樹木伯克霍爾德氏菌DHR18誘導橡膠樹抗病性相關防御酶分析
2024-12-31 00:00:00羅佑紅趙欣陽蔡海濱孟祥佳周燚涂敏
熱帶作物學報 2024年10期


關鍵詞:伯克霍爾德氏菌;橡膠樹;膠孢炭疽菌;誘導抗病性;防御酶
中圖分類號:S794.1 文獻標志碼:A
天然橡膠是一個國家至關重要的戰略資源以及不可替代的工業原料,而全世界98%的天然橡膠是由橡膠樹(Hevea brasiliensis Muell. Arg.)生產[1]。橡膠樹種植受到獨特的地域和氣候限制,其中影響橡膠產量的重要因素之一是生物脅迫[2]。炭疽病是橡膠樹生產中的重要病害,膠孢炭疽菌復合群中的Colletotrichum siamense 是我國植膠區的田間優勢致病種,通過侵染橡膠樹的枝條、花、葉等組織,造成葉片枯萎和落葉,最終導致嚴重減產[3]。目前,主要防治方法是使用化學藥劑,由于橡膠樹屬高大喬木,在防治過程中使用藥劑比較單一、藥量大、易發生漂移,且化學藥劑在防治時存在抗藥性、防治成本高和環境污染等問題[4-5]。因此,急需一種綠色環保、經濟安全的手段來防治橡膠樹炭疽病。
生物防治是通過利用有益微生物或其代謝物來防治植物病害,已經成為病害防控中一種可持續和有效的方法[6-7]。生物防治劑不僅可以直接抑制植物病原體的生長來減少對植物的侵染,還能誘導植物的系統抗性,引發植物廣泛的防御反應,對多種病原體有著長期的防治效果[8]。眾多研究表明,生防菌可以通過誘導植物相關防御酶的升高,從而增強植物的抗性[9]。內生耐鹽芽孢桿菌Cal.l.30 和Cal.f.4 可以誘導番茄對灰葡萄孢的系統抗性,從而有效防治番茄灰霉病[10]。羅氏假單胞菌GC-7 處理的植物根系中過氧化氫酶(catalase,CAT)、多酚氧化酶(polyphenol oxidase,PPO)、苯丙氨酸解氨酶(phenylalanin ammonialyase,PAL)等防御相關酶的活性顯著增加,從而增強植物抗性[11]。
