全硫代反義寡核苷酸對卵巢癌細胞生長抑制作用
2009-05-06 03:35:52劉學單偉曾瑞霞房艷李德華秦書儉
中國實用醫藥 2009年8期
劉 學 單 偉 曾瑞霞 房 艷 李德華 秦書儉
【摘要】 目的 采用針對人端粒酶hTR基因的反義寡聚脫氧核苷酸,探討端粒酶反義寡聚脫氧核苷酸(antisense oligodeoxy nucleotides,ASODN)對卵巢癌HO-8910細胞端粒酶活性及細胞增殖的影響。方法 ①將實驗分為空白對照組、脂質體對照組、端粒酶全硫代正義寡聚脫氧核苷酸(Phosphorathioate sense oligodeoxy nucleotides,PS-SODN)組和不同劑量的全硫代反義寡聚脫氧核苷酸(Phosphorat-hioate antisense oligodeoxy nucleotides,PS-ASODN)組;②脂質體介導的細胞轉染后,PS-ASODN和PS-SODN分別作用于卵巢癌HO-8910細胞后并培養24、48、72 h,分別采用酶聯免疫吸附法(ELISA)、吖啶橙染色法、四甲基偶氮唑藍比色法(MTT)、流式細胞術檢測HO-8910細胞的端粒酶活性、細胞形態、體外增殖、細胞凋亡和細胞周期的改變。結果 ①ELISA法檢測結果顯示端粒酶PS-ASODN作用卵巢癌HO-8910細胞72 h后端粒酶活性表達為陰性,說明端粒酶PS-ASODN能夠抑制端粒酶活性;②吖啶橙染色觀察細胞形態:PS-ASODN作用的卵巢癌HO-8910細胞有明顯凋亡現象,凋亡細胞體積縮小,染色質濃縮,說明PS-ASODN能夠促進卵巢癌HO-8910細胞凋亡;③MTT實驗:PS-ASODN明顯抑制卵巢癌HO-8910細胞的增殖(P<0.0l ),并呈一定劑量和時間依賴關系;④流式細胞儀檢測細胞凋亡和周期:與空白對照組比較,PS-ASODN組G0/G1期細胞明顯增多,差異顯著(P<0.01);在G0/G1期前出現亞二倍體凋亡峰,表明細胞被阻止在G1/S期。結論 ①PS-ASODN作用于卵巢癌HO-8910細胞后,卵巢癌HO-8910細胞增殖受到明顯抑制,并出現凋亡;②PS-ASODN對端粒酶活性的抑制率與PS-ASODN的濃度和作用時間呈依賴關系,即抑制率隨著反義寡核苷酸的濃度和作用時間的增加而增大;③以端粒酶RNA模板區為靶點的PS-ASODN明顯抑制卵巢癌HO-8910細胞的增殖,其機制可能是通過降低細胞的端粒酶活性而誘發細胞的凋亡,PS-ASODN對卵巢癌的治療具有重要價值。
【關鍵詞】
端粒酶全硫代反義寡聚脫氧核苷酸;卵巢癌HO-8910細胞;端粒酶;細胞凋亡
Effects of telomerase antisense oligodeoxy nucle-otides on HO-8910 ovary carcinoma cell
LIU Xue,SHAN Wei,ZENG Rui-xia,et al.Department of Anatomy,Liao Ning Medical College,Jinzhou 121001,China
【Abstract】 Objective In this study,we apply Oligonucleotide aimed directly at human telomerase RNA(hTR),To study the effects of telomerase Phosphorathioate antisense oligodeoxy nucleotides (PS-ASODN) on telomerase activity and proliferation of HO-8910 ovary carcinoma cell.Methods ①HO-8910 ovary carcinoma Cell was transfected by PS-ASODN,Phosphorathioate sense oligodeoxy nucleotides(PS-SODN)mediated by Lipotap Liposomal Transfection Reagen.The experiments were classified into normal group、PS-ASODN groups at varied concentrations 、PS-SODN group and oligofectamineTM alone.