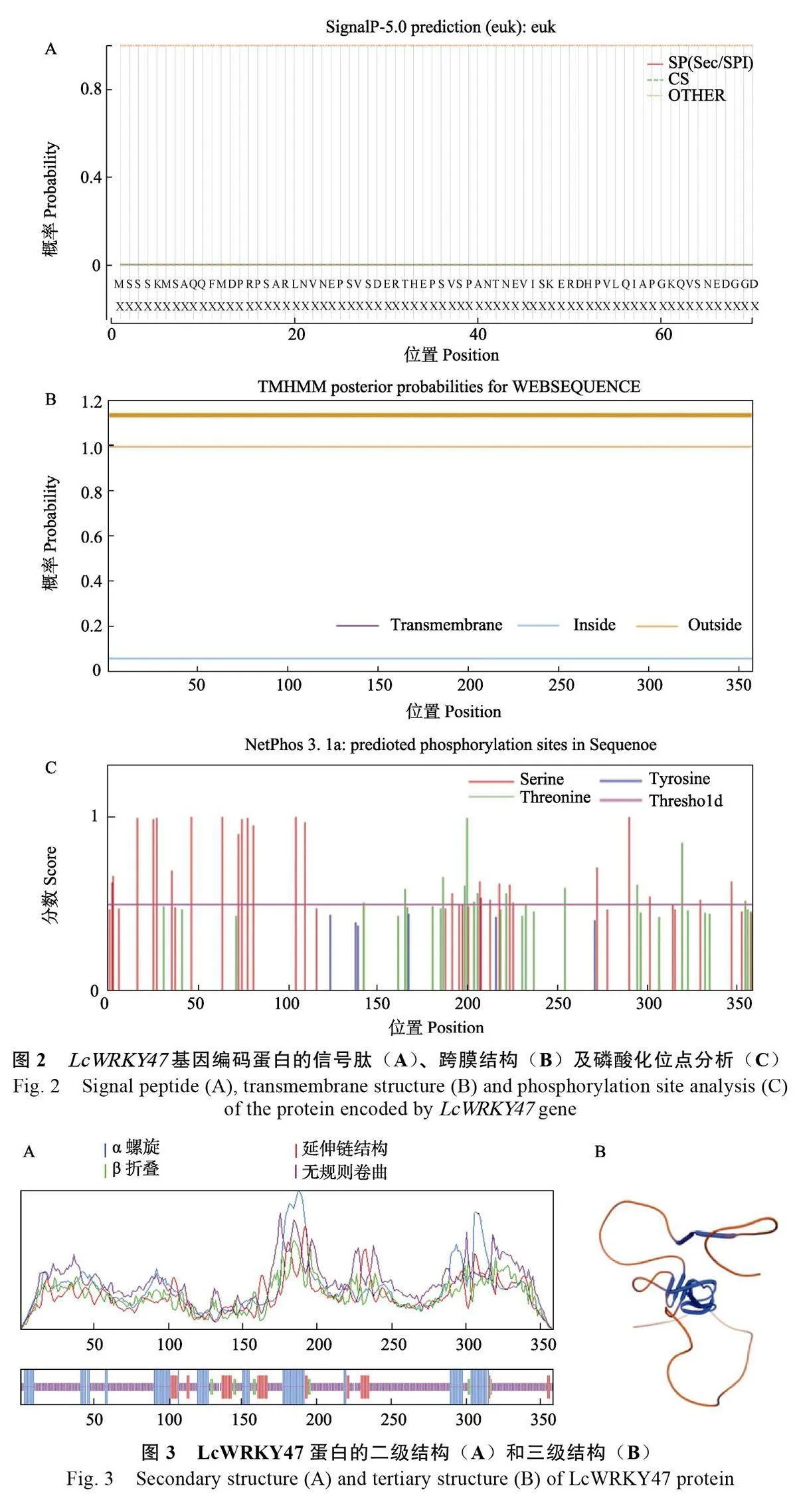
預測結果顯示LcWRKY47 蛋白中不存在信號肽(圖2A),也并未發現跨膜區域(圖2B),不是跨膜蛋白。LcWRKY47 蛋白的磷酸化可能發生在絲氨酸(Ser)和蘇氨酸(Thr)位點上,主要為絲氨酸位點,說明此蛋白發揮調控功能可能是以絲氨酸位點的磷酸化修飾為主(圖2C)。LcWRKY47 蛋白的二級結構以無規則卷曲為主,占比為65.64%,其余為α-螺旋,占21.79%、延伸鏈占9.50%、β-折疊占3.07%(圖3A)。運用SWISS-MODEL 在線程序對荔枝LcWRKY47 蛋白的三維結構進行建模分析,發現LcWRKY47蛋白質的主要結構為無規則卷曲和α-螺旋(圖3B),其結果與二級結構的預測結果相一致。
2.3 LcWRKY47 蛋白motif 分析及同源序列比對
選取9 個與LcWRKY47 同源性較高的WRKY 轉錄因子進行motif 分析,結果顯示motif結構相似,在所有蛋白中均含有相同的6種motif;與荔枝LcWRKY47 所含motif 數目和種類一致的有2 個物種,分別為甜橙和陸地棉(圖4A);氨基酸同源序列比對結果顯示其氨基酸序列均包含1 個WRKY 結構域和1 個C2H2 型鋅指基序。上述結果表明LcWRKY47 蛋白屬于第Ⅱ類WRKY轉錄因子(圖4B)。
2.4 LcWRKY47 蛋白的系統進化分析
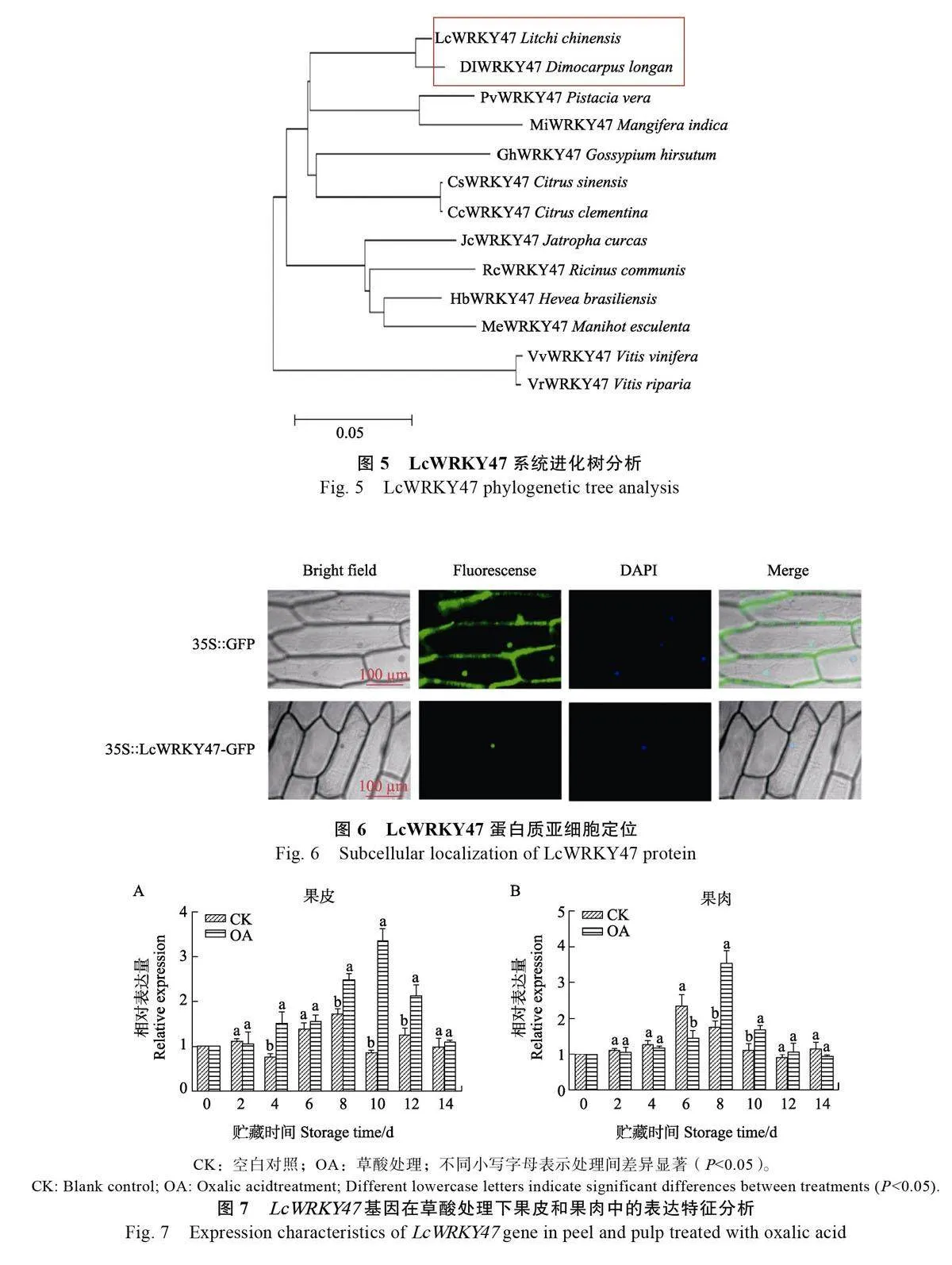
選取無患子科植物、漆樹科植物、蕓香科植物以及大戟科植物等不同物種WRKY 基因構建進化樹,結果顯示,荔枝LcWRKY47 與同為無患子科的龍眼DlWRKY47 聚在一起,親緣關系最近,其次是杧果、阿月渾子和甜橙,而橡膠樹、木薯、葡萄以及河岸葡萄則與荔枝LcWRKY47 親緣關系較遠(圖5)。