伯克霍爾德氏菌(Burkholderia sp.)是一種能有效防治多種植物病害的生防菌,與植物有著密切的共生關系,而且伯克霍爾德氏菌產生的一些次級代謝產物,對大部分植物病原真菌有明顯抑制作用。此外,可以通過誘導植物防御相關酶活性的升高來增強對植物病原菌的抗性[12]。已有研究發現,越南伯克霍爾德氏菌C12 通過誘導水稻防御相關酶活性(CAT、PAL、SOD、PPO 和POD)的升高,從而增強水稻的抗病性[13];吡咯伯克霍爾德氏菌S17-377 可誘導水稻防御相關酶活性變化,有3 種防御相關酶的活性增強,說明伯克霍爾德氏菌可以增強水稻對水稻紋枯病的抗性[14]。本實驗室前期分離得到1 株對橡膠樹膠孢炭疽菌有較強拮抗作用的樹木伯克霍爾德氏菌(B. arboris),本研究系統測定了不同濃度樹木伯克霍爾德氏菌DHR18 發酵液以及DHR18 與膠孢炭疽菌協同處理下橡膠樹葉片中CAT、POD、PAL、SOD 和PPO 活性變化,初步探索樹木伯克霍爾德氏菌誘導橡膠樹對膠孢炭疽菌的抗性機理,以期為橡膠樹炭疽病的綠色防控提供科學依據。
1 材料與方法
1.1 材料
樹木伯克霍爾德氏菌株 DHR18 及橡膠樹膠孢炭疽菌CH-1 均由中國熱帶農業科學院橡膠研究所種質資源實驗室分離、鑒定及保存,其中樹木伯克霍爾德氏菌DHR18 分離自中國熱帶農業科學院試驗場(海南儋州)橡膠樹根病區;橡膠樹膠孢炭疽菌CH-1 分離自國家橡膠種質資源圃(儋州)。
供試橡膠種苗品種為GT1,挑選葉部生長健康、苗長35 cm,胸徑和株高等長勢均一的具1 篷葉淡綠期葉片的種子苗,設置DHR18 發酵液噴施3個濃度處理,以及病原菌與生防菌協同噴施3 個處理,每個處理使用種子苗20 株,每個處理重復5次,所有種子苗均由國家橡膠種質資源圃提供。
1.2方法
1.2.1 DHR18發酵液及膠孢炭疽菌CH-1孢子懸浮液制備挑取 DHR18單菌落于100 mL LB 培養基中,在恒溫搖床(180 r/min,28 ℃)中培養24 h,將DHR18 發酵液濃度調節至1×108 CFU/mL作為原液,用無菌水稀釋得到1×107 CFU/mL 和1×106 CFU/mL 濃度的稀釋液。膠孢炭疽菌CH-1接種在PDB 培養基中,在恒溫搖床(180 r/min,28 ℃)中培養3d待用。炭疽菌接種:參考蔡志英等[15]的方法,用孢子懸浮液(1×106 個/mL)接種,預先用無菌昆蟲針刺傷橡膠葉片,孢子懸浮液用噴霧器均勻噴施,直到橡膠葉片掛滿水滴為止,每個處理重復3 次;空白對照噴施適量無菌水,套袋保濕。