②The proliferation activity of HO-8910 ovary carcinoma cell line was determined by using methyl thiazolyl tetrazolium assay.The telomerase activity was determined by using enzyme-linked immunosorbent assay.The cell morphology was observed by fluorescence microscope stained with acridine orange.Flow cytometry was adopted to examine apoptotic rate and cell cycle.Results ①With ELISA,Telomerase of HO-8910 ovary carcinoma cell was repressed by telomerase PS-ASODN after 72 h later,it showed that telomerase PS-ASODN could lead inhibition of telomerase activity.②The cell morphology was observed by fluorescence microscope stained with acridine orange:Apoptotic morphological characteristic of HO-8910 ovary carcinoma cell transfected by antisense oligonucleotides was observed.Moreover,the apoptotic cell physical volume contracts.The HO-8910 ovary carcinoma cell growth was inhibited by Phosph-orathioate antisense Oligonucleotide and appeared the apoptosis.③PS-ASODN caused significant (P<0.01) inhibition of cell growth,and the rate of inhibition has the discrepancy between the different dose and different time (P<0.01 and P<0.01).④The apoptotic peak was detected with FCM and cells were stayed in G1/S stage and a sub-Gl stage cell apoptotic peak appear in front of GO/G1 stage(P<0.01).Typical apoptotic morphologic feature was discovered under light microscope.Conclusion ①The HO-8910 ovary carcinoma cell growth was inhibited by Phosphorathioate antisense Oligonucleotide and appeared the apoptosis.