2.5 亞細胞定位結果分析
Novopro Bioscience 工具預測分析LcWRKY47蛋白的核定位序列為PFRKARVSVRA,位于蛋白序列中98~108 位置。在熒光生物攝像顯微鏡下觀察到,35S::GFP 對照洋蔥內表皮細胞中的細胞核和細胞壁均檢測到綠色熒光信號, 而35S::LcWRKY47-GFP 融合蛋白的綠色熒光信號僅出現在細胞核中,這表明荔枝LcWRKY47 蛋白在洋蔥內表皮細胞中定位于細胞核,符合轉錄因子所具有的核定位特征(圖6)。
2.6 LcWRKY47基因的表達量分析
外源OA 處理后,LcWRKY47 基因在荔枝果皮和果肉中的表達水平如圖7 所示。在荔枝果皮中LcWRKY47 基因的表達量呈現先上調后下調的趨勢(圖7A),CK 在8 d 達到最大值,然后逐漸下降,而OA 處理組在10 d 達到最大值,之后呈現下降的趨勢,且OA 處理組只有在2 d 時低于CK,8 d 后顯著高于CK;在荔枝果肉中LcWRKY47 基因的表達量呈現先上調后下調的趨勢(圖7B),CK 在6 d 達到最大值,OA 處理組在8 d 達到最大值,之后均呈現下降的趨勢,相較于CK,8 d 后處理組LcWRKY47 基因的表達水平整體高于CK。LcWRKY47 在荔枝果皮與果肉中的表達模式相同,在貯藏后期表達量均上調,且顯著高于CK,表明LcWRKY47 轉錄因子可能在OA 調控果實成熟衰老過程中起正調控作用。
2.7 瞬時過表達驗證LcWRKY47的基因表達量
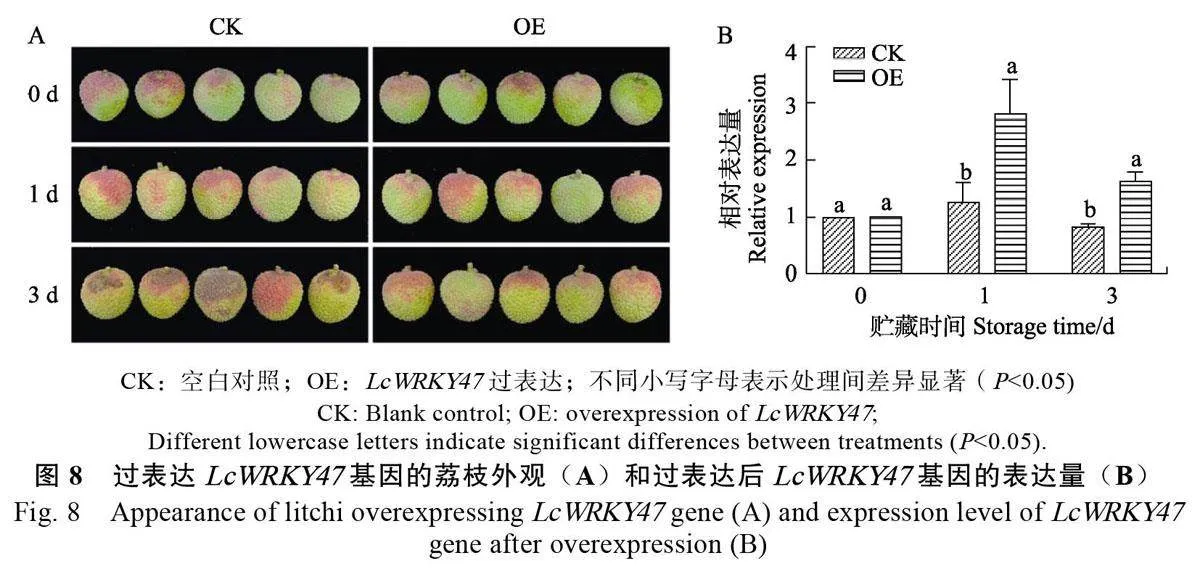