1.2.2 不同濃度的DHR18 發酵液處理橡膠樹參考唐文等[16]的方法,共設置3 個濃度的DHR18發酵液誘導處理(表1),包括1×108、1×107、1×106 CFU/mL 處理,以LB 培養基為對照,取10 mL不同濃度的DHR18 發酵液和對照噴灑植株葉面,每個處理重復5 次。分別于處理后的24、48、72、96、120 h,取位置相同的橡膠樹葉片,用于防御酶的活性測定。
1.2.3 DHR18 與CH-1 協同處理橡膠樹共設置4個處理,處理方式如表2所示。樣品采集和處理與1.2.2 相同。養基中,在恒溫搖床(180 r/min,28 ℃)中培養24 h,將DHR18 發酵液濃度調節至1×108 CFU/mL作為原液,用無菌水稀釋得到1×107CFU/mL 和1×106 CFU/mL 濃度的稀釋液。膠孢炭疽菌CH-1接種在PDB 培養基中,在恒溫搖床(180 r/min,28 ℃)中培養3d待用。炭疽菌接種:參考蔡志英等[15]的方法,用孢子懸浮液(1×106 個/mL)接種,預先用無菌昆蟲針刺傷橡膠葉片,孢子懸浮液用噴霧器均勻噴施,直到橡膠葉片掛滿水滴為止,每個處理重復3 次;空白對照噴施適量無菌水,套袋保濕。
1.2.2 不同濃度的DHR18 發酵液處理橡膠樹參考唐文等[16]的方法,共設置3 個濃度的DHR18發酵液誘導處理(表1),包括1×108、1×107、1×106 CFU/mL 處理,以LB培養基為對照,取10 mL不同濃度的DHR18 發酵液和對照噴灑植株葉面,每個處理重復5 次。分別于處理后的24、48、72、96、120 h,取位置相同的橡膠樹葉片,用于防御酶的活性測定。
1.2.3 DHR18與CH-1協同處理橡膠樹共設置4個處理,處理方式如表2所示。樣品采集和處理與1.2.2相同。