②It showed that the rate of inhibition has the discrepancy between the different dose and different time.In other words,the rate of inhibition enhance along with the augment of the dose and time.③PS-ASODN inhibits strongly the proliferation of HO-8910 ovary carcinoma cell by inhibiting telomerase activity which induce cell apoptosis.PS-ASODN might be a new target to ovary carcinoma therapy.
【Key words】
Telomerase phosphorathioate antisense oligodeoxy nucleotides; HO-8910 ovary carcinoma cell; Telomerase; Cellapoptosis
目前隨著人類生存環境污染的加劇,生活方式的改變,癌癥已成為威脅人類健康和生命的主要殺手。卵巢癌作為我國最常見的惡性腫瘤之一 [1]。卵巢癌的發生和進展涉及多種癌基因和/或抑癌基因的異常改變以及幾個遺傳基因的遺傳失穩[2]。然而除了這些分子生物學變化之外,卵巢癌可能還有另外一個非常重要的生物學特性,即以細胞的永生化來維持它的無限生長[3]。研究表明這種永生化的維持主要是依賴腫瘤細胞中的端粒酶被激活后,使得端粒的長度維持到一定程度,從而使細胞獲得無限增殖的能力[4]。
端粒(Telomere)是真核細胞染色體末端含有串聯重復的DNA序列,其功能為體細胞分裂時端粒的末端會隨每次的復制被逐漸縮短,當縮短到一定程度不能維持染色體的穩定時細胞最終死亡。端粒長度保持在延長和縮短的動態平衡中,在這個平衡體系中端粒酶是關鍵因素[5]。有關研究報告在多種腫瘤細胞中都可能有端粒酶的活性[6]。端粒酶由蛋白催化亞基和指導端粒合成的RNA模板組成,即為具有逆轉錄酶特性的核蛋白酶[7]。端粒酶幾乎在所有惡性腫瘤組織中都有強陽性表達,而在正常體細胞及良性腫瘤組織中表達為陰性[8],因此端粒酶已成為新的腫瘤標志物 [9] 。
端粒酶的三個主要組成成分:端粒酶RNA (human telomerase RNA,hTR )、端粒酶相關蛋白質(telomerase protein 1,TP1/TLP1)及端粒酶催化亞單位(human telomerase reverse transcriptase,hTRT/hTERT)己經被闡明,基因序列也已經被克隆[10]。設想應用反義寡核苷酸封閉該模板區,就可以抑制端粒酶活性,阻止端粒序列的合成,使細胞退出增殖周期,達到治療腫瘤的目的。所以端粒酶不但是特異性惡性腫瘤標志物,更是一個腫瘤治療的理想靶點。
本實驗設計了一段互補于hTR模板區(GenBank登陸號為:NR_001566)的反義核酸,用寡核苷酸脂質體(OligofectamineTM)介導,將互補于人端粒酶RNA模板區的反義寡核苷酸作用于體外培養的卵巢癌HO-8910細胞,采用酶聯免疫吸附法及流式細胞學等技術觀察反義hTR對卵巢癌HO-8910細胞端粒酶活性、細胞增殖、細胞凋亡及細胞周期的影響,旨在探討端粒酶PS-ASODN在卵巢癌治療中的價值。
1 材料與方法
1.1 材料
1.1.1 細胞來源 人HO-8910卵巢癌細胞,遼寧醫學院科學實驗中心。
1.1.2 主要試劑及藥品達爾伯克必需基礎培養基(Deubeccos minimum essential; medium,DMEM)培養基有GIBCO公司生產;小牛血清(fetal bovine serum,FBS)由Hyclone公司生產;胰蛋白酶由Hyclone公司生產;端粒酶全硫代反義寡核苷酸(phosphorathioate antisense oligonucleotide,PS-ASODN)及對照正義全硫代寡核苷酸(phosphorathioate sense oligonucleotide,PS-SODN)均由上海生物工程公司合成。端粒酶PS-ASODN序列為5′-CTCAGTTAGGGTTAGACA-3′;端粒酶PS-SODN序列為5′-TGTCTAACCCTAACTGAG-3′;端粒酶檢測試劑盒由大連泛邦化工技術有限公司生產。恒溫培養箱由美國Forma Scientific生產;流式細胞儀由美國BD公司生產。