為了驗證LcWRKY47 能否調控荔枝衰老褐變,將LcWRKY47 基因轉入完整的荔枝果實。從圖8A 中可以看出,在3 d 時,CK 的整果外觀失色與褐變情況比轉基因荔枝嚴重,而此時轉基因荔枝并未出現褐變等明顯衰老現象。從圖8B 中可以看出,在轉基因果實中LcWRKY47 的表達量高于CK。綜上,LcWRKY47 轉錄因子可能通過延緩荔枝的衰老與褐變來維持果實采后品質。
3討論
在植物生長發育的過程中,WRKY 轉錄因子起著不可忽視的作用[24]。WRKY 蛋白能夠作為激活因子或阻遏因子參與細胞質和細胞核過程,包括從細胞器或細胞質到細胞核的信號傳遞事件,調控植物對生物和非生物刺激的反應過程,參與協調發育過程中的內部信號。WRKY 可與靶基因啟動子中的W-box[TGACC(A/T)]結合,激活或抑制下游基因的表達,調控其應激反應[25]。最新的研究發現WRKY 在蔬菜葉片成熟、衰老等代謝過程中的重要作用,FAN 等[26]從中國白菜葉片中克隆出BrWRKY65,發現BrWRKY65 的表達隨著采后葉片葉綠素降解和變黃而上調,并且可以調控與葉綠素降解相關的BrNYC1 、BrSGR1 和BrDIN1 等3 個衰老相關基因,從而影響采后葉片的衰老。有研究報道WRKY 在果實成熟、衰老等多種生理過程中的重要作用,JI 等[27]發現桃果實中PpSDH 和PpCOX15 等能量代謝基因能與PpWRKY46 互作并被PpWRKY46 上調表達,調節桃子的能量代謝,維持桃果實的品質與抗病性,延緩桃果實的成熟衰老。綜上,推測WRKY 在荔枝果實中可能存在類似的生物學功能,但目前在荔枝中的WRKY 轉錄因子研究卻鮮有報道。本研究中的荔枝LcWRKY47 基因屬于WRKYⅡ類亞家族,蛋白具有親水性,蛋白不穩定,不包含信號肽,無跨膜結構;二級結構和三級結構預測中包括α-螺旋、β-折疊、延伸鏈和無規則卷曲,以無規則卷曲為主,同時具有WRKY 保守結構域和1 個C2H2 型鋅指基序,系統進化樹分析表明LcWRKY47 與同為無患子科的DlWRKY47 親緣關系最近。