1.2.4 橡膠樹防御酶活性測定將 1.2.2 和1.2.3中采集的橡膠樹葉片,使用酶活試劑盒(索萊寶科技有限公司)測定POD、SOD、CAT、PAL 和PPO 活性。按照試劑盒說明書中步驟加入試劑,在相應的波長下測定其吸光值,并按照試劑盒的公式計算其酶活力。
1.3 數據處理
實驗數據采用Excel 2016軟件進行統計、整理,采用IBM SPSS Statistics 25軟件進行統計分析。
2 結果與分析
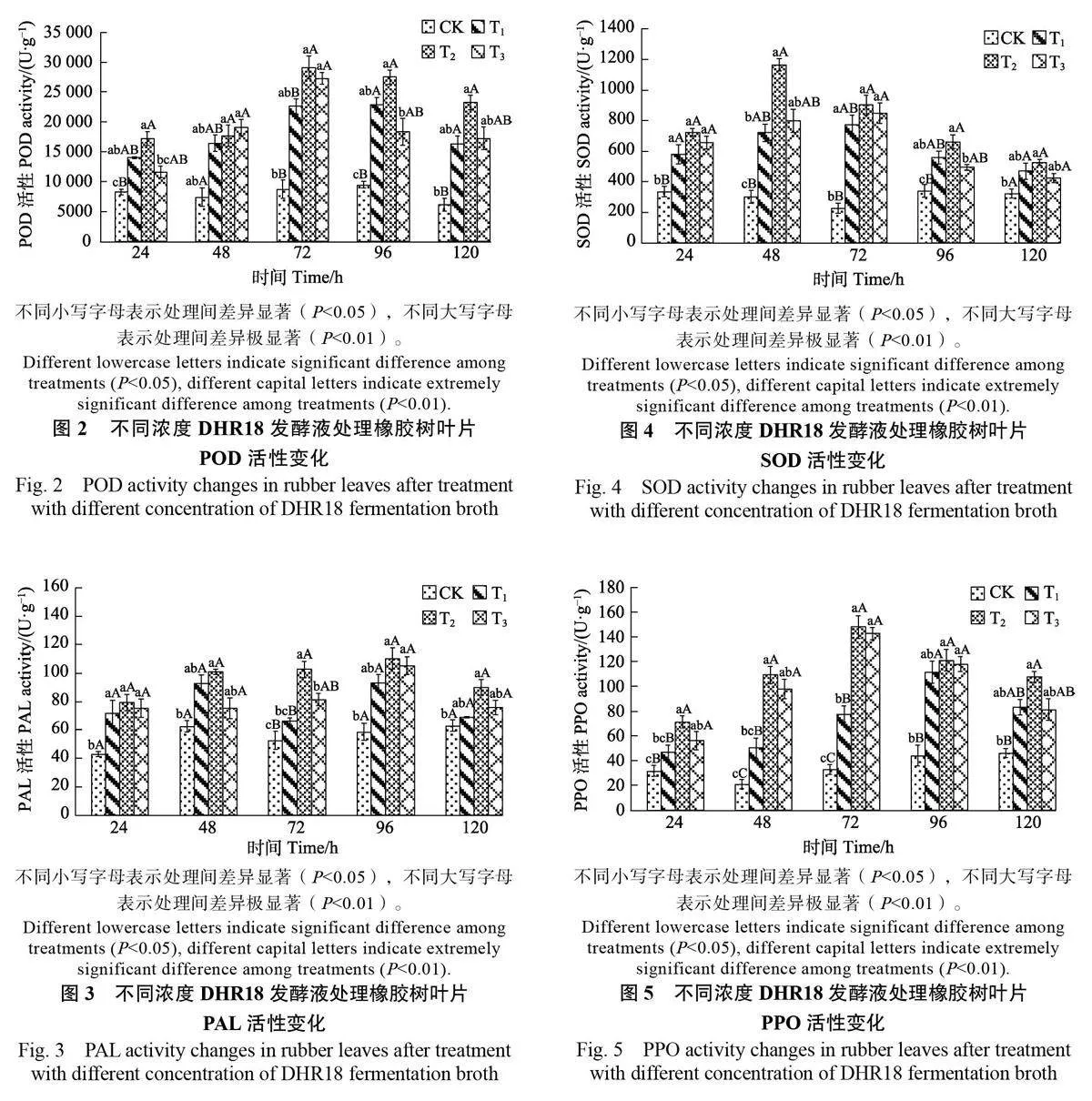
2.1 不同濃度DHR18發酵液處理對橡膠樹防御酶活性的影響
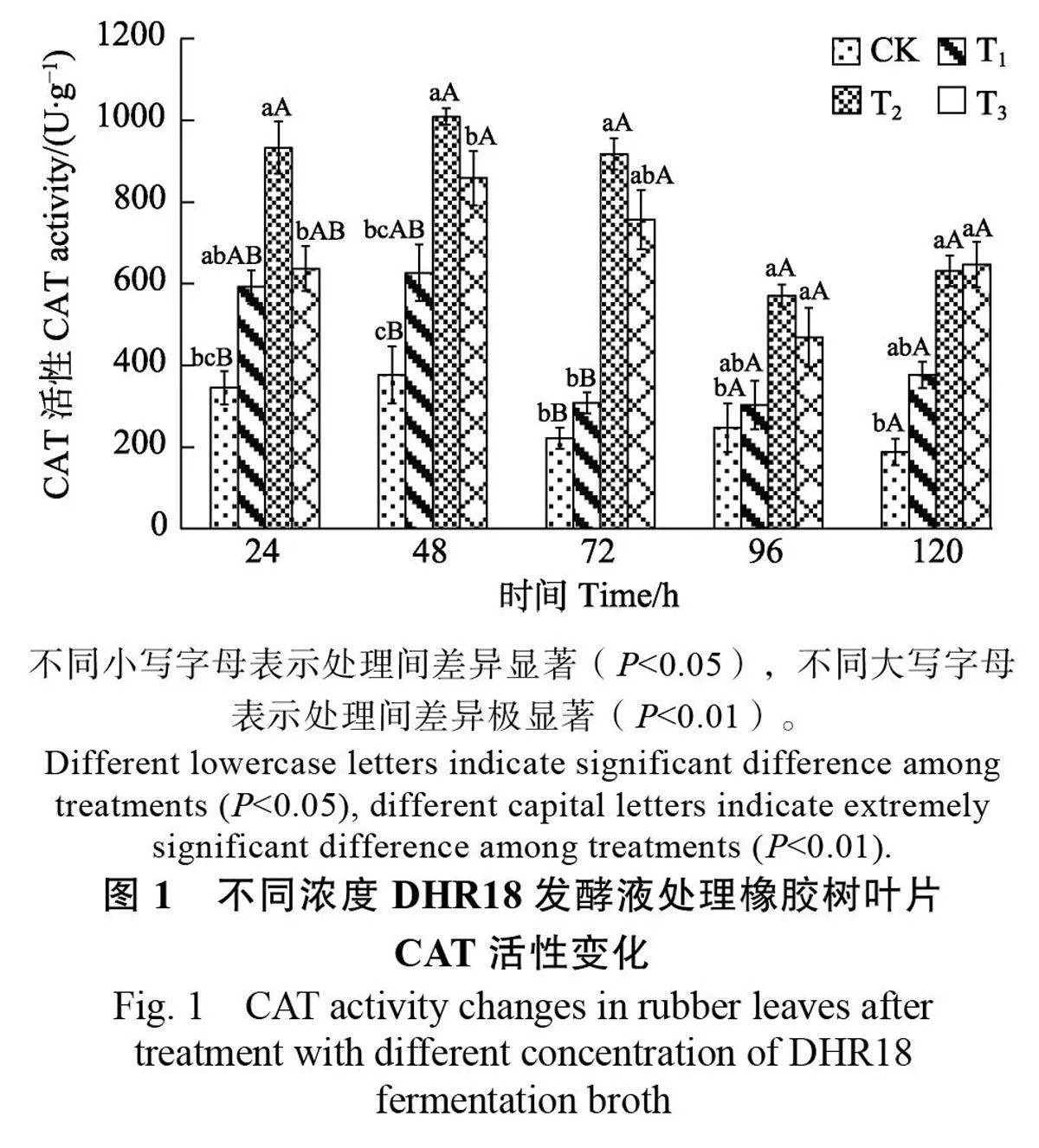
2.1.1 過氧化氫酶活性變化 經不同濃度 DHR18發酵液處理后,橡膠樹葉片中CAT 活性均高于CK。其中,T2、T3處理的CAT 活性呈先升高后降低再升高的趨勢,且在48h 時達到峰值,其活性分別為1009.14 U/g 和858.79 U/g,為CK 的2.68倍和2.29倍,均極顯著高于CK(Plt;0.01);而T1的CAT 活性在48 h 后下降并趨于穩定,但在同時間的酶活性均高于CK(圖1)。
2.1.2 過氧化物酶活性變化 如圖2所示,經不同濃度DHR18 發酵液噴施葉片處理后,處理組葉片 POD 活性均呈先升高后降低的趨勢,并且活性在相同時間均高于CK。在72h時,每個處理組葉片POD活性達到峰值,此時T2和T3處理的POD酶活性達到29 138.67、27 276.67 U/g,分別是CK的3.35倍和3.13倍,均極顯著高于CK(Plt;0.01)。
2.1.3 苯丙氨酸解氨酶活性變化 如圖3所示,T2處理的PAL 活性呈先升高后降低的趨勢,在96 h時,其PAL 活性達到峰值110.24 U/g,是CK 的1.88倍,顯著高于CK(Plt;0.05)。T1 處理的PAL 活性呈先升高后降低再升高的趨勢。在48 h 和96 h 時出現峰值,葉片PAL 活性分別為92.72 U/g 和93.43 U/g,均高于CK。T2 處理效果最明顯,在相同時間,其處理的葉片PAL 活性均高于其他濃度。