1.2 方法
1.2.1 引物的合成 端粒酶全硫代反義寡核苷酸(PS-ASODN)及對照正義全硫代寡核苷酸(PS-SODN):均由上海生物工程公司合成。PS-ASODN序列為5'-CTCAGTTAGGGTTAGACA-3';PS-SODN序列為5'-TGTCTAACCCTAACTGAG-3'。
1.2.2 實驗分組 實驗分組:空白對照(Control)組; 濃度為3 μmol/L的脂質體對照組;濃度為3 μmol/L的PS-SODN組; PS-ASODN濃度分別為1、2及 3 μmol/L三個濃度組。
1.2.3 實驗方法
1.2.3.1 細胞培養 卵巢癌HO-8910細胞以5×104/ml密度接種在含10%新鮮小牛血清、100 U/ml慶大霉素的RPMI-1640培養液中,置370C、5%CO2孵箱中培養,隔天換液一次,取對數生長期細胞用于實驗。
(1)細胞傳代:倒除原培養液,無菌PBS洗滌兩次后加入0.25%的胰蛋白酶0.5 ml消化細胞,倒置顯微鏡下觀察細胞變圓時倒掉胰酶,加入RPMI-1640培養液,用吸管輕輕吹打,制成細胞懸液,以適當細胞密度分裝于幾個培養瓶中,補加培養液,放入培養箱中培養。
(2)復蘇凍存的細胞:從-80℃冰箱中取出凍存管,迅速放入37℃水浴箱中搖勻解凍后,在超凈工作臺中,用吸管吸出細胞懸液裝入離心管,補加10 ml RPMI-1640培養液,混勻后以800轉/分離心5 min,倒除上清,加入培養液1 ml吹勻,再移入培養瓶中,如入培養液適當稀釋后,置培養箱培養,2 h后更換培養液一次,再繼續培養。
1.2.3.2 細胞轉染 先將裝有10 OD干膜狀寡聚脫氧核苷酸的EP管離心(10 000 g,10 s),然后加入550 μl雙蒸水,充分振蕩5 s,配成存儲濃度為100 μmol/L寡聚脫氧核苷酸溶液,之后再對其進行稀釋,使其存儲濃度為20 μmol/L以后根據實驗需求再稀釋(PS-SODN為3 μmo1/L; PS-ASODN為1 、2及3 μmo1/L)。
脂質體介導的端粒酶反義寡聚脫氧核苷酸轉染按說明書操作。實驗分6組,每組設4個復孔。6組分別為終濃度為3、 2及1 μmol/L的PS-ASODN組、終濃度為3 μmol/L的脂質體對照組、終濃度為3 μmol/L的PS-SODN組以及空白對照組(僅加培養液)。用藥后將小牛血清的濃度調至5%,以減少寡聚脫氧核苷酸的降解。24 h后棄去轉染液,換新鮮培養液,并繼續培養,收集細胞用于以下實驗。
1.2.3.3 ELISA法檢測端粒酶活性 ①細胞提取物的制備:取處理因素作用后的細胞5×104,PBS洗滌后離心去上清,加裂解液10 μl,冰浴放置30 min,4℃、1 5000 g離心10 min,取上清液保存于-800C,以備端粒酶活性檢測。按試劑盒說明書操作,檢測每組在450nm處的A值,并計算相應的端粒酶活性;②分組:本實驗按ELISA試劑盒說明書共分為8即:
陰性對照組:陰性反應液+酶聯反應物+底物A+底物B+終止液;
陽性對照組:陽性反應液+酶聯反應物+底物A+底物B+終止液;
空白對照組:空白反應液+酶聯反應物+底物A+底物B+終止液。作用時間為72 h的不同濃度PS-ASODN檢測組(3、2、1 μmol/L)、脂質體對照組(3 μmol/L)和 PS-SODN(3 μmol/L)。各濃度提取物+酶聯反應物+底物A+底物B+終止液;③實驗操作:96孔培養板每組4個孔重復3次,(各組的實驗條件都相同只有藥物濃度不同)。待試劑混勻、平衡到室溫(20~25℃)再進行實驗操作。
