木薯MeMinD的互作蛋白篩選
2024-12-31 00:00:00潘沐陸小花姚遠陳銀華郭建春王亞杰
熱帶作物學報 2024年10期

關鍵詞:木薯;MeMinD;MeNAPRT2;酵母雙雜交;質體
中圖分類號:S533 文獻標志碼:A
植物質體(葉綠體、淀粉體等)的分裂由一系列蛋白協同完成[1]。MinD 參與質體分裂的精細調控,在質體形態維持中起著關鍵作用。MinD有2 個重要的結構域:MinC 和MinE 結合結構域。MinC 基因所編碼的MinC 蛋白是細胞分裂抑制因子,在MinD 的ATPase 活性催化下,與其相互作用并結合形成復合物MinCD,之前,充當“鐘擺”功能的MinE 在兩極之間來回移動,驅動MinCD復合物來回振蕩,使Z 環只能在中部進行組裝,促進質體可以均等分裂成2 個[2]。當MinD 過量表達,會讓復合物MinCD 遍布質體所有位點,抑制Z環形成,從而抑制質體分裂;而MinD 的減少或缺失,使Z 環的形成不局限在中部,形成偏離中心或多個FtsZ 環,質體分裂發生在不適當的位置[3-4]。
目前關于質體的研究多以葉綠體為對象,過量表達擬南芥AtMinD 會導致葉綠體增大[5],而減少AtMinD 表達量或缺失該基因的突變體,引起葉綠體不對稱分裂[4]。將AtMinD 在煙草過表達,轉基因植株的葉綠體大,且數目減少[6]。這些結果都進一步證實了MinD 在質體分裂中的關鍵作用。淀粉體是木薯塊根淀粉積累的場所,葉綠體是葉片臨時性淀粉合成和降解的細胞器。前期,本實驗室克隆了木薯的MeMinD 基因,異源表達于大腸桿菌可導致菌體的分裂位點錯位和絲狀形成,亞細胞定位分析表明MeMinD 可定位于葉綠體[7]。通過基因編輯敲除MeMinD 基因,突變體木薯的葉綠體和淀粉體顯著增大(待發表)。以上結果表明MeMinD 蛋白參與調控木薯質體的分裂,然而與MeMinD 蛋白協同調控木薯質體分裂的相關蛋白尚未明確。本研究將MeMinD 構建至pGBKT7 載體,進行酵母雙雜交文庫篩選實驗,挖掘與MeMinD 互作的質體分裂相關蛋白,為解析木薯質體分裂調控機制提供新的信息。
1 材料與方法
1.1 材料