2.1.4 超氧化物歧化酶活性變化 如圖4所示,經不同濃度DHR18 發酵液噴施葉片處理后,處理組葉片SOD活性呈先升高后降低的趨勢,且其活性在相同時間均高于CK。T2 處理的SOD 活性在48h 達到峰值1164.89 U/g,是CK 的3.83倍,極顯著高于CK(Plt;0.01)。T1 和T3 處理的SOD活性在72 h 達到峰值,分別為774.93U/g 和848.18 U/g,是CK 的3.41 倍和3.73倍,顯著(Plt;0.05)和極顯著高于CK(Plt;0.01)。
2.1.5 多酚氧化酶的活性變化 由圖 5 可知,T1、T2 和T3處理的PPO活性呈先升高后降低的趨勢,其活性在相同時間均高于CK。T2、T3處理的PPO 活性在72 h 時分別達到峰值148.00 U/g和142.80 U/g,是CK 的4.51 倍和4.35 倍。T1 處理的PPO 活性在96 h 時達到峰值(111.60 U/g)。經不同濃度DHR18 發酵液處理的葉片PPO 活性在峰值時均極顯著高于CK(Plt;0.01)。
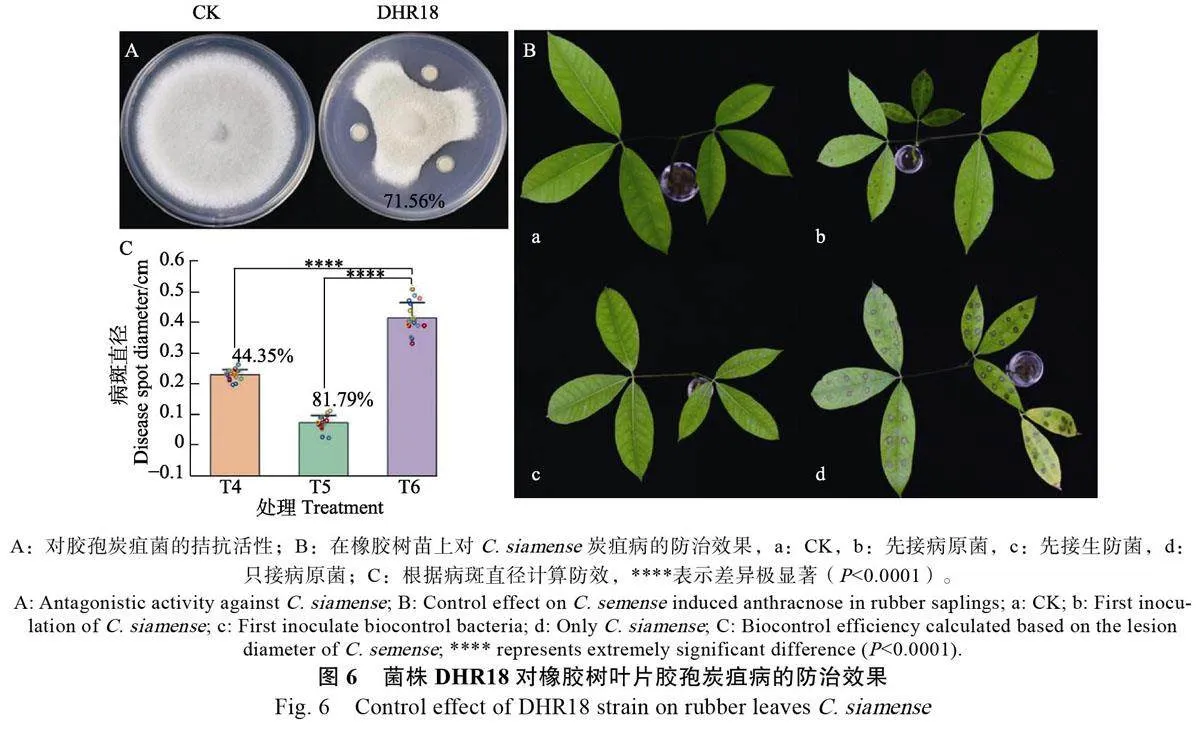
2.2 菌株DHR18對膠孢炭疽病的防治效果
如圖6所示,菌株DHR18在平板上對橡膠樹膠孢炭疽菌CH-1 表現出明顯的抑制效果,抑制率為 71.56%。按照表2,使用DHR18 與CH-1 協同處理橡膠樹幼苗,72 h 后,CK 的葉片未發病,T5處理(先接生防菌)的防治效果為81.79%,T4處理(先接病原菌)的防治效果為44.35%,均顯著高于T6處理(只接病原菌)(Plt;0.0001)。
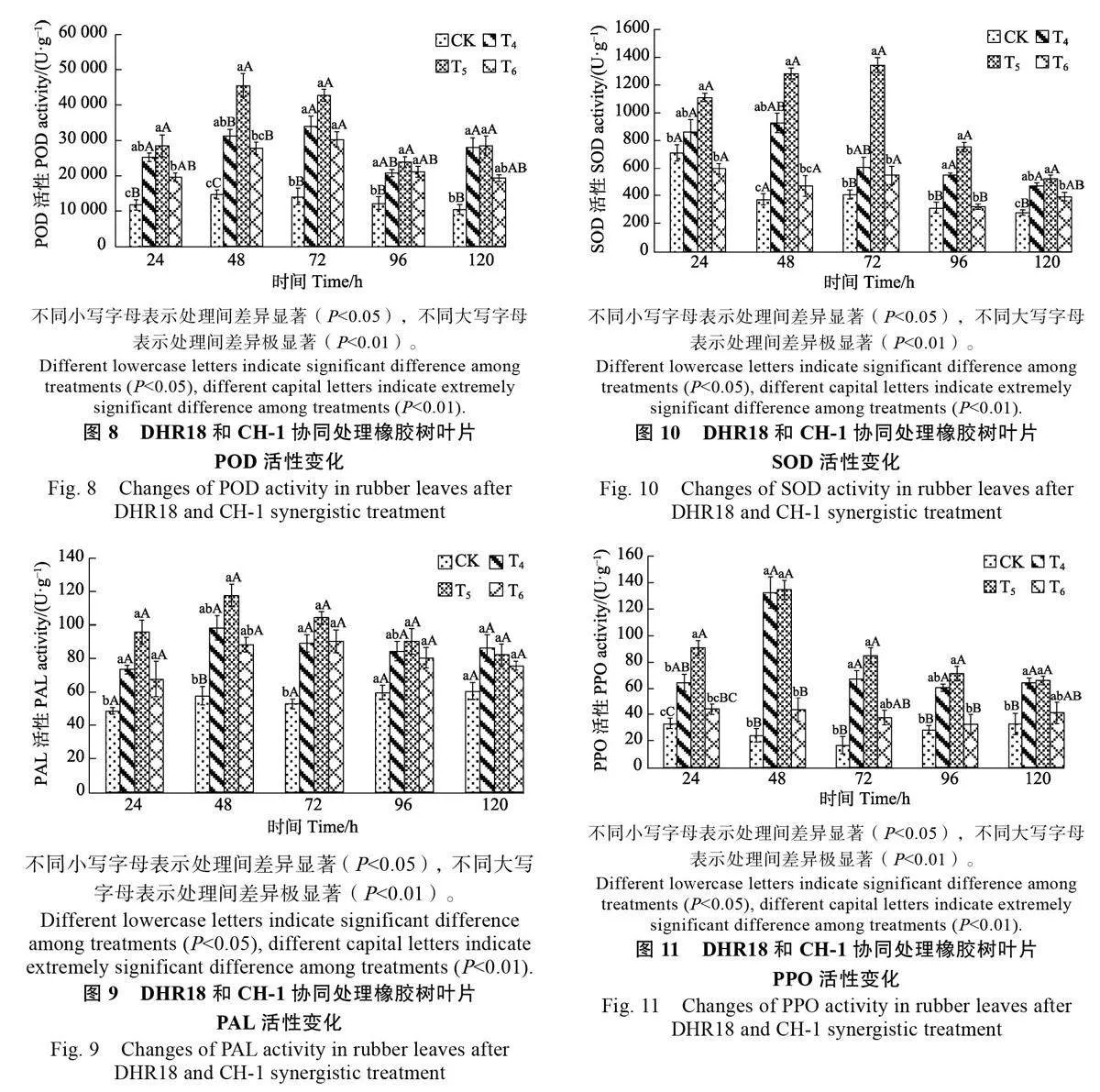
2.3 DHR18 與CH-1 協同處理對橡膠樹相關防御酶活性的影響
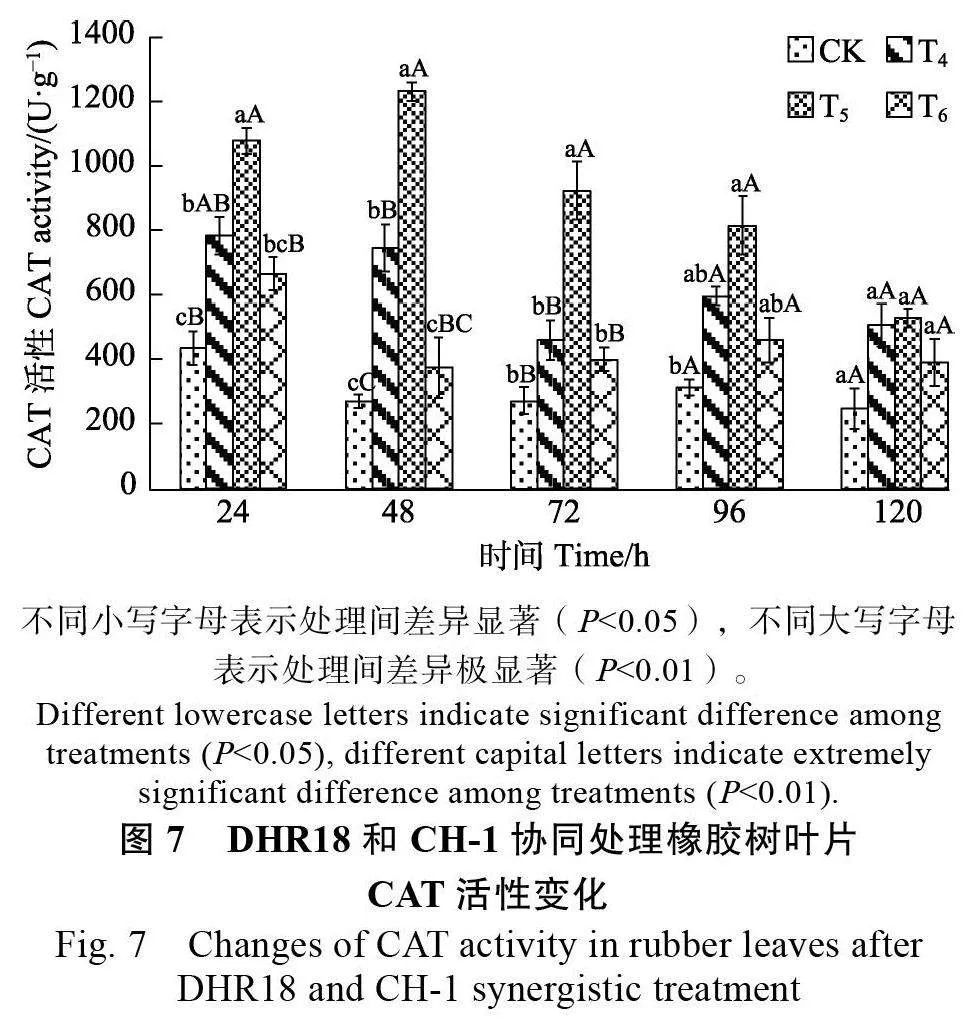