①陽性對照組加入50 μl陽性對照液;陰性對照組加入50 μl陰性對照液;空白對照組加入50 μl空白對照液;各實驗組分別加入50 μl不同濃度藥物的提取物;②8組中每孔都加入50 μl酶聯反應物,輕輕混勻,30 s后用自黏封孔蓋箔封住板孔,37℃溫育60 min;③洗板時甩盡板內液體,8組中每孔內分別加入洗滌液350 μl洗滌反應板,并去除水滴(在厚疊吸水紙上拍干),反復洗滌5次;④8組中每孔依次加入50 μl底物A和50 μl底物B;⑤8組中每孔加100 μl終止液輕輕混勻30 s,30 min內在450 nm處讀OD值(再換算成A值)。
結果判斷:檢測組的A值與空白組的A值之比(index)≥1.00為陽性,在0.91~0.99之間為可疑陽性:≤0.90為陰性。
1.2.3.4 吖啶橙染色法觀察細胞形態 取對數生長期細胞1×105/ml接種于24孔板中,細胞接種、轉染和培養方法同上。10 mg吖啶橙溶解于100 ml pH值為6.8的PBS中,4℃避光保存。取轉染3 μmol/L PS-ASODN 和3 μmol/L PS-SODN 72 h后的細胞,按照說明書操作,熒光顯微鏡觀察細胞形態。
1.2.3.5 MTT法測定PS-ASODN對卵巢癌HO-8910細胞增殖的影響 細胞以5×104/ml密度接種在96孔培養板上,每孔100 μl,待細胞生長活躍時加處理因素,分組方法與細胞轉染的方法一樣,分別培養24、48、72 h后檢測。檢測方法:每孔加入5 mg/ml的MTT 20 μl,繼續培養4 h,翻板倒掉液體,每孔加入二甲基亞砜(DMSO)150 μl,輕輕振蕩10 min,用自動酶標儀檢測 540nm 處的吸光度(A)值。
抑制率=(空白對照組平均A值-實驗組平均A值)/空白對照組平均A值 ×100%
1.2.3.6 流式細胞儀檢測細胞凋亡和細胞周期 ①PI染色:取對數生長期細胞1×105/ml細胞數接種于6孔培養板中,細胞接種、培養,轉染方法同端粒酶檢測實驗。將轉染72 h的細胞離心收集,PBS洗一次,離心棄PBS,用剩余的PBS充分混勻細胞,加入預冷的75%乙醇固定,混勻,用封口膜封口,4℃冰箱過夜。上機檢測前棄乙醇,PBS清洗細胞,調整細胞濃度至1×106/ml,加入PI(50 μg/ml) 800 μl混勻,4℃避光染色1 h,300目尼龍網濾過,上機檢測,進行DNA含量、細胞周期的分析,低于G1期DNA含量主峰(亞G1期峰)的細胞為凋亡細胞。每項實驗重復3次;②AnnexinV-FITC/PI染色:將各組細胞用胰酶消化,再用PBS洗滌,調整待測細胞的濃度為1×106個/ml。取1 ml細胞,1000 g,4℃離心10 min,棄上清;用PBS洗3次后將細胞重懸于200 μl Binding Buffer;加入10 μl AnnexinV-FITC和5 μl PI,輕輕混勻,避光室溫反應15 min;加入300 μl Binding Buffer,在1 h內上機檢測
1.2.4 統計學方法 實驗數據用(x±s)表示,用SPSS 11.5軟件包處理,組間比較采用單因素方差分析。
2 結果
2.1 PS-ASODN對卵巢癌HO-8910細胞端粒酶活性的影響 檢測72 h各組端粒酶活性陰陽性結果表明:3.0 μmol/L的PS-SODN和3.0 μmol/L脂質體對照組的端粒酶活性表現均為陽性;而濃度分別為1、2、3 μmol/L的PS-ASODN組端粒酶活性表現均為陰性,表明PS-SODN和脂質體對卵巢癌HO-8910細胞端粒酶活性無影響,而PS-ASODN 對卵巢癌HO-8910細胞端粒酶活性有抑制作用(見表1)。
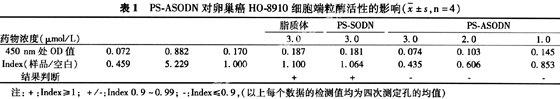
2.2 形態學觀察結果 吖啶橙染色72 h后熒光顯微鏡下觀察發現,空白對照組細胞呈圓形或橢圓形,胞漿豐富,胞核及分裂正常(見圖 1) ;PS-ASODN組卵巢癌HO-8910細胞體積縮小,核皺縮并有細胞膜出芽,有明顯凋亡現象,表明PS-ASODN具有促進卵巢癌HO-8910細胞凋亡的作用(見圖 2)。
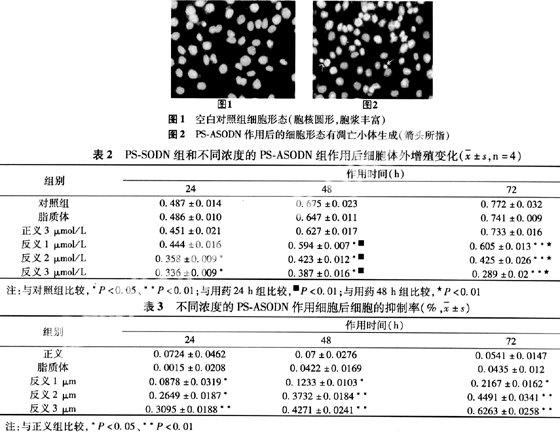