華南8 號木薯(SC8, Manihot esculenta Crantz)、木薯cDNA 文庫、酵母雙雜交質粒pGBKT7、pGADT7、pGADT7-T、pGBKT7-lam、pGBKT7-p53由本實驗室提供;大腸桿菌DH5α 感受態、酵母菌AH109 感受態購自上海唯地生物技術有限公司;缺陷培養基SD/-Trp(SD/T)、SD/-Trp-Leu(SD/TL)、SD/-Trp-Leu-His-Ade(SD/TLHA)、X-α-Gal 購自北京酷來搏科技有限公司。
1.2 方法
1.2.1 pGBKT7-MeMinD的構建與毒性檢測 將酶切純化的誘餌載體pGBKT7(BD)載體與純化MeMinD 基因片段進行同源重組,轉化大腸桿菌DH5α 感受態細胞,進行菌落PCR,篩選陽性單克隆,構建酵母雙雜交載體pGBKT7-MeMinD。
提取pGBKT7-MeMinD 重組質粒和pGBKT7(BD)空載質粒并分別轉化酵母菌AH109,將陽性克隆用SD/T 液體培養基擴大培養,將OD600 控制在0.8~1.0,再將菌液按1、10–1、10–2、10–3、10–4濃度梯度進行稀釋,分別點在SD/T 平板上,30 ℃培養2~3 d,觀察菌落生長情況。
1.2.2 自激活檢驗將 pGBKT7-MeMinD 質粒與pGADT7(BD)空載質粒共轉入AH109 酵母菌株,將陽性酵母液體培養基擴大培養,分別點在SD/TL、SD/TLHA、SD/TLHA+X-α-Gal 三種平板上培養,觀察實驗組(pGADT7+pGBKT7-MeMinD)和對照組(陽性對照:pGBKT7-p53+pGADT7-T,陰性對照:pGBKT7-lam+pGADT7-T)的酵母菌在3 種平板上的生長情況,以此確定MeMinD 蛋白是否存在自激活現象。
1.2.3 MeMinD互作蛋白篩選 參照仇婷婷[8]的方法進行酵母雙雜交文庫篩選MeMinD 的候選互作蛋白。將變藍的酵母菌斑進行PCR 檢測,PCR 產物送至生工生物工程(上海)股份有限公司進行測序,再利用NCBI Blast 軟件進行序列比對,篩選出木薯MeMinD 可能互作的蛋白并進行功能注釋。