2.3.1 氧化氫酶活性變化 生防菌DHR18 和CH-1 共同處理后橡膠樹的相關防御酶CAT 活性有顯著變化,3種不同處理組CAT 活性在同時間均高于CK。其中,T5 處理的CAT 活性呈先升高后降低的趨勢,在48 h 時達到峰值(1230.85 U/g),極顯著高于其他處理(Plt;0.01),是CK 的4.56倍;T4 處理的CAT 活性呈先降低后升高再降低的趨勢,在48 h 極顯著高于CK(Plt;0.01)(圖7)。
2.3.2 過氧化物酶活性變化" DHR18 與CH-1 協同處理下,3種處理組的POD活性均高于CK,在同時間均達顯著水平(Plt;0.05),其中,T5 的POD 活性在48 h 時達到峰值(45 504.67 U/g),是CK 的3.10 倍;T4 和T6 處理的POD 活性在72h 時達到峰值(30 249.33、33875.33 U/g),分別為CK 的2.14倍和2.40倍。3種不同處理在POD活性峰值時均極顯著高于CK(Plt;0.01)(圖8)。
2.3.3 苯丙氨酸解氨酶活性變化 由圖9可知,T5處理的PAL 活性總體呈先升高后降低的趨勢,在同時間均高于CK,72 h 時與CK 差異顯著(Plt;0.05),在48 h 時達到峰值(117.53 U/g),是 CK 的2.04 倍,極顯著高于CK(Plt;0.01);T4 和T6 處理的PAL 活性變化不明顯,活性保持在85.00 U/g 左右,在72 h 顯著高于CK(Plt;0.05)。
2.3.4 超氧化物歧化酶活性變化 由圖 10 可知,T5 處理的SOD 活性在72 h 極顯著高于CK(Plt;0.01),SOD 活性達1342.17 U/g,是CK 的3.30 倍;T4 的SOD 活性呈先升高后降低的趨勢,在48 h 達到峰值,活性為924.49 U/g,是CK 的2.47 倍,顯著高于CK(Plt;0.05);T6 的SOD 活性在24 h 時低于CK,與CK 無顯著差異。
2.3.5 多酚氧化酶活性變化 由圖 11 可知,T5的PPO 活性呈先升高后降低的趨勢,在同時間均高于 CK,且在48 h 時達到峰值,為134.40 U/g,是CK 的5.6 倍;T4 的PPO 活性在48 h 時達到峰值,為134.40 U/g,是CK 的5.51 倍,均極顯著高于對照CK(Plt;0.01);T6 的PPO 活性變化不明顯,未出現明顯的峰值,活性略高于CK。