2.3 PS-ASODN對卵巢癌HO-8910細胞增殖的影響 與空白對照組比較,正義寡聚脫氧核苷酸組和脂質體組細胞增殖不顯著(P>0.05);濃度分別為1、2、3 μmol/L反義寡聚脫氧核苷酸組均能顯著抑制細胞增殖(P<0.05) 并呈一定的時間和劑量依賴關系(見表2)。
抑制率的計算結果表明,PS-ASODN各濃度組差異顯著(P<0.05),并呈一定的時間和劑量依賴關系,即抑制率隨著PS-ASODN的作用時間和劑量的增加而增大(見表3)。
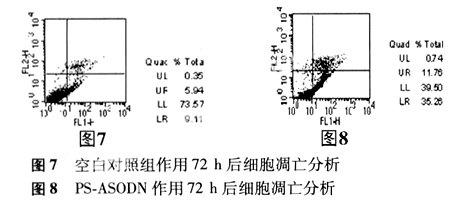
2.4 流式細胞儀檢測結果 與空白對照組比較,濃度分別為1.0、2.0、3.0 μmol/L的PS-ASODN轉染細胞72 h后,G0/G1期細胞明顯增多,差異顯著(P<0.01)(見表4),在G0/G1期前出現亞二倍體凋亡峰(見圖3、4、5、6),表明細胞被阻止在G1/S期。

在雙變量流式細胞儀的散點圖上,轉染72 h后空白對照組活細胞數量占73.57%,凋亡細胞數量占9.11%;濃度為3.0 μmol/L PS-ASODN組活細胞數量占39.50%,凋亡細胞數量占35.26%。表明PS-ASODN 能夠明顯促進細胞調亡(見圖9、圖10 )。
3 討論
端粒是真核線形染色體末端結構,由端粒DNA和端粒結合蛋白組成。它由一端高度保守的重復序列(TTAGGG ) n組成,方向5'→3',長度約5~15 kb[11],端粒長度的維持需要端粒酶的激活。自身攜帶模板是端粒酶區別于一般的純蛋白逆轉錄酶的主要特征之一。
端粒酶調節細胞分裂及細胞的壽命,參與細胞增殖、分化、衰老等活動,尤其與細胞的癌變密切相關,在正常人胚胎組織中,未分化的神經上皮細胞中有最高的hTR表達,大多數分化高的組織其表達缺乏或降低。成人組織中,睪丸細胞hTR高表達,淋巴小結中度表達,上皮細胞微弱表達,而且只限于特定的有絲分裂期細胞,神經系統和間質來源細胞無表達[12]。 1995年Feng[13]首次克隆了人端粒酶RNA (hTR)組分,發現反義或突變的RNA結構都可能影響端粒酶的活性,從而抑制腫瘤細胞增殖。
國外學者[14]對乳腺癌、卵巢癌、肺癌、膀胱癌、食管癌等進行廣泛深入研究。研究證實反義寡聚脫氧核苷酸對癌細胞的端粒酶活性和細胞增殖均起到了抑制作用,并促進細胞周期改變和誘導細胞發生凋亡。其作用機制是:特定反義核酸分子與進入細胞質的mRNA上的特異位點雜交,形成DNA-RNA復合物,抑制mRNA與核糖體的結合,同時激活RNase H,RNase H能使mRNA降解,從而使特定的基因不能表達,相應的蛋白質合成受到抑制[15] 。但尚未發現反義寡聚脫氧核苷酸能對卵巢癌HO-8910細胞起同樣作用的報道。
本實驗采用反義hTR轉染卵巢癌HO-8910細胞,為減少細胞對反義核酸的降解作用并增加其入胞效率,采用全硫代磷酸修飾其分子。 其中PS-ASODN是根據靶基因序列,以堿基互補配對方式設計而成,因此進入細胞后只能作用于靶基因,而不與非相關基因作用,故針對性強,特異性高且片段短,可以完全被細胞吸收進入細胞核,經硫代修飾后具有很強的抗核酶能力,再經寡聚脫氧核苷酸脂質體轉染入卵巢癌HO-8910細胞。實驗結果顯示反義寡聚脫氧核苷酸轉染細胞后,卵巢癌HO-8910細胞的增殖明顯受到抑制、端粒酶活性明顯降低,與空白對照組、脂質體組、正義寡聚脫氧核苷酸組比較,差異均有統計學意義(P<0.01),表明反義寡聚脫氧核苷酸是良好的端粒酶抑制劑。
另有研究結果表明,端粒酶活性與細胞增殖有關,因此,端粒酶抑制劑抑制了細胞端粒酶活性,自然也使其增殖受到抑制[16]。本實驗結果也證實,端粒酶反義寡聚脫氧核苷酸抑制端粒酶活性,同時也抑制了細胞增殖。
Zhu等[17]研究發現,端粒酶活性是細胞周期依賴性調控。端粒酶在DNA復制期激活,以保持端粒延長,因此,在細胞周期中,端粒酶活性經G1/S期逐漸升高,S期最高,G2/M最低,G0/G1期幾乎處于靜止期。