1.2.4 酵母雙雜交點對點驗證 設計候選互作蛋白的基因擴增引物(表1),克隆其編碼區并連接到pGADT7 載體。PCR 篩選陽性克隆并測序驗證。將候選互作蛋白基因的pGADT7 重組質粒與pGBKT7-MeMinD 質粒共轉AH109 酵母菌株,進行酵母雙雜交點對點驗證,篩選MeMinD 的互作蛋白。
2 結果與分析
2.1 誘餌載體pGBKT7-MeMinD的構建
以SC8 木薯cDNA為模板,PCR 擴增MeMinD的編碼區序列。電泳檢測結果顯示擴增片段與預期大小(987 bp)一致(圖1)。將MeMinD 基因片段與線性化的pGBKT7 載體重組連接,轉化大腸桿菌感受態細胞。挑取5 個單菌落進行菌落PCR 鑒定,獲得4 個陽性克隆并進行測序驗證(圖2)。提取測序正確的pGBKT7-MeMinD 誘餌載體,用于后續酵母雙雜交文庫篩選。
2.2 毒性檢測和自激活檢測
將pGBKT7-MeMinD 和pGBKT7空載分別轉化酵母感受態AH109,將菌液稀釋不同濃度(1、10–1、10–2、10–3、10–4)后在SD/TL 培養基上培養。結果顯示轉pGBKT7-MeMinD 和pGBKT7空載的酵母菌生長狀態無差異(圖3),說明MeMinD對AH109 酵母菌無毒性,可進行后續實驗。
將pGBKT7-MeMinD 和pGADT7 空載共轉酵母菌株進行自激活檢測。在SD/TL 培養基上,共轉pGBKT7-MeMinD 和pGADT7 空載的酵母菌、陽性對照(pGBKT7-p53+pGADT7-T)和陰性對照(pGBKT7-lam+pGADT7-T)的酵母菌在SD/TL培養基上均能正常生長,但在SD/TLHA 和SD/TLHA+X-α-Gal 培養基上只有陽性對照的酵母菌生長,且陽性對照在SD/TLHA+X-α-Gal 培養基上的酵母菌落變藍。此結果表明MeMinD 蛋白不具有自激活活性,可用于酵母雙雜交篩選木薯cDNA 文庫(圖4)。
2.3 MeMinD互作蛋白篩選
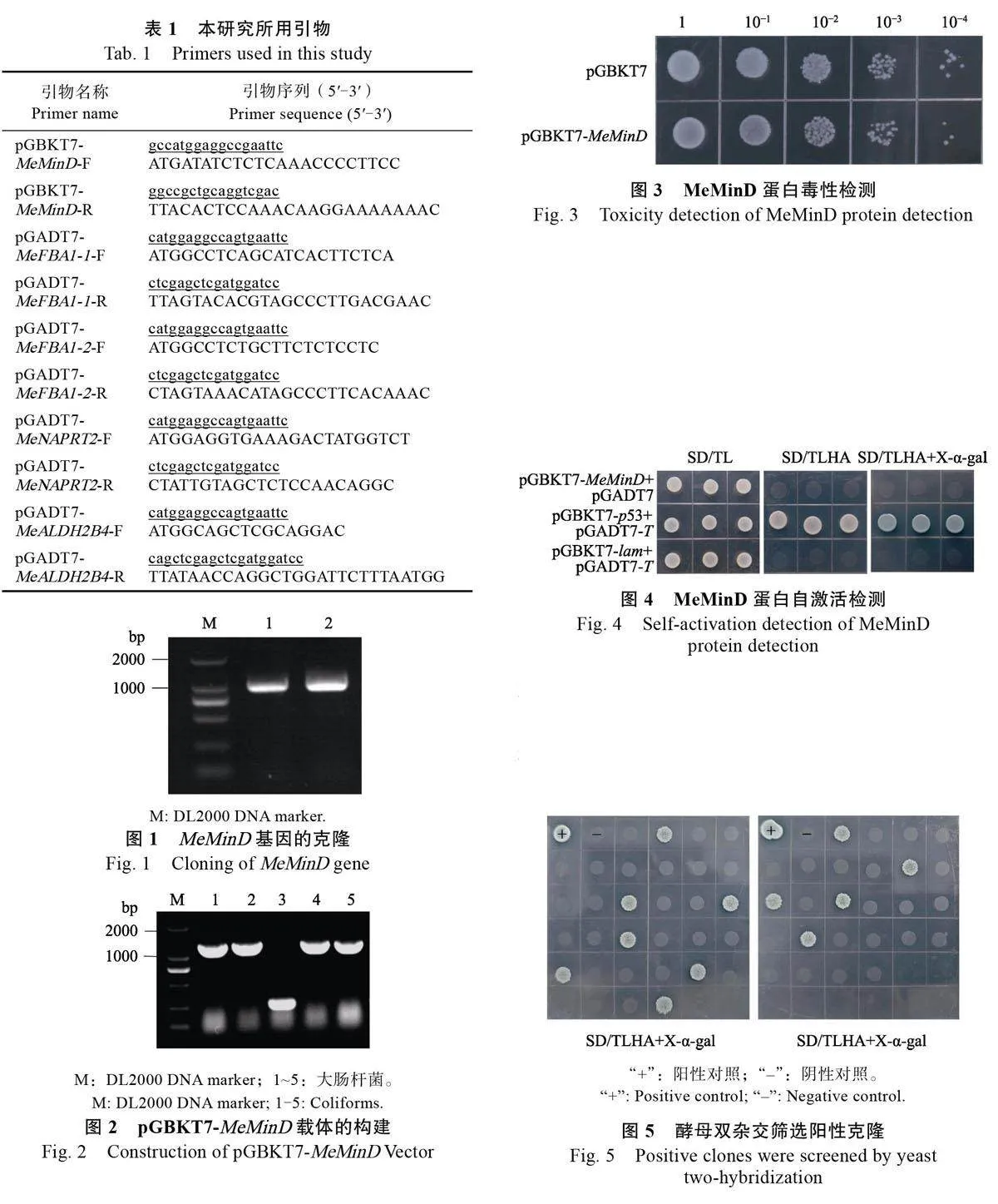
將本實驗室前期制備的木薯酵母雙雜交質粒文庫大規模轉化到含有誘餌載體pGBKT7-MeMinD 的AH109 酵母感受態。轉化后的酵母菌液涂布于SD/TLHA 固體培養基上培養,獲得單克隆酵母菌,并分別點在SD/TLHA+X-α-Gal 培養基上進行培養(圖5)。將變藍的陽性菌落用于PCR擴增插入片段并測序,最終獲得8 個候選互作蛋白(表2)。這些蛋白功能涉及卡爾文循環、細胞代謝、NAD(P)H 再生和細胞熱耐受等多種生物學過程。