3 討論
過氧化氫酶(CAT)[17]、過氧化物酶(POD)和超氧化物歧化酶(SOD)是植物主要的活性氧清除酶,可消除因病害侵染或逆境脅迫產生過多的活性氧,從而避免對自身造成損傷[18];苯丙氨酸解氨酶(PAL)可催化苯丙氨酸合成酚類化合物,而酚類化合物是植物多種次生代謝產物的中間產物,對植物抗病蟲害有重要作用[19];多酚氧化酶(PPO)是一類金屬蛋白酶,可以催化多種生化反應,調節生理功能,與植物抗逆抗病等息息相關[20]。這些防御相關酶活性對于植物的抗病性具有重要決定作用,也是評價植物抗性水平的重要參考標準[21-22]。
研究表明,生防菌通過誘導和增強一系列防御相關酶活性,從而提高植物的系統抗病能力,是其重要的抗病機制之一[23-24]。CHEN 等[25]研究發現植物乳桿菌CM-3 可以通過增加葡萄POD、PPO、CAT 和PAL 的活性來增強葡萄的抗病能力。鄒強等[26]用生防貝萊斯芽孢桿菌TP-1 處理后葡萄的APX、PPO 和PAL 活性提高,增強了對灰霉菌的抗性。本研究中,生防菌DHR18 顯著提高橡膠樹中CAT、POD、SOD、PAL 和PPO 五種防御酶活性,增強了橡膠樹的抗性,其中多酚氧化酶的變化最為明顯,最高可以達到對照的4.51倍。陳秀香等[27]研究發現,經枯草芽孢桿菌FC21的10倍稀釋液處理后的植株葉片中CAT、POD、SOD活性最高,生防菌誘導的防御相關酶活性與菌液濃度不呈正相關。本研究發現107 CFU/mL 濃度的DHR18 發酵液對橡膠樹防御誘導效果最好,可作為伯克霍爾德氏菌DHR18 防治炭疽病的濃度參考。因此,本研究結果表明,樹木伯克霍爾德氏菌DHR18 能誘導橡膠樹體內CAT、POD和SOD酶活增強來清除過量活性氧;能誘導PAL 和PPO酶活性促使橡膠樹產生抗菌物質等,從而提高橡膠樹系統抗性。