妃子笑荔枝主要產于海南、廣東,皮薄肉厚核小,香味濃郁,多汁爽口,是中早熟優質品種之一,具有很高的經濟價值和研究價值[28]。然而荔枝采后難貯存、易變質、果皮易褐變等情況一直是荔枝保鮮目前存在的現實問題[29]。OA 與植物的抗微生物、生物和非生物抗性有關[30],是極好的抗氧化劑,可以增強超氧陰離子的清除能力,阻止活性氧的形成[31],活性氧的產物之一是丙二醛,是植物細胞膜脂質過氧化的產物,造成細胞衰老和死亡[32]。OA 處理可通過延緩呼吸速率和成熟,降低丙二醛濃度,延遲衰老,增強抗病能力,有效保持園藝產品的新鮮度和品質[33]。近年來,OA 處理采后果實的保鮮效果受到廣泛關注,已有研究報道OA 能夠延遲香蕉的成熟,抑制杧果的腐爛,并防止荔枝果實產生褐變[34–35]。OA 作為一種果實保鮮劑,可能通過影響植物轉錄因子的表達來延緩果實的衰老與褐變,從而起到保鮮的效果。因此本研究將采后妃子笑荔枝作為試驗材料,使用OA 進行處理,研究LcWRKY47基因的表達模式。本研究鑒定的LcWRKY47 基因定量結果表明,LcWRKY47 在荔枝果皮和果肉中的表達趨勢一致,在儲藏后期LcWRKY47 基因表達量與CK 相比差異顯著,均顯著高于CK,處于較高表達水平。將LcWRKY47 連接到過表達SK 載體,對荔枝果實進行侵染實驗,使LcWRKY47 基因得到過表達,結果表明從外觀上看,過表達后的荔枝果實比CK 衰老及褐變程度更低,同時LcWRKY47基因的表達量顯著高于CK。綜合以上結果推測,LcWRKY47 轉錄因子可能在荔枝衰老與褐變過程中起著一定的調節作用,延緩了果實的衰老與褐變, 保持采后妃子笑荔枝的貯藏品質。關于LcWRKY47 基因后續的功能驗證還有待更深入的研究。