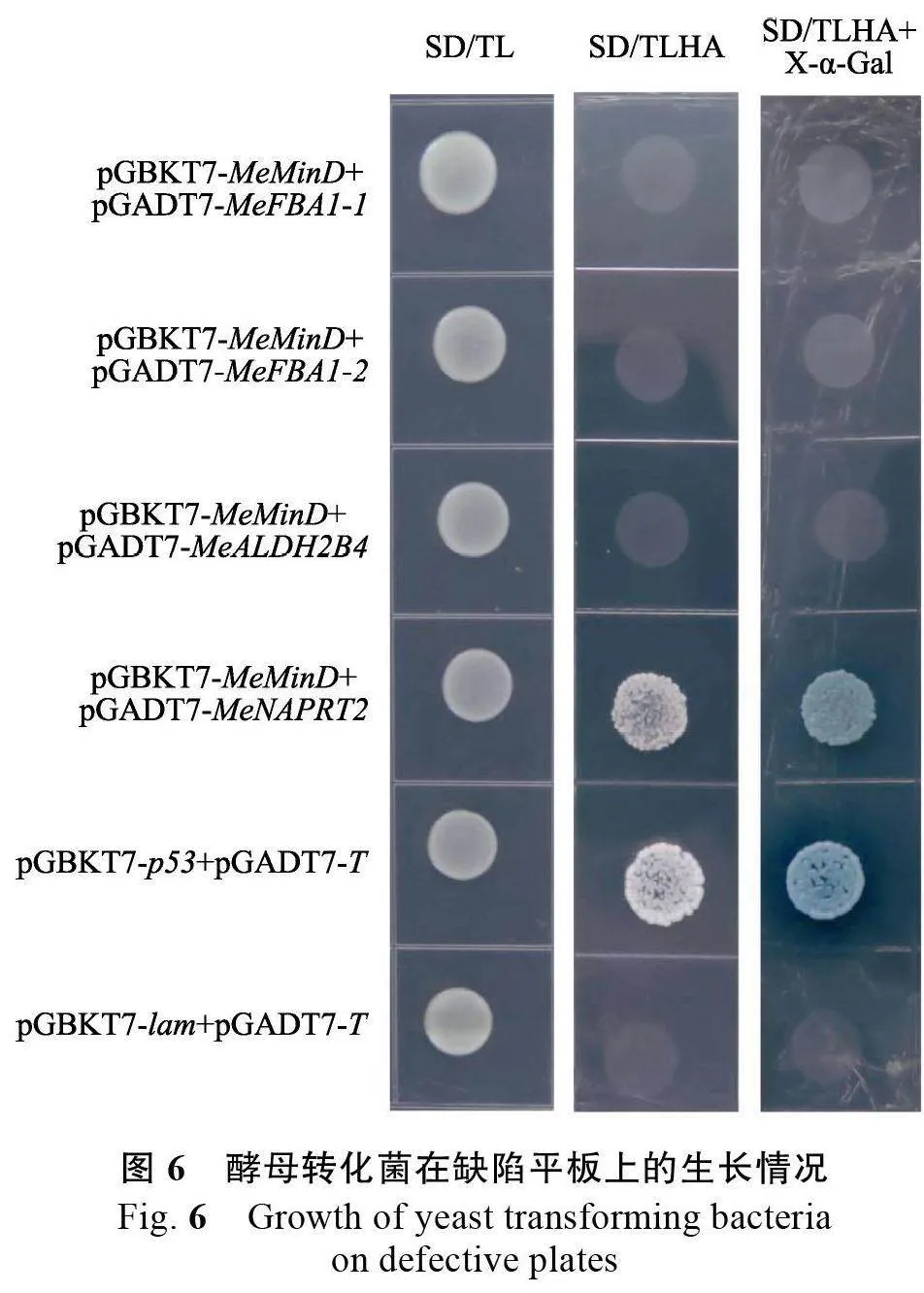
2.4 MeMinD 互作蛋白點對點驗證
根據候選蛋白的注釋結果,挑選出4個候選蛋白進行酵母雙雜交點對點驗證,包括MeFBA1-1、MeFBA1-2、MeNAPRT2、MeALDH2B4。以SC8 木薯品種cDNA 為模板,PCR 擴增出4 個候選互作蛋白基因的CDS 序列,將其分別構建到pGADT7 載體。將誘餌載體和獵物載體共轉酵母。結果顯示,pGBKT7-MeMinD+pGADT7-MeFBA1-1、pGBKT7-MeMinD+pGADT7-MeFBA1-2、pGBKT7-MeMinD+pGADT7-MeALDH2B4、pGBKT7-MeMinD+pGADT7-MeNAPRT2 以及對照組在SD/TL 上均可以長出菌斑并正常生長(圖6),說明質粒共轉化酵母成功。酵母雙雜交驗證結果顯示,僅有pGBKT7-MeMinD+pGADT7-MeNAPRT2 能在SD/TLHA 上生長出菌斑且在SD/TLHA+X-α-Gal 上變藍,表明在酵母細胞內MeMinD 與MeNAPRT2 蛋白存在相互關系。
3討論
淀粉體和葉綠體均屬于質體,由前質體發育而來,具有相似的分裂機制。植物質體的分裂過程,包括FtsZ 蛋白組裝形成Z 環,MinD1 和MinE1互作形成復合體,Min 復合體與ARC3 蛋白協同調控Z 環精準定位于質體的中部,Z 環形成后招募更多的質體分裂相關蛋白,Z 環在多種蛋白的作用下引導質膜收縮,最終導致質體分裂。過量或減少擬南芥AtMinD1 的表達都會影響葉綠體的分裂,從而影響葉綠體的大小[9]。
木薯塊根富含的淀粉和葉片臨時性淀粉的合成和貯存均在質體完成,挖掘鑒定木薯質體分裂相關基因的功能,解析淀粉體和葉綠體的分裂機制將有助于提高木薯產量和淀粉品質。GENG 等[10]克隆了木薯FtsZ 蛋白的3 個基因,MeFtsZ1、MeFtsZ2-1 和MeFtsZ2-2。