植物在受到病原物侵染后,活性氧作為植物抵御和延緩病原物侵染的第一道物理屏障,在第一時間反應并影響活性氧消除酶的合成酶基因的表達,從而增加CAT、POD 和SOD 等的活性繼而實現解毒作用,更好地應對各種脅迫,其體內防御酶活性也會上升[28]。感染病原物后,活性氧消除酶普遍呈先上升后下降的變化趨勢。耿莉娜等[29]的研究發現煙草的抗病品種Beinhart1000-1在接種赤星病菌后,POD、PAL、SOD 活性明顯高于對照,呈先升高后降低的變化趨勢。本研究中,在接種炭疽病CH-1后,橡膠樹的SOD和PAL活性無明顯變化,可能與植株品種抗病性有關。羅富方等[30]研究發現在青枯病脅迫下,烤煙再接種印度梨形孢,其4 種防御酶(PAL、PPO、POD和SOD)活性增加均高于對照,且呈先升高后降低的趨勢。本研究中DHR18和CH-1協同處理下,處理組的防御酶(CAT、POD、SOD、PAL 和PPO)活性均有提高,主要呈先上升后下降的趨勢;協同處理的CAT、SOD 和PPO酶活性在24、48、72 h 顯著高于其只接病原菌的酶活,表明伯克霍爾德氏菌DHR18 在橡膠樹受到炭疽病菌侵染過程中會增強其活性氧清除系統,并促進合成抗病物質,從而增強植株的抗病能力。唐文等[16]先噴施枯草芽孢桿菌Czk1 后接種病原菌處理的5 種防御相關酶(CAT、POD、PAL、SOD 和PPO)活性升高,顯著高于其他處理。本研究橡膠樹受到炭疽病侵染前噴施DHR18發酵液,對防御酶活性影響最大,5 種防御酶活性高于其他處理組,更有利于誘導抗病性。本研究發現,先噴施生防菌伯克霍爾德氏菌DHR18 再接種病原菌處理組的POD活性比只接病原菌和先接病原菌處理組的更快達到峰值,在48 h 時顯著高于其他處理組,能更快引起植物的防御反應;周東興等[31]研究發現貝萊斯芽孢桿菌DX-25與番茄枯萎病互作的情況下可以更快地引起植株番茄的抗性反應,與本研究結果相似。
在本研究中,生防菌DHR18 發酵液可促使葉片中CAT、POD、PAL、SOD 和PPO 酶活性上升,誘導橡膠樹產生系統抗性,1×107 CFU/mL 濃度對橡膠樹的誘導效果最好。研究結果初步明確了樹木伯克霍爾德氏菌具有誘導抗病性,為探索伯克霍爾德氏菌誘導抗病性提供了部分理論依據。