本研究FCM檢測提示:72 h后空白對照組細胞增殖旺盛,S期細胞占細胞周期的23.26%,但凋亡率為9.03%。PS-SODN處理組凋亡率僅為10.63%,兩者差異無顯著性(P>0.05)。而相同條件下PS-ASODN組FC-M檢測發現能阻止HO-8910細胞進入DNA合成期,使S期細胞明顯減少,降為5.47%,凋亡率為35.26%,表明細胞分裂增殖活動減弱,細胞被阻止在G1/S期。細胞凋亡時,細胞的DNA裂解,在流式細胞儀上呈現亞二倍體凋亡峰。表明反義寡聚脫氧核苷酸能夠誘導卵巢癌HO-8910細胞凋亡并促使細胞周期發生改變。根據FCM檢測結果推測PS-ASODN可能通過與端粒酶RNA模板的特異性結合而阻斷其表達,從而有效地抑制S期端粒酶活性,端粒合成受到抑制,使染色體末端不穩定,腫瘤細胞阻滯于M2期,細胞周期檢查點監控功能增強,激活了凋亡相關基因啟動細胞程序性死亡,導致腫瘤細胞凋亡。
一些學者認為[18-20],端粒酶抑制劑對細胞的增殖抑制可能與誘導凋亡有關。而端粒酶抑制劑誘導細胞凋亡可能通過兩條途徑:①端粒酶活性被抑制導致端粒延伸障礙,端粒長度不能維持,當細胞分裂導致端粒縮短到一定程度,通過p53等凋亡調節蛋白[21],誘導細胞凋亡;②端粒酶活性被抑制導致一些已經處于極短的端粒不能維持功能,細胞染色體發生斷裂、融合、DNA破壞,觸發細胞凋亡[22]。本實驗用吖啶橙染色轉染反義核酸的卵巢癌HO-8910細胞,在熒光顯微鏡下可見,卵巢癌HO-8910細胞出現細胞膜出芽,細胞核皺縮、碎裂、凋亡小體等明顯的凋亡形態學改變。
本實驗結果表明:(1)互補于hTR模板區的反義寡聚脫氧核苷酸明顯抑制卵巢癌HO-8910細胞的增殖及端粒酶活性,作用方式呈時間和劑量依賴性,即隨著反義寡聚脫氧核苷酸濃度加大和培養時間延長,抑制作用逐漸增強。這主要是由于反義寡聚脫氧核苷酸封閉了hTR模板區,阻止了端粒酶合成端粒,從而有效地抑制了卵巢癌HO-8910細胞的增殖;(2)反義寡聚脫氧核苷酸轉染卵巢癌HO-8910細胞后,端粒酶活性顯著下降。筆者認為,很可能反義寡聚脫氧核苷酸本身不一定具有調節凋亡的作用,其對凋亡的調節可能是通過改變端粒長度所致,因為端粒長度的維持、染色體的穩定是凋亡的一個抑制因素,而端粒的縮短可觸發細胞凋亡。以上結果說明反義寡聚脫氧核苷酸能夠通過抑制hTR基因的表達,抑制卵巢癌HO-8910細胞端粒酶活性,從而達到抑制卵巢癌HO-8910細胞的增殖。這也為卵巢癌基因治療提供了新的靶點,結合其他治療方法有望提高卵巢癌基因治療的成功率。
盡管端粒酶在幾乎所有惡性腫瘤都有強陽性表達,而且反義hTR寡核苷酸制劑由于特異性封閉hTR模板區而抑制了端粒酶活性,在腫瘤的治療方面顯示出巨大的潛力。但對端粒酶陽性的生殖細胞和干細胞所帶來的后果不得不引起我們的關注,尤其是生殖細胞,如果針對腫瘤細胞的端粒酶抑制劑同樣可以進入人生殖細胞,使其端粒酶失活,后果將是一個種屬的滅絕;但也有人持不同的觀點,認為正常細胞的端粒較多數腫瘤細胞的長,并且原始干細胞較少進行有絲分裂,其端粒縮短的速度遠低于腫瘤細胞,因此端粒酶抑制劑的療程可在正常細胞的端粒酶耗盡前停止,當抑制端粒酶治療結束后,正常細胞的端粒酶活性可得到修復。此外部分腫瘤細胞具有較長的端粒,而隨細胞有絲分裂的進行,端粒的縮短是一個緩慢的過程,在這種情況下端粒酶抑制劑是否有效,尚待進一步研究。
參 考 文 獻
[1] 李連弟,魯鳳珠,張思維,等. 1990-1992年中國惡性腫瘤死亡流行分布情況分析.中華腫瘤雜志,1996,18:403-407.
[2] Palmer JE,Sant Cassia LJ,Irwin CJ,et al.The prognostic and predictive value of syntactic structure analysis in serous carcinoma of the ovary.Int J Gynecol Pathol,2008,27(2):191-198.
[3] Zhang PN,Sun H.Expression of phosphatidylinositol-3 kinase in epithelial ovarian carcinoma.Zhonghua Fu Chan Ke Za Zhi,2007,42(3):196-200.