MeFtsZ 蛋白均能定位在葉綠體中,并形成Z 環結構。MeFtsZ2-1 和MeFtsZ2-2 在塊根的表達量顯著高于MeFtsZ1。過表達MeFtsZ2-1 和MeFtsZ2-2 的擬南芥葉綠體增大,數目減少。WANG 等[11]克隆了木薯MeMinE基因,MeMinE-GFP 在木薯原生質體中的瞬時表達表明,MeMinE 蛋白位于葉綠體中。MeMinE蛋白在木薯葉肉細胞原生質體中的過表達,可導致葉綠體分裂異常,葉綠體數目減少,面積變小。KE 等[7]克隆了木薯的MeMinD 基因,該基因在葉片中的表達量較高,MeMinD 定位于葉綠體,在大腸桿菌異源表達可導致菌體的分裂位點錯位和絲狀形成。通過基因編輯敲除MeMinD 基因,突變體木薯的葉綠體和淀粉體顯著增大(數據待發表)。解析MeMinD 參與調控木薯質體分裂的機制,將有助于利用分子生物技術培育高產、高淀粉木薯新品種。
目前,與MeMinD 蛋白互作,協同調控木薯質體分裂的蛋白尚未明確。本研究采用酵母雙雜交技術對MeMinD 進行了互作蛋白的篩選,發現MeMinD 與MeNAPRT2 存在互作關系。NAPRT2(nicotinate phosphoribosyl transferase 2)在植物中參與煙酸(nicotinic acid)的磷酸核糖基轉移反應,這是煙酸轉化為NAD(煙酰胺腺嘌呤二核苷酸)途徑中的關鍵步驟[12]。NAD 是細胞內重要的輔酶,參與多種代謝過程,包括能量代謝和信號傳導[13-15]。NAD+(NAD 的氧化形式)在氧化還原反應中接受電子和質子,形成NADH。NADH隨后在電子傳遞鏈中將電子傳遞給氧氣,最終產生ATP。MinD 蛋白是一種ATPase,通過ATP 的水解,MinD 蛋白能夠在質膜上形成動態的極性模式,與MinE 和MinC 蛋白相互作用,共同調控質體的分裂過程。MeMinD 與MeNAPRT2 的互作表明能量代謝可能參與木薯質體的分裂過程。水稻OsNAPRT1 基因突變會導致植株矮化,葉片過早枯萎[16]。目前,尚無NAPRT2 參與質體分裂的研究報道。后續將通過雙分子熒光互補試驗進一步驗證MeMinD 與MeNAPRT2 的互作關系,利用轉基因或基因編輯技術探究其在木薯質體分裂過程中的功能。