[4] Sun PM,Wei LH,Luo MY,et al.The telomerase activity and expression of hTERT gene can serve as indicators in the anti-cancer treatment of human ovarian cancer.Eur J Obstet Gynecol Reprod Biol,2007,130(2):249-257.
[5] Jiao XL,Chen ZX,Cen JN,et al.Expression of human telomerase reverse transcriptase and survivin gene in patients with myelodysplastic syndrome.Zhongguo Shi Yan Xue Ye Xue Za Zhi,2008,16(2):294-298.
[6] 衛建平,吳麗然,王晨,等.端粒酶hTERT蛋白與基因在人腫瘤細胞中表達的相分析.中國藥物與臨床,2004,4(3):187-190.
[7] Cairney CJ,Hoare SF,Daidone MG.et al.High level of telomerase RNA gene expression is associated with chromatin modification,the ALT phenotype and poor prognosis in liposarcoma.Br J Cancer,2008,98(8):1467-1474.
[8] Dasi F,Martinez-Rodes P,March JA,et al.Real-time quantification of human telomerase reverse transcriptase mRNA in the plasma of patients with prostate cancer.Ann N Y Acad Sci,2006,1075:204-210.
[9] Sun L,Wang F,Sun H,et al.Effects of c-myc antisense oligodeoxynucleotide on the telomerase activity and the induction of apoptosis in HL-60 cells.Zhongguo Shi Yan Xue Ye Xue Za Zhi,2006,13(4):605-609.
[10] Nakayama J,Tahara H,Tahara E,et al.Telomerase activity by hTRT in human normal fibroblasts and hepatocellular carcinomas.Nat Genet,1998,18:65-68.
[11] Shammas MA,Koley H,Batchu RB,et al.Telomerase inhibition by siRNA causes senescence and apoptosis in Barrett's adenocarcinoma cells:mechanism and therapeutic potential.Mol Cancer,2007,15(4):24.
[12] Zhang S,Dong M,Teng X,et al.Quantitative assay of telomerase activity in head andneck squamous cell carcinoma and other tissues.Arch Otolaryngol Head Neck Surg,2005,127(5):581-585.
[13] Feng 1,Walter DF,Wang SS,et al.The RNA component of human telomerase.Science,1995,269:1236-1240.
[14] Zorn KK,Jazaeri AA,Awtrey CS,et al.Choice of normal ovarian control influences determination of differentially expressed genes in ovarian cancer expression profiling studies.Clin Cancer Res,2003,9(13):4811-4818.
[15] Zamaratski E,Pradeepkumar PI,Chattopadhyaya J.A critical survey of the structure-function of the antisense oligo/RNA heteroduplex as substrate for RNAse H.J Biochenm Biophys Methods,2006,48(3):189-208.
[16] Chen Z.Koeneman KS.Corey DR.Consequences of telomerase inhibition and combination treatments for the proliferation of cancer cells Cancer Res,2005,63:5917-5925.
[17] Tonooka K,Kabashima T,Shibata T.et al.Facile Assay of Telomerase Activity Utilizing a DNA-detectable Chemiluminogenic Reagent.Anal Sci,2008,24(4):471-475.
[18] Chen M,Yang B,Zhang X.Effects of telomerase activity on carcinogenesis and survival in laryngeal carcinoma.J Int Med Res,2008,36(2):336-342.
[19] Wong SC,Yu H,Moochhala SM,et al.Antisense telomerase induced cell growth inhibition,cell cycle arrest and telomerase activity down-regulation in ovary and colon cancer cells.Anticancer Res,2003,23:465-469.
[20] Feng RH,Zhu ZG,Li JF,et al.Inhibition of human telomerase in MKN-45 cell line by antisense hTR expression vector induces cell apoptosis and growth arrest.World J Gastroenterol,2007,8(3):436-400.
[21] Yamaguchi K,Chijiiwa K,Torata N.et al.Telomerase activity,P53 mutation and Ki-ras codon 12 point mutation of the peripheral blood in patients with hepato pancreato biliary diseases.HPB (Oxford),2002,4(2):75-82.
[22] Glover L,Alsford S,Beattie C.et al.Deletion of a trypanosome telomere leads to loss of silencing and progressive loss of terminal DNA in the absence of cell cycle arrest.Nucleic Acids Res,2007,35(3):872-880.
